不同品種小麥幼苗耐鎘差異
2018-05-10 08:11:40王華忠岳潔瑜
江蘇農(nóng)業(yè)科學(xué) 2018年7期
張 欣, 王華忠, 王 利, 岳潔瑜
(天津市動植物抗性重點(diǎn)實(shí)驗(yàn)室/天津師范大學(xué),天津 300387)
鎘是一種毒性較強(qiáng)的重金屬元素,與其他重金屬相比,鎘在土壤中移動性較強(qiáng),更易被植物吸收積累[1]。而且鎘不是植物生長中的必需元素,它不參與生物的代謝活動,當(dāng)植物過量吸收時,會產(chǎn)生脅迫作用,對植物生長、發(fā)育、生理生化等都有影響[2-4],使植物體內(nèi)代謝紊亂,生長發(fā)育遲緩,產(chǎn)量下降,甚至導(dǎo)致植物死亡[5]。
已有研究表明,鎘是目前造成農(nóng)田受污染的毒性最強(qiáng)、最普遍的重金屬污染之一。我國目前正遭受鎘污染的農(nóng)田區(qū)域包括11個省25個地區(qū),如江西新余、浙江溫州、遼寧沈陽張土灌區(qū)、貴州赫章鉛鋅礦鎘污染區(qū)面積等,污染面積已超過28萬hm2。天津和廣東省廣州市郊等受鎘脅迫的區(qū)域,產(chǎn)出的含鎘超標(biāo)的農(nóng)產(chǎn)品近150萬t/年,嚴(yán)重威脅環(huán)境、動物和人類健康[6]。對重金屬典型污染區(qū)冬小麥及土壤重金屬進(jìn)行分析發(fā)現(xiàn),冬小麥田土壤中鎘污染最嚴(yán)重,超標(biāo)4.5倍,且冬小麥果實(shí)中鎘含量可達(dá)2.17 mg/kg,遠(yuǎn)超過GB2762—2012《食品污染物限量標(biāo)準(zhǔn)》中規(guī)定的限值[7]。降低鎘在作物中的累積,是保障農(nóng)產(chǎn)品質(zhì)量安全的重要途徑。因此,闡釋鎘離子在小麥幼苗體內(nèi)的轉(zhuǎn)運(yùn)和累積的規(guī)律、解析小麥響應(yīng)鎘脅迫的生理基礎(chǔ),可為找到減少重金屬損害小麥的有效措施和提高重金屬耐性水平的辦法提供理論基礎(chǔ),對保障農(nóng)作物安全和人類健康,增強(qiáng)農(nóng)業(yè)可持續(xù)生產(chǎn)尤為重要。
在已取得的研究成果中,關(guān)于鎘脅迫對植物形態(tài)及生理生化影響的報(bào)道居多,而且這些研究主要集中在擬南芥與水稻等傳統(tǒng)模式植物、東南景天與龍葵等超富集植物、楊樹等木本植物上[8-9]。目前,國內(nèi)外有關(guān)鎘脅迫對小麥的研究主要集中在鎘脅迫對小麥幼苗生長及抗氧化系統(tǒng)[10]、小麥植株元素分布[11]、小麥細(xì)胞染色體等細(xì)胞學(xué)水平上的影響[12];不同基因型作物及不同品種作物在響應(yīng)重金屬脅迫中表現(xiàn)出耐性的差異,且這種耐性差異受到植物不同生育期的影響,植物的發(fā)芽期和苗期對鎘脅迫比較敏感,因?yàn)榇藭r植物的防御機(jī)制尚未完全建立,因此出苗期和苗期可以作為對鎘脅迫進(jìn)行鑒定的時期[13]。本研究以津農(nóng)6號、中麥629、揚(yáng)麥158、92R137、濟(jì)麥22、山農(nóng)20、良星66、輪選519、衡觀35、河農(nóng)825、河農(nóng)6425、輪選987和京冬8號13個小麥常規(guī)品種為試材,采用苗期水培法,以空白對照組與添加100 μmol/L Cd2+的試驗(yàn)組進(jìn)行對比試驗(yàn),主要分析鎘處理下13個小麥品種幼苗期間的生理指標(biāo)變化,地上部分與地下部分的鎘含量、鎘富集系數(shù)和轉(zhuǎn)移能力,對比13個小麥品種對鎘脅迫的耐受性程度,為進(jìn)一步研究和篩選小麥耐鎘品種提供理論依據(jù)與數(shù)據(jù)參考。
1 材料與方法
1.1 不同品種小麥幼苗的培養(yǎng)及處理
本試驗(yàn)于2016年11月7日在天津師范大學(xué)進(jìn)行。選取大小均勻一致、飽滿、色澤勻稱、表皮光滑、健康無損傷的13個常規(guī)小麥品種種子,用自來水沖洗3~4次,再將種子分別單層放于鋪有3層濕潤紗布的培養(yǎng)盆中,自來水培養(yǎng)(25 ℃),間隔12 h換1次水,待小麥幼苗長至2葉1心時,從中篩選生長一致的麥苗,分別移栽至帶孔泡沫板上,并用脫脂棉固定,13個供試品種的80株麥苗分為對照組和處理組,分別移至含0、100 μmol/L Cd2+的1/10 Hoagland營養(yǎng)液中培養(yǎng),每3 d更換1次營養(yǎng)液。
1.2 試驗(yàn)方法
1.2.1 形態(tài)觀察及測定 在鎘處理小麥幼苗期間,每天觀察小麥幼苗生長狀況,每隔2 d測量1次生理指標(biāo),每次測量每個品種的對照組和試驗(yàn)組15株小麥幼苗,包括株高、葉長、葉寬、最長根長。數(shù)碼相機(jī)拍照。
1.2.2 小麥幼苗中鎘含量的測定 鎘脅迫處理小麥幼苗 14 d 后,分別收獲對照組和試驗(yàn)組小麥幼苗的地上和地下部分,先后用自來水、蒸餾水和去離子水沖洗干凈,晾干表面水分,分別稱鮮質(zhì)量,再放于烘箱中45 ℃烘2 d,之后85 ℃烘干至恒質(zhì)量,分別稱量地上部和地下部干質(zhì)量,再分別研磨呈粉末狀態(tài),用于消化分析。消化前分別稱取研磨粉末0.2 g置于潔凈的小燒杯中,緩慢加入濃硝酸5 mL浸泡過夜;再依次加入高氯酸2 mL,80 ℃加熱3 h后,再120 ℃加熱除去剩余硝酸。冷卻后再依次加入濃硝酸3 mL、高氯酸2 mL,加熱至燒杯中溶液呈橙黃色時再加入高氯酸2 mL,200 ℃加熱至燒杯中有白色晶體析出。晶體冷卻后,用稀硝酸溶液(10%)溶解和定容后,用電感耦合等離子發(fā)射光譜ICP-AES(LEEMAN LABS INC.,New Hampshire,USA)測定鎘在小麥幼苗地上部分和地下部分器官中的積累量。每個樣品重復(fù)測3次。
1.3 數(shù)據(jù)處理與分析
本試驗(yàn)數(shù)據(jù)處理過程中所用到的公式如下:
(1)變異系數(shù)(CV,%)。
變異系數(shù)=標(biāo)準(zhǔn)差/平均值×100%。
(2)轉(zhuǎn)移系數(shù)。
轉(zhuǎn)移系數(shù)=地上部分(莖葉)鎘含量/根部鎘含量[14]。
(3)根系耐性指數(shù)。
根系耐性指數(shù)=處理組根的平均長度/對照組根的平均長度[15]。
(4)含水量(%)。
含水量=(鮮質(zhì)量-干質(zhì)量)/鮮質(zhì)量×100%。
(5)耐鎘指數(shù)。
耐鎘指數(shù)=鎘處理下的值/對照值[16]。
試驗(yàn)中的數(shù)據(jù)采用Microsoft Office Excel 2007和IBM SPSS Statistics 22.0軟件進(jìn)行方差分析、相關(guān)性分析和聚類分析。
2 結(jié)果與分析
2.1 鎘處理對小麥幼苗生長的影響
與對照組相比,鎘處理后的小麥葉長和根長均變短(圖1),且隨著處理時間變長,抑制效果越明顯,葉片黃化并出現(xiàn)黃褐色斑點(diǎn)(圖1-b),觀察葉片黃褐色斑點(diǎn)出現(xiàn)時間,其中92R137、中麥629、輪選519在鎘處理2 d時出現(xiàn)黃褐色斑點(diǎn);津農(nóng)6號、山農(nóng)20、良星66、京冬8號、衡觀35、河農(nóng)825、河農(nóng)6425、輪選987在處理4 d時先后出現(xiàn)黃褐色斑點(diǎn);濟(jì)麥22、楊麥158在處理6 d后開始出現(xiàn)黃褐色斑點(diǎn)。葉寬幾乎沒有差異。

試驗(yàn)期間,每隔2 d測量1次小麥的生長指標(biāo)。對比各小麥生理指標(biāo)來看,津農(nóng)6號小麥在鎘處理之后一直處于生長狀態(tài);輪選519、河農(nóng)825和輪選987小麥在鎘處理后幾乎沒有生長;其他品種小麥在鎘處理4~6 d后停止生長。變異系數(shù)分析表明,小麥幼苗葉長的變異系數(shù)變化范圍最大(表1、表2、表3)。變異程度是衡量一個性狀能否作為篩選指標(biāo)的標(biāo)準(zhǔn),變異系數(shù)越大,說明該指標(biāo)越靈敏[17]。因此,小麥幼苗葉長可作為分析鎘脅迫效應(yīng)的生理指標(biāo)。
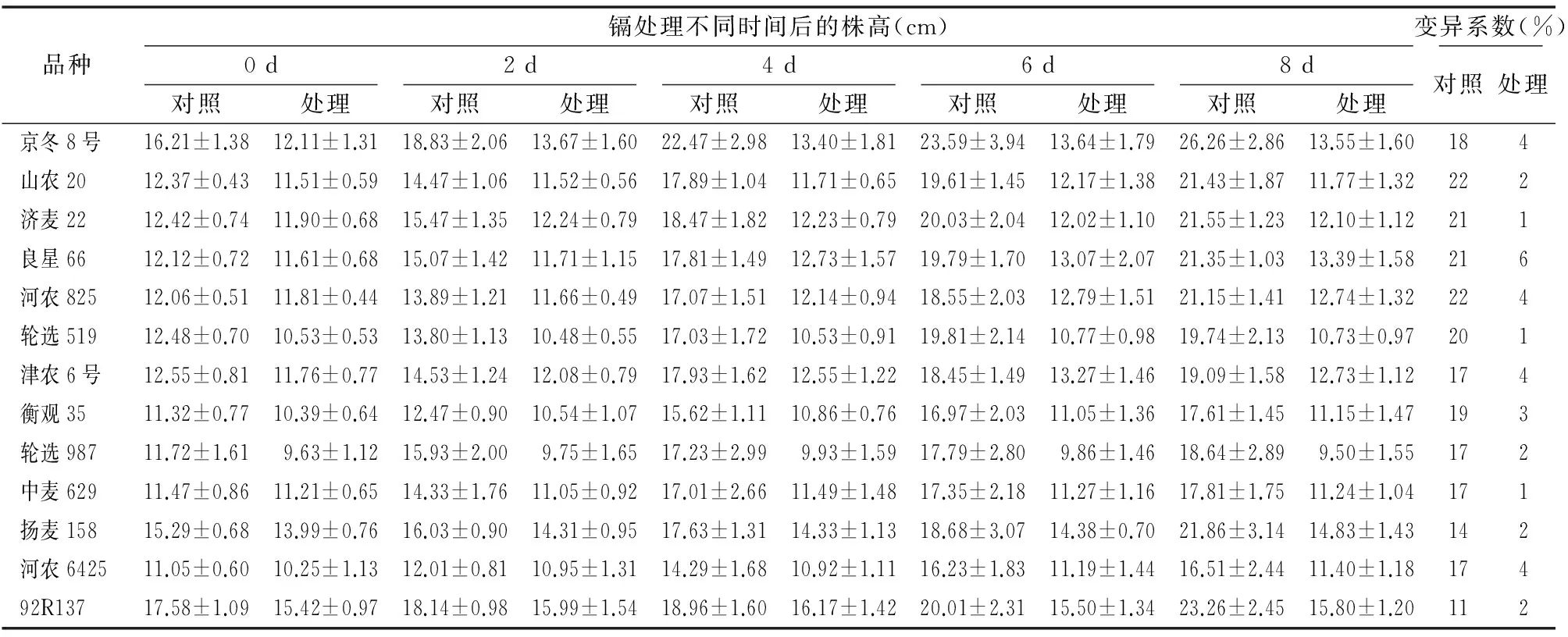
表1鎘處理不同時間對小麥幼苗株高的影響
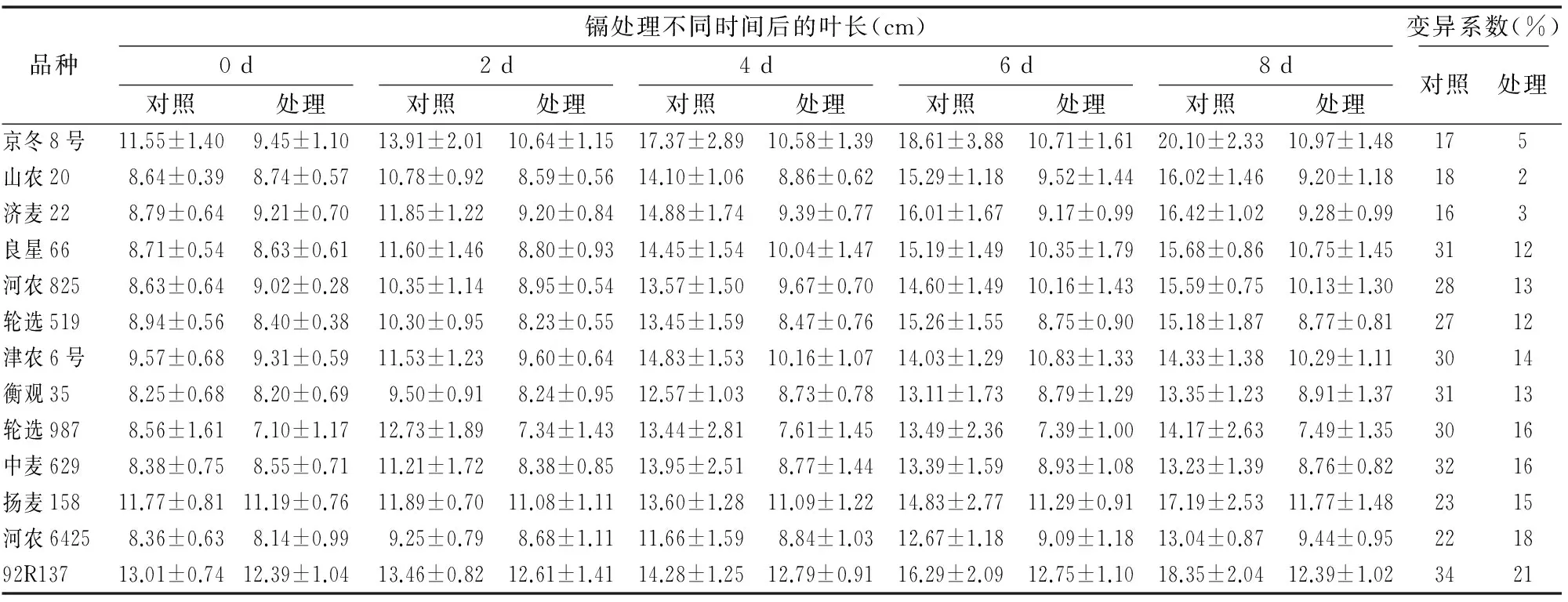
表2鎘處理不同時間對小麥幼苗葉長生長情況

表3鎘處理不同時間對小麥幼苗最長根長生長情況
2.2 含水量
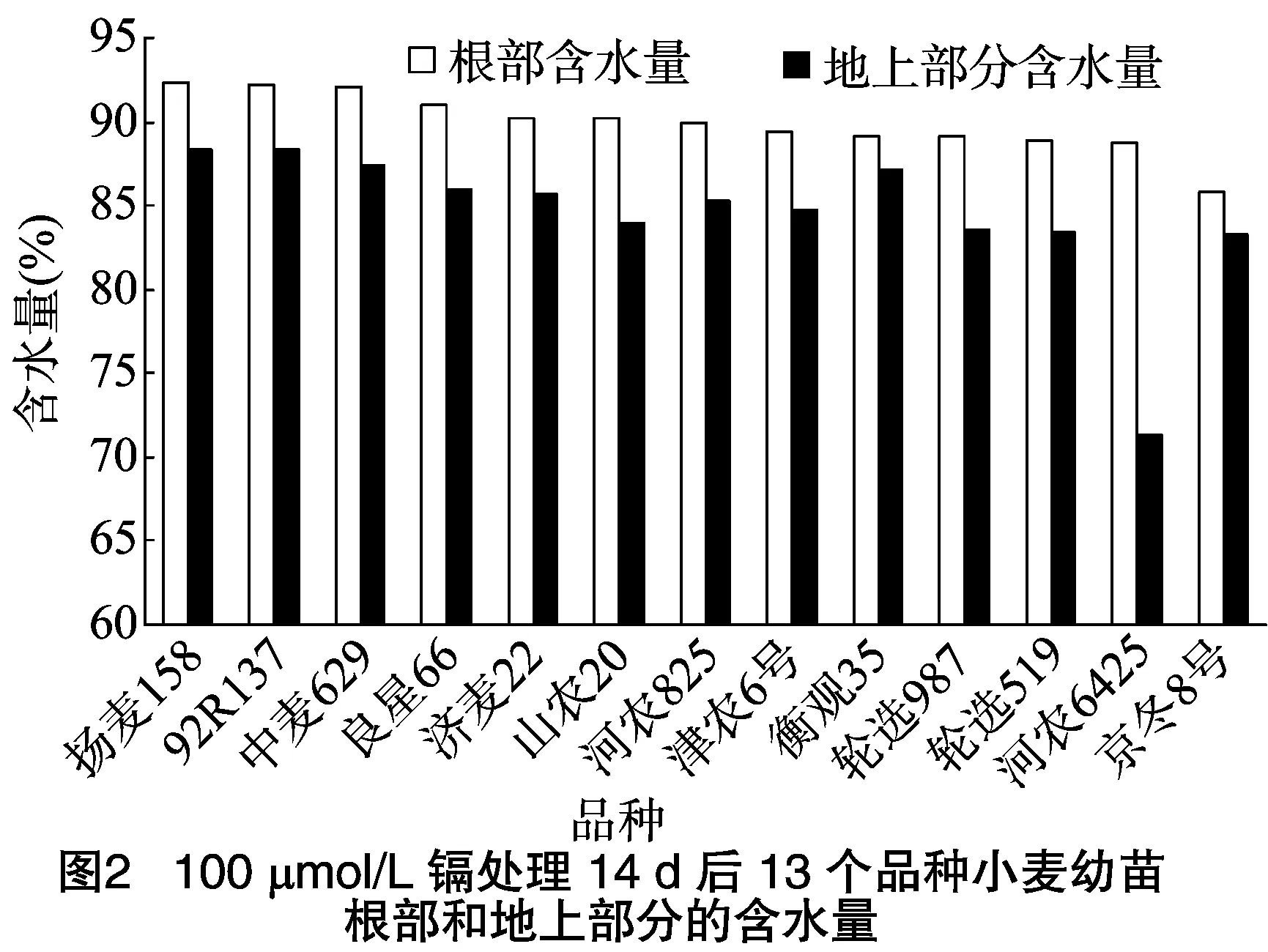
水分是植物的重要組成部分,含水量能在一定程度上顯示其生長狀況,自由水含量越大,代謝越旺盛[18]。對照組小麥幼苗根部平均含水量為93%,地上部分平均含水量89%,均高于其他處理組小麥含水量。由圖2可知,揚(yáng)麥158和92R137這2個品種小麥的根部含水量和地上部分含水量均高于其他品種,生長活性高于其他品種小麥,河農(nóng)6425的地上部分含水量僅為71%,葉片枯黃,受鎘脅迫嚴(yán)重。
2.3 鎘含量、鎘轉(zhuǎn)移系數(shù)與根系耐鎘指數(shù)
不同品種小麥鎘轉(zhuǎn)移能力差異較大,13個品種的鎘轉(zhuǎn)移系數(shù)總的變化范圍為0.31~0.68,13個品種小麥的鎘轉(zhuǎn)移能力由高到低依次為濟(jì)麥22>輪選987>92R137>良星66>輪選519>揚(yáng)麥158>河農(nóng)825>中麥629>衡觀35>京冬8號>山農(nóng)20>津農(nóng)6號,鎘轉(zhuǎn)移能力強(qiáng)的品種能將較多的鎘轉(zhuǎn)運(yùn)至莖葉等地上部分,說明其體內(nèi)可能存在較好的鎘運(yùn)輸和解毒方式。濟(jì)麥22根中鎘含量最少,轉(zhuǎn)移系數(shù)最高;津農(nóng)6號、山農(nóng)20和河農(nóng)8425的根中鎘含量較多但轉(zhuǎn)移系數(shù)小;京冬8號根中鎘含量最高,但轉(zhuǎn)移系數(shù)較低。92R137、揚(yáng)麥158、濟(jì)麥22、輪選987和津農(nóng)6號的耐性較高,但是結(jié)合根部吸收鎘的總量來看,輪選987、揚(yáng)麥158和濟(jì)麥22吸收量較少,而92R137和津農(nóng)6號在鎘吸收量比較多的情況下仍保持較高耐性,所以92R137和津農(nóng)6號的耐鎘性較好,92R137轉(zhuǎn)移系數(shù)大于津農(nóng)6號,而根系耐性指數(shù)與轉(zhuǎn)移系數(shù)沒有表現(xiàn)出相關(guān)性(圖3、圖4)。
2.4 不同品種間小麥幼苗耐鎘指數(shù)的相關(guān)性分析
由表4可知,6個耐鎘指數(shù)的變異系數(shù)為1.98%~15.98%,相對根長的變異系數(shù)最大,表明相對根長對鎘毒害反應(yīng)敏感。由表5可知, 相對株高和相對葉長之間存在極顯著正相關(guān);相對根長和根含水量之間亦存在極顯著正相關(guān);相對地上部分含水量與相對株高和相對根長呈顯著正相關(guān)。

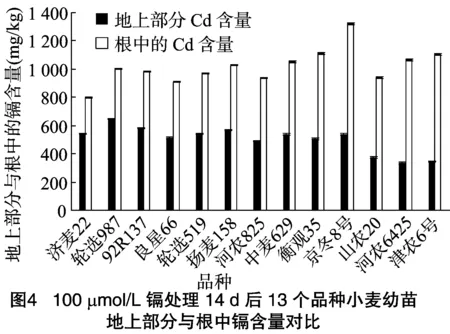
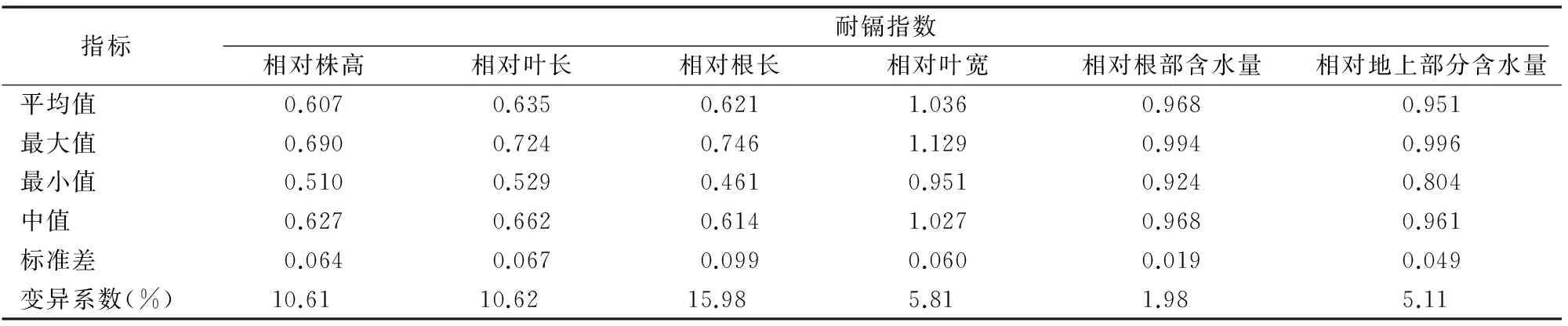
表4100 μmol/L 鎘處理14 d后13個供試品種小麥幼苗耐鎘指數(shù)的描述性統(tǒng)計(jì)
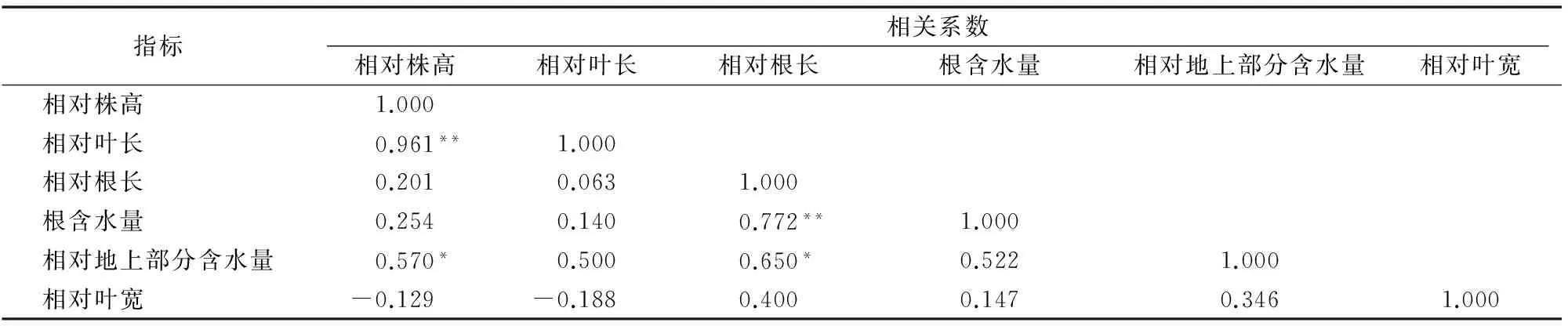
表5100 μmol/L鎘處理14 d后13個品種小麥幼苗耐鎘指數(shù)的相關(guān)性分析
注:*、**分別表示在0.05、0.01水平差異顯著。
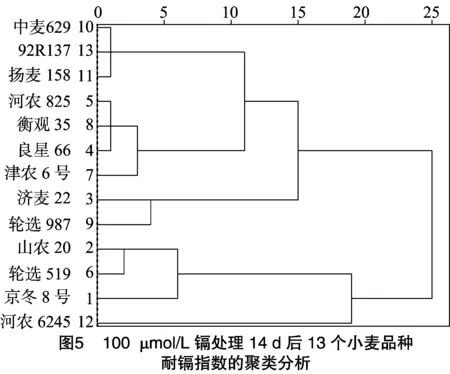
根據(jù)相對根長和相對葉長的耐鎘指數(shù)對13個不同小麥品種進(jìn)行聚類分析(圖5)可知,依據(jù)耐鎘指數(shù),13個供試小麥品種可分成耐鎘型、較耐鎘型、鎘較敏感型、鎘敏感型4種類型。其中耐鎘型品種包括中麥629、92R137、揚(yáng)麥158、河農(nóng)825、衡觀35、良星66和津農(nóng)6號;較耐鎘型品種包括濟(jì)麥22和輪選987;鎘較敏感型品種包括山農(nóng)20、輪選519、京冬8號;鎘敏感型品種包括河農(nóng)6425。
3 討論與結(jié)論
鎘污染是植物生長過程中遭受逆境脅迫中最普遍的重金屬污染之一。鎘離子在土壤中的高度移動性致使其容易進(jìn)入植物體內(nèi),累積到一定程度,擾亂植物新陳代謝過程,從而對植物產(chǎn)生毒害[19]。鎘對植物的毒害首先直觀地表現(xiàn)在生物量的降低、植物變矮、葉片黃化、根的生長受到抑制等。本研究表明,100 μmol/L鎘處理對13個小麥品種幼苗生長均表現(xiàn)出抑制效應(yīng),且鎘處理時間越長,抑制效應(yīng)越明顯。不同品種的小麥在鎘脅迫下亦表現(xiàn)出明顯的差異[20],通過對13種小麥的耐鎘性分析可為進(jìn)一步篩選耐鎘品種提供參考。
在評價(jià)植物響應(yīng)鎘脅迫的眾多生物學(xué)指標(biāo)中,轉(zhuǎn)移系數(shù)可反映鎘在植物不同器官中的轉(zhuǎn)移規(guī)律。轉(zhuǎn)移系數(shù)可以用植物體內(nèi)重金屬含量與培養(yǎng)液中重金屬含量的比值來計(jì)算。已有研究表明,轉(zhuǎn)移系數(shù)可以用來評價(jià)植物由下至上轉(zhuǎn)移重金屬的能力[21-23]。轉(zhuǎn)移系數(shù)高說明植物能在鎘脅迫環(huán)境下,將吸收的鎘較多地運(yùn)輸?shù)降厣喜课弧Q芯勘砻鳎参锬軐⒏糠e累的鎘運(yùn)輸?shù)降厣锨o、葉中,植物體在重金屬鎘脅迫下可能存在較好的運(yùn)輸和解毒機(jī)制,可利用自身的解毒機(jī)制減少鎘產(chǎn)生的危害[14]。本研究中13個小麥品種幼苗對鎘的轉(zhuǎn)移能力差異顯著,如濟(jì)麥22根中鎘含量最少,轉(zhuǎn)移系數(shù)最高;津農(nóng)6號、山農(nóng)20和河農(nóng)825根中鎘含量較多,但轉(zhuǎn)移系數(shù)小;京冬8號根中鎘含量最高,轉(zhuǎn)移系數(shù)較低。進(jìn)一步分析13個品種小麥幼苗的生長、含水量等6個耐性指數(shù),結(jié)果表明,92R137、揚(yáng)麥158、濟(jì)麥22、輪選987和津農(nóng)6號的鎘耐性較高,但是結(jié)合根部吸收鎘的總量來看,輪選987、揚(yáng)麥158和濟(jì)麥22吸收量較少,而92R137和津農(nóng)6號在吸收量比較多的情況下仍保持較高耐性,所以92R137和津農(nóng)6號的耐鎘性較好,92R137轉(zhuǎn)移系數(shù)大于津農(nóng)6號,根系耐性指數(shù)與轉(zhuǎn)移系數(shù)沒有表現(xiàn)出相關(guān)性。
在重金屬脅迫下,植物根系對重金屬產(chǎn)生吸收或排斥作用,有研究表明,重金屬在植物根中的累積可導(dǎo)致植物根尖分生區(qū)細(xì)胞有絲分裂過程異常,表現(xiàn)為植物的生理生化過程改變,阻礙其正常生長發(fā)育進(jìn)程[24-25]。敏感型小麥在重金屬脅迫下,易受重金屬影響,會導(dǎo)致植物生長緩慢,抑制根系生長;耐鎘型小麥則受重金屬影響較小。可用根系耐性指數(shù)來評價(jià)植物耐受重金屬脅迫能力的大小[26]。本研究中的根系耐性指數(shù)與轉(zhuǎn)移系數(shù)沒有表現(xiàn)出相關(guān)性,因此轉(zhuǎn)移系數(shù)不能作為篩選耐鎘性小麥品種的依據(jù)。
植物生長在受到鎘脅迫之后,生長指標(biāo)最先發(fā)生變化,如根、莖、葉等生物量都會受到影響[7]。研究表明,13個小麥品種幼苗的生長均受到鎘脅迫的抑制,表現(xiàn)為不同程度的損傷。其中相對葉長在生長指標(biāo)的測量中變異系數(shù)最大,表明相對葉長對鎘處理反應(yīng)比較敏感,可以作為篩選耐鎘性品種的指標(biāo)之一。在耐鎘指數(shù)的相關(guān)性分析中表明,相對根長對于鎘處理反應(yīng)敏感,也可作為篩選耐鎘品種的指標(biāo)。本研究結(jié)果與蔡保松等研究在鎘污染下小麥根的長度對鎘毒害反應(yīng)的結(jié)果[27]一致。李惠英等研究也表明,作物根系對重金屬元素反應(yīng)很敏感,也可以在一定程度上反映出對重金屬的忍耐能力[19]。因此,本研究中的聚類分析以各品種小麥的相對葉長和相對根長作為分析根據(jù)。分析結(jié)果表明,根系耐性指數(shù)和耐鎘指數(shù)分析結(jié)果基本一致,可以作為篩選依據(jù)。
綜上所述,在濃度為100 μmol/L的鎘處理下,13種小麥幼苗表現(xiàn)出不同程度的耐鎘性。根據(jù)小麥幼苗耐鎘指數(shù),可將13個小麥品種分為4種類型,包括耐鎘型、較耐鎘型、鎘較敏感型和鎘敏感型。13個小麥品種中中麥629、92R137耐性較強(qiáng),河農(nóng)6425耐性最弱。
參考文獻(xiàn):
[1]歐陽燕莎,劉愛玉,李瑞蓮. 鎘對作物的影響及作物對鎘毒害響應(yīng)研究進(jìn)展[J]. 作物研究,2016,30(1):105-110.
[2]張國平,深見元弘,關(guān)本根. 不同鎘水平下小麥對鎘及礦質(zhì)養(yǎng)分吸收和積累的品種間差異[J]. 應(yīng)用生態(tài)學(xué)報(bào),2002,13(4):454-458.
[3]顧翠花,王懿祥,白尚斌,等. 四種園林植物對土壤鎘污染的耐受性[J]. 生態(tài)學(xué)報(bào),2015,35(8):2536-2544.
[4]慈敦偉,姜 東,戴廷波,等. 鎘毒害對小麥幼苗光合及葉綠素?zé)晒馓匦缘挠绊慬J]. 麥類作物學(xué)報(bào),2005,25(5):88-91.
[5]張 玲,王煥校. 鎘脅迫下小麥根系分泌物的變化[J]. 生態(tài)學(xué)報(bào),2002,22(4):496-502.
[6]王維薇,林 清. 國內(nèi)外土壤鎘污染及其修復(fù)技術(shù)的現(xiàn)狀與展望[J]. 綠色科技,2017(4):90-93,102.
[7]孫洪欣,薛培英,趙全利,等. 鎘、鉛積累與轉(zhuǎn)運(yùn)在冬小麥品種間的差異[J]. 麥類作物學(xué)報(bào),2015,35(8):1161-1167.
[8]何彩甄,王建煌,裴 婕,等. 少花龍葵與蘆薈間種對鎘脅迫的效應(yīng)[J]. 安徽農(nóng)業(yè)科學(xué),2015,43(10):96-100.
[9]鄂志國,張玉屏,王 磊. 水稻鎘脅迫應(yīng)答分子機(jī)制研究進(jìn)展[J]. 中國水稻科學(xué),2013,27(5):539-544.
[10]Kumari N,Parmar P,Sharma V. Differential gene expression in two contrasting wheat cultivars under cadmium stress[J]. Biologia Plantarum,2015,59(4):701-707.
[11]Liu W T,Liang L C,Zhang X E,et al. Cultivar variations in cadmium and lead accumulation and distribution among 30 wheat (TriticumaestivumL.) cultivars[J]. Environmental Science and Pollution Research,2015,22(11):8432-8441.
[12]龐 楠,張飛雄. 低氧對小麥根端分生細(xì)胞核仁結(jié)構(gòu)和功能的影響[J]. 西北植物學(xué)報(bào),2013,33(3):507-511.
[13]孟桂元,唐 婷,周 靜,等. 不同玉米品種種子萌發(fā)期耐鎘性分析[J]. 分子植物育種,2016(11):3166-3171.
[14]顧翠花,王懿祥,白尚斌,等. 四種園林植物對土壤鎘污染的耐受性[J]. 生態(tài)學(xué)報(bào),2015,35(8):2536-2544.
[15]燕傲蕾,吳亭亭,王友保,等. 三種觀賞植物對重金屬鎘的耐性與積累特性[J]. 生態(tài)學(xué)報(bào),2010,30(9):2491-2498.
[16]慈敦偉,姜 東,戴廷波,等. 長江中下游麥區(qū)小麥地方品種幼苗耐鎘性的差異[J]. 麥類作物學(xué)報(bào),2011,31(6):1035-1039.
[17]徐順莉,房偉民,管志勇,等. 耐低鉀切花菊品種篩選及其苗期耐性生理研究[J]. 園藝學(xué)報(bào),2013,40(12):2463-2471.
[18]馮漢青,杜變變,王慶文,等. 鎘脅迫下活性炭對小麥幼根的保護(hù)作用[J]. 生態(tài)學(xué)報(bào),2016,36(10):2962-2968.
[19]李惠英,田魁祥,趙欣勝. 不同小麥品系耐鎘能力對比研究[J]. 土壤與作物,2001,17(4):279-282.
[20]Di B D,Castagnab A,Tognetti R,et al. Early responses to cadmium of two poplar clones that differ in stress tolerance[J]. Journal of Plant Physiology,2014,171(18):1693-1705.
[21]Rehman Z U,Khan S,Brusseau M L,et al. Lead and cadmium contamination and exposure risk assessment via consumption of vegetables grown in agricultural soils of five-selected regions of Pakistan[J]. Chemosphere,2017,168:1589-1596.
[22]Lin L J,Liao M,Lv X L,et al. Addition of straw from hyperaccumulator plants to cadmium-contaminated soil increases cadmium uptake by loquat seedlings[J]. Environmental Monitoring and Assessment,2017,189(5):217.
[23]Zahoor M,Irshad M,Rahman H,et al. Alleviation of heavy metal toxicity and phytostimulation ofBrassicacampestrisL. by endophyticMucorsp. MHR-7[J]. Ecotoxicology and Environmental Safety,2017,142:139-149.
[24]Shahid M,Shamshad S,Rafiq M,et al. Chromium speciation,bioavailability,uptake,toxicity and detoxification in soil-plant system:a review[J]. Chemosphere,2017,178:513-533.
[25]王紀(jì)華,趙春江. 土壤水分對小麥葉片含水量及生理功能的影響[J]. 麥類作物學(xué)報(bào),2001,21(4):42-47.
[26]Yan M L,Liu L L,Wang H H,et al. Accumulation characteristics of cadmium for three plants in red soil[J]. Journal of Agro-Environmental Science,2009,28(1):72-77.
[27]蔡保松,曹林奎. 鎘對小麥生長發(fā)育的影響及其基因型間差異[J]. 西北農(nóng)林科技大學(xué)學(xué)報(bào)(自然科學(xué)版),2003,31(1):62-66.
猜你喜歡
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
紅領(lǐng)巾·萌芽(2016年1期)2016-09-10 07:22:44

- 江蘇農(nóng)業(yè)科學(xué)的其它文章
- PEG預(yù)處理對水分脅迫下水稻根系抗氧化酶同工酶及其表達(dá)的影響
- CBF冷信號通路基因在熱處理誘導(dǎo)的香蕉抗病性中的作用
- 河南省農(nóng)村土地收益保證貸款的農(nóng)戶意愿影響因素分析
——基于合理行動理論(TRA)的估計(jì) - 江蘇省農(nóng)業(yè)現(xiàn)代化發(fā)展水平測算及政策建議
——基于WSR視角 - 社會網(wǎng)絡(luò)對農(nóng)戶智能農(nóng)業(yè)技術(shù)采納的影響
——基于江蘇省13市370戶農(nóng)戶的調(diào)查 - 新生代農(nóng)民工市民化對其工資收入的影響
——基于全國797份問卷調(diào)查數(shù)據(jù)的實(shí)證研究