四溴雙酚A厭氧共代謝降解性能研究
2018-04-25 13:08:39陳英文范夢(mèng)婕孫靖云沈樹(shù)寶石油石化污染物控制與處理國(guó)家重點(diǎn)實(shí)驗(yàn)室北京1006南京工業(yè)大學(xué)生物與制藥工程學(xué)院江蘇南京10009
中國(guó)環(huán)境科學(xué) 2018年4期
陳英文,范夢(mèng)婕,孫靖云,沈樹(shù)寶 (1.石油石化污染物控制與處理國(guó)家重點(diǎn)實(shí)驗(yàn)室,北京 1006;.南京工業(yè)大學(xué)生物與制藥工程學(xué)院,江蘇 南京 10009)
作為一種廣泛應(yīng)用的溴系阻燃劑[1],四溴雙酚 A(TBBPA)在許多生物基質(zhì)中都有檢出[2],對(duì)人類健康具有嚴(yán)重危害[3].目前,TBBPA 的主要降解方法包括吸附法[4],光催化[5]和生物降解法[6].能夠原位修復(fù)污染物的生物降解是目前的研究熱點(diǎn).由于 TBBPA的特殊物化性質(zhì)使其易沉積于底泥沉積物厭氧環(huán)境中,因此研究TBBPA的厭氧降解更具有實(shí)際應(yīng)用價(jià)值.Wang等[7]研究發(fā)現(xiàn) Shewanella sp.XB的黃素分泌依賴于TBBPA的初始濃度,當(dāng)TBBPA初始濃度為80mg/L時(shí),黃素分泌受到了抑制.同時(shí),TBBPA降解產(chǎn)物分析顯示厭氧條件下 TBBPA只能被還原脫溴成雙酚 A(BPA)而不能進(jìn)一步轉(zhuǎn)化[8].因此深入研究 TBBPA厭氧降解途徑具有重要的實(shí)際應(yīng)用價(jià)值.
厭氧共代謝是指通過(guò)微生物代謝其他化合物產(chǎn)生的酶或者輔助因子對(duì)污染物的降解過(guò)程[9-10].很多研究者利用微生物共代謝對(duì)環(huán)境中難降解有機(jī)物進(jìn)行去除,如 4-烷基酚(4-HBA)[11]和磺胺甲惡唑(SMX)[12].與 TBBPA 相同的是,4-HBA與SMX都具有化學(xué)性質(zhì)穩(wěn)定,辛醇/水分配系數(shù)高,脂溶性強(qiáng)等特點(diǎn),因此推測(cè)厭氧共代謝技術(shù)能夠?qū)崿F(xiàn)TBBPA的高效深度降解.
本文基于微生物厭氧共代謝原理,探索TBBPA厭氧共代謝可行性.構(gòu)建厭氧共代謝反應(yīng)器,分析研究 TBBPA 的厭氧共代謝降解效率.通過(guò)微生物特征菌群以及TBBPA降解途徑的研究探討 TBBPA的厭氧共代謝轉(zhuǎn)化機(jī)理,對(duì)研究厭氧環(huán)境中TBBPA深度降解具有重要科學(xué)意義.
1 材料與方法
1.1 反應(yīng)器的構(gòu)建與啟動(dòng)
構(gòu)建兩組反應(yīng)器,分別記為 C-Glu和 C-T,反應(yīng)器為圓柱形有機(jī)玻璃裝置,總?cè)萘繛?50mL.菌株來(lái)源為南京市某污水處理廠,經(jīng)葡萄糖底物溶液馴化,培養(yǎng)并保存.將營(yíng)養(yǎng)液和含有菌株的污泥按照 1:1的比例混合后添加進(jìn)反應(yīng)器中,以碳布作為微生物支撐載體,進(jìn)行馴化掛膜.營(yíng)養(yǎng)液由以下物質(zhì)組成:每升水中含葡萄糖 1.00g, NH4Cl 0.31g, KCl 0.13g,Na2HPO4·12H2O 11.88g, NaH2PO4·2H2O 2.55g,MgSO4·7H2O 0.20g 和微量元素 10mL.每 3d 更換一次營(yíng)養(yǎng)液,當(dāng)反應(yīng)出水溶液TOC低于40mg/L,且持續(xù)3次反應(yīng)循環(huán)時(shí),表明微生物掛膜完成.掛膜完成后,將兩組反應(yīng)器置于恒定溫度(33℃)的分批模式下操作,當(dāng)反應(yīng)出水溶液TOC趨于穩(wěn)定時(shí),更換底物,結(jié)束一次反應(yīng)循環(huán).
1.2 實(shí)驗(yàn)方法
以 1g/L葡萄糖作為反應(yīng)底物加入反應(yīng)器C-Glu;以1g/L葡萄糖作為共基質(zhì),將濃度分別為0,25,50,75,100,200和500μg/L的TBBPA按照葡萄糖溶液與TBBPA溶液體積比為7:3比例混合后依次加入反應(yīng)器 C-T,并根據(jù)葡萄糖在反應(yīng)系統(tǒng)中的降解特點(diǎn),分別在12, 24, 36和60h時(shí)取樣,為了排除數(shù)據(jù)的偶然性,TBBPA的每個(gè)濃度梯度都完成了4次降解反應(yīng).
以200μg/L TBBPA為唯一碳源,在反應(yīng)12h時(shí)取樣并通過(guò)質(zhì)譜分析中間代謝產(chǎn)物;以1g/L葡萄糖和200μg/L TBBPA混合溶液為碳源,在反應(yīng)12h時(shí)取樣并通過(guò)質(zhì)譜分析中間代謝產(chǎn)物.
1.3 分析方法
1.3.1 樣品的檢測(cè)與分析 ①樣品預(yù)處理.從反應(yīng)器中取15mL出水樣品與15mL二氯甲烷混合后,震蕩萃取10min,隨后將萃取液旋蒸至干.用甲醇定容到 3mL,用孔徑為 0.22μm 的有機(jī)系濾膜進(jìn)行過(guò)濾,完成樣品準(zhǔn)備.將處理好的樣品通過(guò)高效液相-質(zhì)譜聯(lián)用儀進(jìn)行檢測(cè).②質(zhì)譜條件.通過(guò)電噴霧電離(ESI)離子源,負(fù)離子模式及多反應(yīng)監(jiān)測(cè)(MRM)對(duì)目標(biāo)產(chǎn)物進(jìn)行定性分析.氣簾氣(CUR),噴霧氣(GS1),輔助加熱氣(GS2)和碰撞氣(CAD)的壓力分別為 206851.8, 241327.1,275802.4和 41370.36Pa,源溫度為 400℃,離子化電壓(IS)為 5500V,母離子峰和子離子峰分別為542.7和417.9,碰撞能(CE),去簇電壓(DP),入口電壓(EP),碰撞室出口電壓(CXP)分別為-53, -75,-7和-10V.③液相色譜條件.在ZORBAX Eclipse Plus C18色譜柱(150mm×2.1mm,3.5μm), 0.02%(V/V)氨水(A)/50%甲醇(B)作流動(dòng)相,柱溫為40℃,進(jìn)樣量為5μL的條件下對(duì)TBBPA進(jìn)行定量分析.測(cè)量時(shí)所采用的流動(dòng)相梯度見(jiàn)表1.

表1 分析TBBPA的流動(dòng)相梯度Table 1 Gradient elution program for the analysis of TBBPA
1.3.2 生物多樣性分析 待反應(yīng)完成后,從不同反應(yīng)體系中的陽(yáng)極生物膜上取生物膜樣品作為檢測(cè)對(duì)象,反應(yīng)器C-Glu中生物膜記為C-Glu,反應(yīng)器C-T中生物膜記為C-T.將生物膜樣品送至上海美吉生物醫(yī)藥科技有限公司在 Illumina平臺(tái)上利用細(xì)菌16S rRNA測(cè)序分析微生物多樣性.
1.3.3 計(jì)算方法 TBBPA的產(chǎn)物通過(guò)質(zhì)譜進(jìn)行分析,TBBPA的濃度通過(guò)高效液相-質(zhì)譜聯(lián)用儀(HPLC-MS)進(jìn)行檢測(cè).TBBPA去除率的計(jì)算方法為:

反應(yīng)一級(jí)速率常數(shù)的計(jì)算公式為
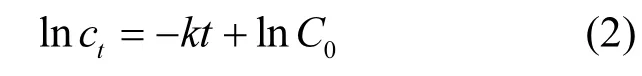
式中:C0為 TBBPA 的初始濃度,μg/L;Ct為TBBPA在反應(yīng)時(shí)間為t時(shí)的濃度,μg/L;k為速率常數(shù).
TBBPA半衰期的計(jì)算公式為
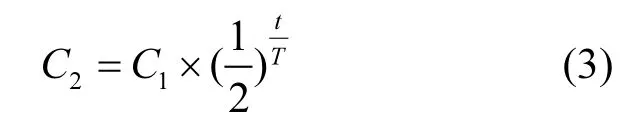
式中:ΔRT為TBBPA的去除率;C1為TBBPA的進(jìn)水濃度,μg/L;C2為 TBBPA 的出水濃度,μg/L;t為反應(yīng)時(shí)間;T為TBBPA的半衰期.
2 結(jié)果與討論
2.1 TBBPA降解效率分析
TOC代表溶液中總有機(jī)碳成分,即溶液中微生物能夠直接代謝的總碳源.在 12h前,溶液TOC快速下降(圖1),TOC去除率達(dá)到60%左右(圖2),12h后溶液TOC下降緩慢(圖1).當(dāng)出水溶液TOC低于40mg/L時(shí),完成一次反應(yīng)循環(huán),即此時(shí)反應(yīng)溶液中的有機(jī)碳源已不足以供給微生物,微生物處于饑餓狀態(tài).當(dāng)?shù)诙螤I(yíng)養(yǎng)底物加入時(shí),微生物的代謝活性能夠迅速提高直至營(yíng)養(yǎng)物被消耗,之后微生物的活性又回復(fù)到較低水平,從底物加入至底物耗盡,微生物經(jīng)歷飽腹至饑餓的循環(huán).因此,12h后直至反應(yīng)完成的TOC下降幅度小于在12h前的下降幅度.由圖1和圖 2亦可知隨著 TBBPA濃度的升高,出水TOC升高,TOC去除率下降,一方面是因?yàn)門BBPA對(duì)微生物的毒性作用[13],一方面是由于TBBPA的物化性質(zhì)與化學(xué)結(jié)構(gòu)使其難于被微生物代謝及轉(zhuǎn)化.
不同濃度TBBPA在厭氧共代謝條件下的降解動(dòng)力學(xué)分析如表2所示.由表2可知,在厭氧共代謝條件下,TBBPA的降解符合一級(jí)動(dòng)力學(xué)方程.而有機(jī)化合物的生物半衰期代表了其在生物條件下的轉(zhuǎn)化及降解速度,TBBPA的生物半衰期及降解時(shí)間隨著 TBBPA濃度的升高而升高,最低生物半衰期為 25μg/L時(shí)的 21.9h,最高生物半衰期為500μg/L時(shí)的64.4h,相比于自然環(huán)境及其它研究者報(bào)道(表 3),厭氧共代謝顯著縮短了TBBPA的生物半衰期.由圖3可知,濃度為75μg/L時(shí)的TBBPA在不同時(shí)間點(diǎn)去除率都高于其他濃度,結(jié)合不同濃度 TBBPA在反應(yīng)完成時(shí)最終去除率都高于75%的分析可知,TBBPA在厭氧共代謝體系中被還原脫溴,進(jìn)一步地,結(jié)合 TOC分析(圖 1和圖 2)可知,在厭氧共代謝體系中 TBBPA很大程度上被共代謝分解而成為微生物可利用的碳源.因此,TBBPA在厭氧共代謝體系中實(shí)現(xiàn)了有效降解.

圖1 不同TBBPA濃度條件下TOC隨時(shí)間變化趨勢(shì)Fig.1 TOC of different concentrations of TBBPA at different time points

圖2 不同TBBPA濃度條件下TOC去除率隨時(shí)間的變化規(guī)律Fig.2 TOC removal rates of different concentrations of TBBPA at different time points

表2 不同濃度下TBBPA的降解動(dòng)力學(xué)研究Table 2 Kinetic results of TBBPA with different conentrations

表3 不同研究所做TBBPA厭氧降解效率對(duì)比Table 3 The comparison of anaerobic degradation efficiency of TBBPA by different researches

圖3 不同TBBPA濃度條件下TBBPA去除率隨時(shí)間的變化規(guī)律Fig.3 The removal rate of TBBPA with different concentrations at different degradation time
2.2 TBBPA厭氧共代謝途徑
已有研究證明在厭氧環(huán)境中 TBBPA只能被還原脫溴為BPA[8],不能進(jìn)一步進(jìn)行生物轉(zhuǎn)化.本文也對(duì)以 TBBPA為單一碳源的厭氧降解進(jìn)行了實(shí)驗(yàn),分析發(fā)現(xiàn) TBBPA的厭氧代謝終產(chǎn)物除了BPA未有其他單環(huán)產(chǎn)物檢出.而厭氧共代謝系統(tǒng)中代謝產(chǎn)物不僅存在BPA(圖4A),還存在單環(huán)產(chǎn)物3,4-二甲氧基苯甲酸甲酯(圖4B).有研究表明苯甲酸和對(duì)羥基苯甲酸都可以被微生物有效降解[17],因此推測(cè)化學(xué)結(jié)構(gòu)相似于苯甲酸和對(duì)羥基苯甲酸的 3,4-二甲氧基苯甲酸甲酯也是一種易于降解的化合物,充分證明了 TBBPA在厭氧共代謝系統(tǒng)中能夠被深度降解.
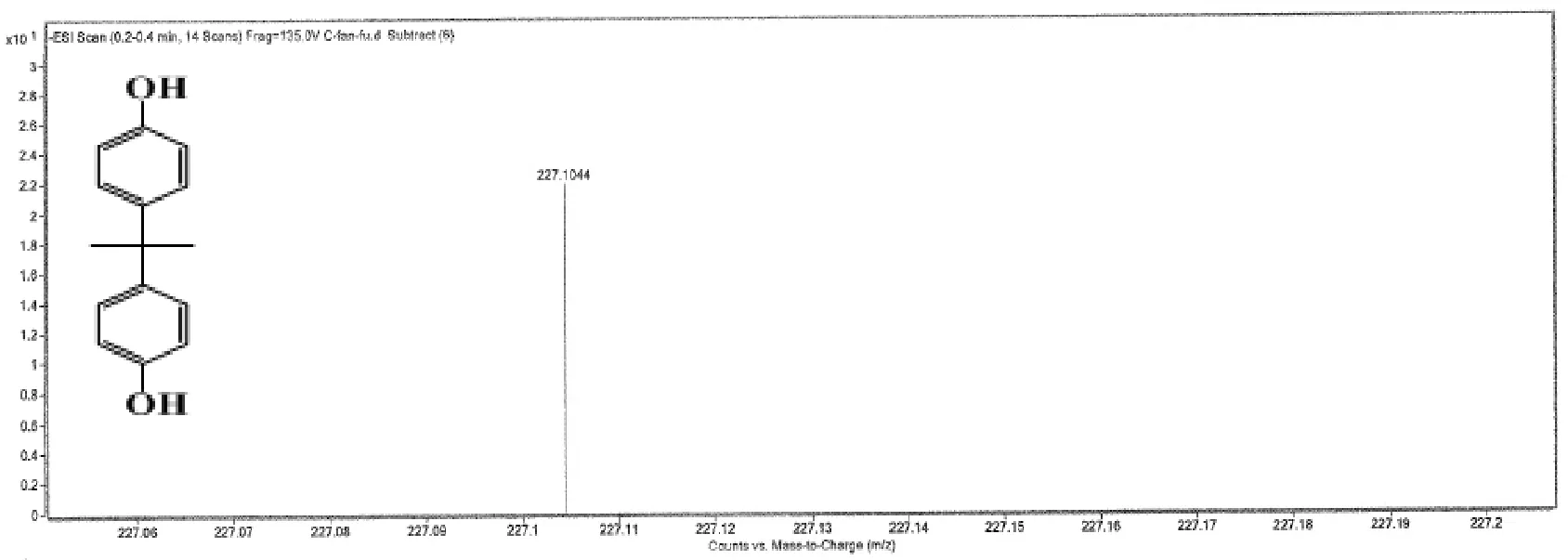

圖4 TBBPA的中間代謝產(chǎn)物Fig.4 The mass spectrometry analysis of TBBPA degradation products
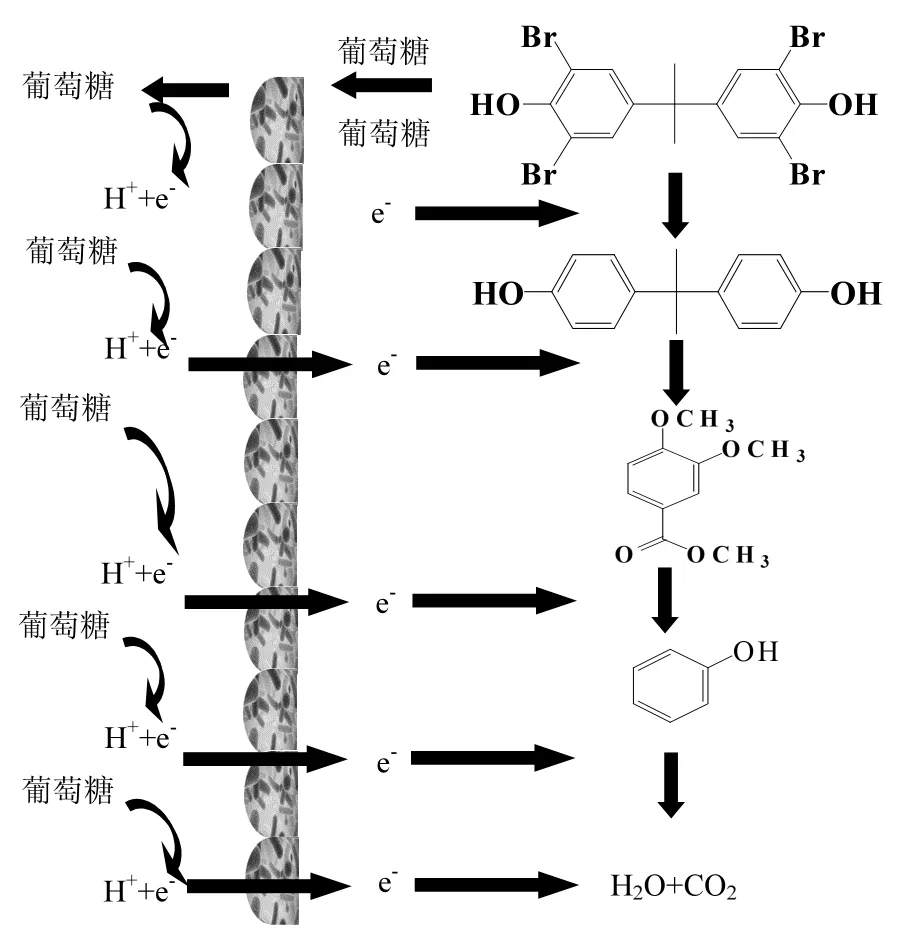
圖5 TBBPA在厭氧共代謝系統(tǒng)中的降解途徑Fig.5 A proposed degradation pathway of TBBPA in conventional co-metabolic reactor
作為 TBBPA 厭氧共代謝的共基質(zhì),葡萄糖一方面緩解 TBBPA對(duì)微生物的毒性[18],另一方面為TBBPA 的分解及轉(zhuǎn)化提供質(zhì)子和電子.厭氧還原脫鹵作用是芳香族化合物在環(huán)境中礦化的關(guān)鍵步驟[19].由圖4可知,在厭氧共代謝條件下,TBBPA首先厭氧還原脫溴成 BPA,隨后通過(guò)氧化,分解及其它共代謝協(xié)同作用將BPA轉(zhuǎn)化成單環(huán)產(chǎn)物3,4-二甲氧基苯甲酸甲酯,并最終將TBBPA轉(zhuǎn)化成H2O和CO2,實(shí)現(xiàn)TBBPA的深度降解(圖5).
當(dāng)以葡萄糖為共代謝底物時(shí),不僅檢測(cè)到了雙環(huán)產(chǎn)物雙酚A,還檢測(cè)到了TBBPA的單環(huán)產(chǎn)物.結(jié)合 TBBPA的生物半衰期(表 2)和反應(yīng)結(jié)束時(shí)不同濃度下 TBBPA 去除率(圖 3)分析可知,TBBPA在反應(yīng)體系中是被共代謝降解而非其他代謝方式.
2.3 微生物特征分析
在Illumina平臺(tái)通過(guò)16S rRNA對(duì)反應(yīng)完成后的菌群進(jìn)行微生物多樣性分析,確定代謝轉(zhuǎn)化TBBPA的特征菌群.香農(nóng)指數(shù)被認(rèn)為是微生物多樣性指數(shù),香農(nóng)曲線越平緩表明測(cè)序數(shù)據(jù)量足夠大,能反映微生物樣品中的大部分信息[20],C-Glu反應(yīng)體系和 C-T反應(yīng)體系中香農(nóng)指數(shù)分別為3.46和3.60,這表明TBBPA的加入引起了C-Glu和C-T兩個(gè)樣品之間微生物多樣性的差異.
由圖6可知,在厭氧共代謝系統(tǒng)中,只以葡萄糖為降解底物時(shí),Blvii28_wastewater-sludge和PHOS-HE36兩種菌株為主要菌屬, Blvii28_wastewater-sludge通常在厭氧廢水或淡水環(huán)境中,是污泥中的典型菌屬[21],PHOS-HE36是一種能夠以葡萄糖作為碳源的厭氧菌屬[22].而相較于厭氧共代謝系統(tǒng),TBBPA厭氧共代謝體系中固氮弧菌屬和毛球菌屬所占豐度比例分別為 11%和19%.這種現(xiàn)象的發(fā)生說(shuō)明TBBPA的加入促使固氮弧菌屬和毛球菌屬豐度增加,成為了C-T反應(yīng)體系中的優(yōu)勢(shì)菌屬.固氮弧菌屬屬于變形菌門,是一種兼性厭氧產(chǎn)電菌,最主要的兩個(gè)特征是利用有機(jī)碳作為碳源及對(duì)氮的固定[23].近年來(lái),很多研究者集中于固氮弧菌屬對(duì)苯系物(BTEX)降解能力的研究[24].另外,研究者發(fā)現(xiàn)固氮弧菌屬可以有效降解高效氯氰菊酯[25].毛球菌屬屬于厚壁菌門,是一種常從污泥中分離出來(lái)的兼性厭氧菌,能夠在厭氧反應(yīng)器中處理有機(jī)廢水[26].同時(shí),Sun等[27]發(fā)現(xiàn)毛球菌屬具有能夠降解剛果紅的能力,而剛果紅為一種極難生物降解的聯(lián)苯胺類偶氮染料.因此,結(jié)合 TBBPA 降解途徑,TBBPA 降解效率分析,可以推測(cè)固氮弧菌屬和毛球菌屬是厭氧共代謝降解TBBPA的特征菌屬.

圖6 微生物群落組成圖(屬)Fig.6 The composition of microbial community (genus)
3 結(jié)論
厭氧共代謝降解 TBBPA研究表明,在 12h時(shí),溶液TOC去除率達(dá)到60%左右,最低生物半衰期為 25μg/L時(shí)的 21.9h,最高生物半衰期為500μg/L時(shí)的64.4h,濃度為75μg/L時(shí)的 TBBPA去除率最高,且不同濃度 TBBPA的去除率都高于70%.微生物群落結(jié)構(gòu)及TBBPA中間代謝產(chǎn)物分析表明固氮弧菌屬和毛球菌屬為代謝TBBPA的特征菌屬.
參考文獻(xiàn):
[1]Zhou Q, Xing A, Zhao D, et al. Tetrabromobisphenol A photoelectrocatalytic degradation using reduced graphene oxide and cerium dioxide comodified TiO2, nanotube arrays as electrode under visible light [J]. Chemosphere, 2016,165:268-276.
[2]Hlou?ková V, Lanková D, Kalachová K, et al. Occurrence of brominated flame retardants and perfluoroalkyl substances in fish from the Czech aquatic ecosystem [J]. Science of the Total Environment, 2013,461-462(461-462C):88-98.
[3]Birnbaum L S, Staskal D F. Brominated flame retardants: cause for concern? [J]. Environmental Health Perspectives, 2004,112(1):9-17.
[4]Yang J, Li J Y, Qiao J Q, et al. Magnetic solid phase extraction of brominated flame retardants and pentachlorophenol from environmental waters with carbon doped Fe3O4, nanoparticles [J].Applied Surface Science, 2014,321(321):126-135.
[5]Bao Y, Niu J. Photochemical transformation of tetrabromobisphenol A under simulated sunlight irradiation:Kinetics, mechanism and influencing factors [J]. Chemosphere,2015,134:550-556.
[6]Li F, Wang J, Nastold P, et al. Fate and metabolism of tetrabromobisphenol A in soil slurries without and with the amendment with the alkylphenol degrading bacterium Sphingomonas sp. strain TTNP3 [J]. Environmental Pollution,2014,193(10):181-188.
[7]Wang J, Fu Z, Liu G, et al. Mediators-assisted reductive biotransformation of tetrabromobisphenol-A by Shewanella sp.XB [J]. Bioresource Technology, 2013,142(8):192-197.
[8]Chang B V, Yuan S Y, Ren Y L. Anaerobic degradation of tetrabromobisphenol-A in river sediment [J]. Ecological Engineering, 2012,49(12):73-76.
[9]Luo W, Zhao Y, Ding H, et al. Co-metabolic degradation of bensulfuron-methyl in laboratory conditions [J]. Journal of Hazardous Materials, 2008,158(1):208-214.
[10]Moreira I S, Amorim C L, Carvalho M F, et al. Co-metabolic degradation of chlorobenzene by the fluorobenzene degrading wild strain Labrys portucalensis [J]. International Biodeterioration& Biodegradation, 2012,72(4):76-81.
[11]Shibata A, Katayama A. Anaerobic co-metabolic oxidation of 4-alkylphenols with medium-length or long alkyl chains by Thauera sp., strain R5 [J]. Applied microbiology and biotechnology, 2007,75(5):1151-1161.
[12]Müller E, Schüssler W, Horn H, et al. Aerobic biodegradation of the sulfonamide antibiotic sulfamethoxazole by activated sludge applied as co-substrate and sole carbon and nitrogen source [J].Chemosphere, 2013,92(8):969-978.
[13]Arbeli Z, Ronen Z, Díaz-Báez M C. Reductive dehalogenation of tetrabromobisphenol-A by sediment from a contaminated ephemeral streambed and an enrichment culture [J]. Chemosphere 2006,64(9):1472-1478.
[14]Voordeckers J W, Fennell D E, Jones K, et al. Anaerobic biotransformation of tetrabromobisphenol A, tetrachlorobisphenol A, and bisphenol A in estuarine sediments [J]. Environmental Science & Technology, 2002,36(4):696-701.
[15]Chang B V, Yuan S Y, Ren Y L. Anaerobic degradation of tetrabromobisphenol-A in river sediment [J]. Ecological Engineering, 2012,49(12):73-76.
[16]Li G Y, Xiong J K, Wong P K, et al. Enhancing tetrabromobisphenol A biodegradation in river sediment microcosms and understanding the corresponding microbial community [J]. Environmental Pollution, 2016,208:796-802.
[17]毛 寧,薛泉宏,唐 明.2株放線菌對(duì)土壤中苯甲酸和對(duì)羥基苯甲酸的降解作用 [J]. 西北農(nóng)林科技大學(xué)學(xué)報(bào):自然科學(xué)版,2010,38(5):143-148.
[18]Peng X, Qu X, Luo W, et al. Co-metabolic degradation of tetrabromobisphenol A by novel strains of Pseudomonas sp. and Streptococcus sp [J]. Bioresource Technology, 2014,169:271-276.
[19]Ronen Z, Abeliovich A. Anaerobic-aerobic process for microbial degradation of tetrabromobisphenol A [J]. Applied &Environmental Microbiology, 2000,66(6):2372-2377.
[20]Wang Y, Sheng H F, He Y, et al. Comparison of the Levels of Bacterial Diversity in Freshwater, Intertidal Wetland, and Marine Sediments by Using Millions of Illumina Tags [J]. Applied &Environmental Microbiology, 2012,78(23):8264-8271.
[21]蔣 容.生物抑制劑在降低聚合物溶液黏度損失中的應(yīng)用 [J].石油規(guī)劃設(shè)計(jì), 2016,27(4):19-22.
[22]Zheng J, Xiao X, Zhang Q, et al. The Placental Microbiota Is Altered among Subjects with Gestational Diabetes Mellitus: A Pilot Study [J]. Frontiers in physiology, 2017,8:675.
[23]Hurek T, Reinhold-Hurek B. Azoarcus sp. strain BH72as a model for nitrogen-fixing grass endophytes [J]. Journal of Biotechnology, 2003,106(2/3):169-178.
[24]Rather L J, Bill E, Ismail W, et al. The reducing component BoxA of benzoyl-coenzyme A epoxidase from Azoarcus evansii is a[4Fe-4S]protein [J]. Biochimica et Biophysica Acta,2011,1814(12):1609-1615.
[25]Ma Y, Chen L, Qiu J. Biodegradation of beta-cypermethrin by a novel Azoarcus indigens strain HZ5 [J]. Journal of Environmental Science and Health, Part B, 2013,48(10):851-859.
[26]Garcia-Mancha N, Puyol D, Monsalvo V M, et al. Anaerobic treatment of wastewater from used industrial oil recovery [J].Journal of Chemical Technology and Biotechnology,2012,87(9):1320-1328.
[27]Sun J, Li Y, Hu Y, et al. Understanding the degradation of Congo red and bacterial diversity in an air–cathode microbial fuel cell being evaluated for simultaneous azo dye removal from wastewater and bioelectricity generation [J]. Applied Microbiology and Biotechnology, 2013,97(8):3711-3719.
