花期增溫對香梨幼果期果實同化物積累及坐果率和萼片脫落的影響
2018-04-25 00:42:28努爾麥麥提艾麥提覃偉明夏熱帕提艾則孜艾力江麥麥提李允齊曼尤努斯
新疆農業科學 2018年12期
努爾麥麥提·艾麥提,覃偉明,夏熱帕提·艾則孜,艾力江·麥麥提,李允,齊曼·尤努斯
(1.新疆農業大學林學與園藝學院,烏魯木齊 830052;2.新疆巴音郭楞蒙古自治州沙依東園藝場,新疆庫爾勒 841000)
0 引 言
【研究意義】新疆庫爾勒香梨歷史悠久,品質上佳,是著名的區域特有品牌,在新疆林果業發展中具有重要地位[1]。庫爾勒香梨不僅成了當地農村經濟發展和農民增收的優勢產品,而且也成為巴州地區經濟可持續發展的支柱產業[2]。南疆獨特的自然氣候條件和地理位置非常適合果樹的發展,是香梨、蘋果、紅棗、核桃生產的適宜區,巴州地區、阿克蘇地區是庫爾勒香梨發展的最佳區域,表現出香梨自身的優良特性[3-4],但通過對新疆南疆盆地近45 a氣溫變化特征分析發現,南疆盆地年平均氣溫在1989年開始發生暖突變。從1980年代末夜間溫度開始異常升高,白天溫度也隨之升高,導致南疆盆地整體溫度異常暖突變[5]。作為自然生態系統重要組成部分的地表植物必將受到氣候變暖的影響,研究花期溫度變化對庫爾勒香梨萼片和果實脫落的影響機制,對改善庫爾勒香梨栽培管理并提高商品率及經濟效益有實際意義。【前人研究進展】目前,丁家鳴和Choi J J等學者已對梨樹花期溫度或者花后溫度變化,對梨果實萼片和果實脫落影響研究工作進行了初步報道[6-7]。廖明康[8]研究表明,庫爾勒香梨果實形成突頂果的原因與樹體養分平衡有密切關系,而養分平衡又與合理的負載量、施肥及修剪有關。齊笑笑等[9]對庫爾勒香梨果實進行高通量測序發現,與花萼脫落相關的差異表達基因主要涉及光合作用、植物激素信號轉導、細胞壁修飾、轉錄調控和糖代謝。研究表明,同化物對蠶豆[10-11]、棉花[12]、柑橘[13-14]、葡萄[15]、荔枝[16-17]、蘋果[18]等植物器官脫落均有調節作用。溫度是影響植物同化物積累與分配的重要環境因子[19],而同化物作為底物用于維持組織的生長發育,花期溫度變化能否影響香梨幼果實同化物積累,進而影響果實和萼片的脫落等問題值得研究。【本研究切入點】坐果始期是落花落果嚴重的關鍵時期,也是香梨萼片脫落的關鍵前期。通過改變花期溫度研究同化物在萼片離層形成期前后果實中的積累以及與萼片脫落和果實坐果率的相關性,分析梨果實在坐果始期、萼片離層形成期和萼片脫落期幼果實同化物動態變化并對萼片脫落與宿存的影響,為萼片和果實器官脫落機制研究提供參考依據。研究香梨花期溫度變化,對同化物在萼片離層形成期前后果實中的積累以及對萼片脫落和果實坐果率的影響。【擬解決的關鍵問題】通過改變庫爾勒香梨不同花期小氣候環境,研究在萼片離層形成期前后果實中同化物動態變化與萼片脫落和果實坐果率之間的關系,為萼片和幼果器官脫落機制研究提供科學依據。
1 材料與方法
1.1 材 料
選取新疆庫爾勒沙依東園藝場常規管理的10年生的庫爾勒香梨樹,并在初花期和盛花期期間(2017年4月6~9日和2017年4月9~14日),對果園內同等樹齡和樹勢的香梨樹木個體進行增溫處理。在坐果始期、萼片離層形成期和萼片脫落期對幼果實進行采樣并利用液氮保存作為待測樣品。
1.2 方 法
1.2.1 試驗設計
分別設置初花期和盛花期增溫處理并設立對照組(CK),在香梨初花期(2017年4月6~9日)和盛花期期間(2017年4月9~14日),利用鋼管架和溫室大棚薄膜圍起香梨樹,在當地時間20:00~08:00放置加熱器加熱。初花期白天大棚薄膜半打開,盛花期全開,使香梨樹體正常接收日照,同時處理期間用自動溫濕度計記錄溫度變化。
1.2.2 糖含量測定
可溶性糖和淀粉測定采用蒽酮比色法[20],果糖、蔗糖測定采用間苯二酚法[21]。
1.2.3 脫萼果率和宿萼果率統計
在生理落果期后統計各處理及對照的脫萼果率,花序和花朵坐果率。
1.3 數據處理
所得試驗數據均用Excel 軟件制圖和數據整理,SPSS19.0統計軟件進行數據分析。
2 結果與分析
2.1花期自然溫度和花期增溫處理溫度記錄
研究表明,初花期期間自然條件下,晝夜溫差小,對初花期梨樹晝夜小氣候進行增溫處理,提高晝夜溫度;盛花期溫度變化幅度大,晝夜溫差大,夜間出現最低溫2.5℃,對盛花期夜間小氣候進行了增溫處理。圖1
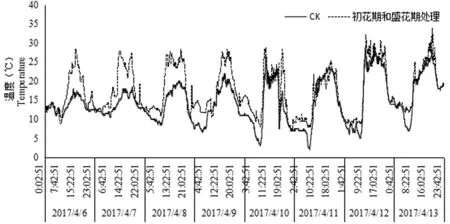
圖1 花期自然溫度處理和和花期增溫處理溫度
Fig.1 temperature record of control and warming treatment at flowering stage
2.2 花期增溫處理對香梨脫萼率和坐果率影響
研究表明,花期增溫處理后脫萼率均下降,初花期處理脫萼率與對照脫萼率無顯著差異,盛花期處理后脫萼率極顯著性下降,降到20.95%,比對照降低了27.11%(圖2A)。花期增溫處理后花序坐果率均上升,但初花期處理花朵坐果率下降,而盛花期處理花朵坐果率增加(P>0.05)(圖2B)。圖2
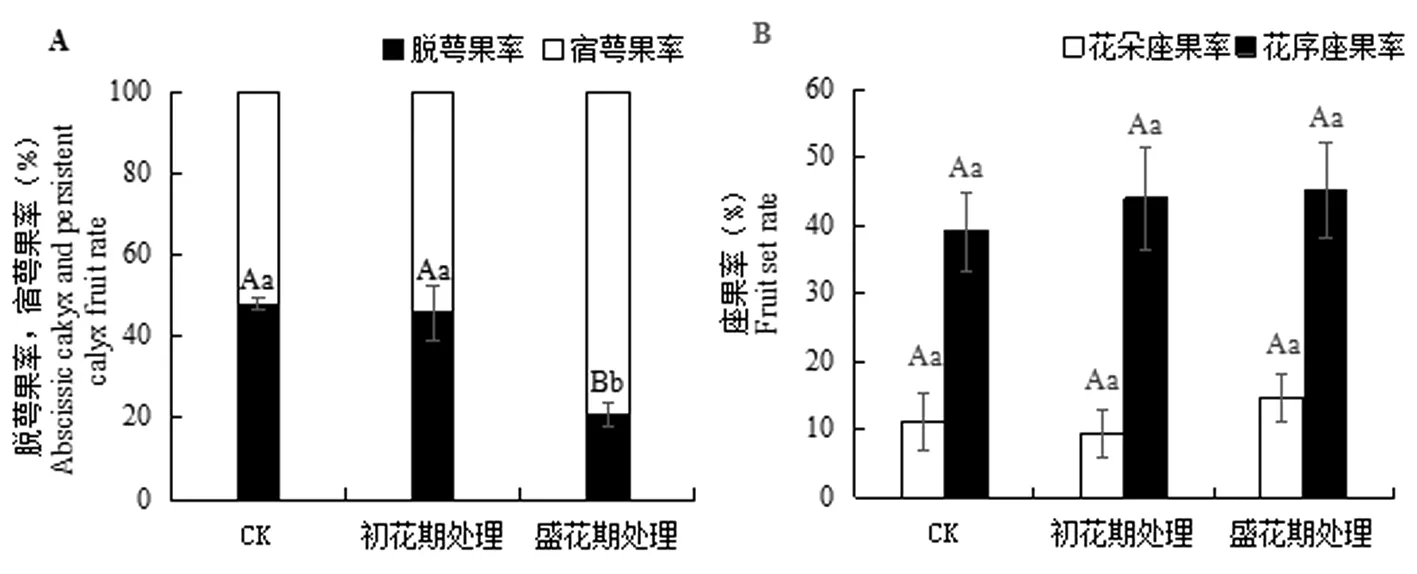
圖2 花期增溫處理下香梨脫萼率和坐果率變化
Fig.2 Effect of warming treatments on the calyx abscission rate and fruit setting rate of korla fragrant pear
2.3 花期增溫處理對坐果始期香梨果實同化物積累的影響
研究表明,盛花期增溫處理后坐果始期果實可溶性糖含量顯著增加,初花處理后降低,但初花處理與對照差異不顯著;初花期和盛花期處理后蔗糖和淀粉含量均增加,并且在盛花處理后蔗糖含量增加顯著,相比對照高2.8倍。表1
表1 花期增溫處理下坐果始期香梨果實同化物積累變化
Table 1 Effects of warming treatments on assimilate accumulation in pear fruit at the beginning of fruit setting stage
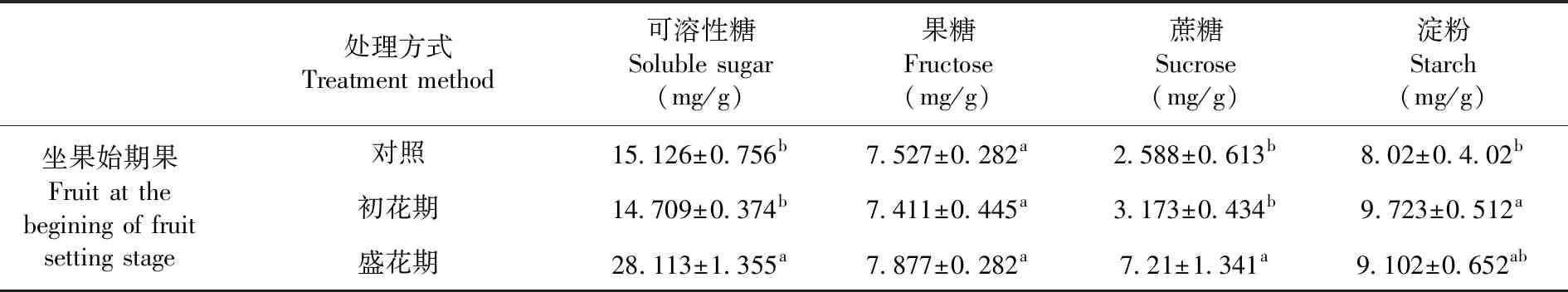
處理方式Treatment method可溶性糖Soluble sugar(mg/g)果糖Fructose(mg/g)蔗糖Sucrose(mg/g)淀粉Starch(mg/g)坐果始期果Fruit at the begining of fruit setting stage對照15.126±0.756b7.527±0.282a2.588±0.613b8.02±0.4.02b初花期14.709±0.374b7.411±0.445a3.173±0.434b9.723±0.512a盛花期28.113±1.355a7.877±0.282a7.21±1.341a9.102±0.652ab
注:不同小寫字母分別表示差異顯著(P<0.05)
Note: The small letters represent significant difference at 0.05
2.4 花期增溫處理對離層形成期香梨果實同化物積累的影響
研究表明,初花期和盛花期處理后在萼片離層形成期離層形成果實可溶性糖,果糖和蔗糖含量均顯著降低,淀粉含量顯著增加,初花期處理后蔗糖含量最低為0.54 mg/g。未形成離層果實果糖和淀粉含量增加,蔗糖含量降低。在萼片離層形成期各處理組未形成離層果實可溶性糖、蔗糖和淀粉含量均高于離層形成果,而對照和盛花期處理組未形成離層果實果糖含量低于離層形成果。表2
2.5 花期增溫處理對萼片脫落期香梨果實同化物積累的影響
研究表明,盛花期增溫處理后萼片脫落期脫萼果可溶性糖和蔗糖含量均高于對照,而初花期增溫處理后可溶性糖、蔗糖和淀粉含量比對照均顯著性下降,分別為9.321、2.169和2.75 mg/g。花期增溫處理組宿萼果中可溶性糖,蔗糖和淀粉含量均顯著性下降,而脫萼果和宿萼果果糖含量均增加。脫萼果相比,各處理宿萼果果糖和淀粉含量均高,同時初花期增溫處理和對照蔗糖和可溶性糖含量也均高于脫萼果,而盛花期增溫處理后低于脫萼果。表3
表2 花期增溫處理下離層形成期香梨果實同化物積累變化
Table 2 Effect of warming treatments on assimilate accumulation in Pear Fruit during abscission zone formation stage
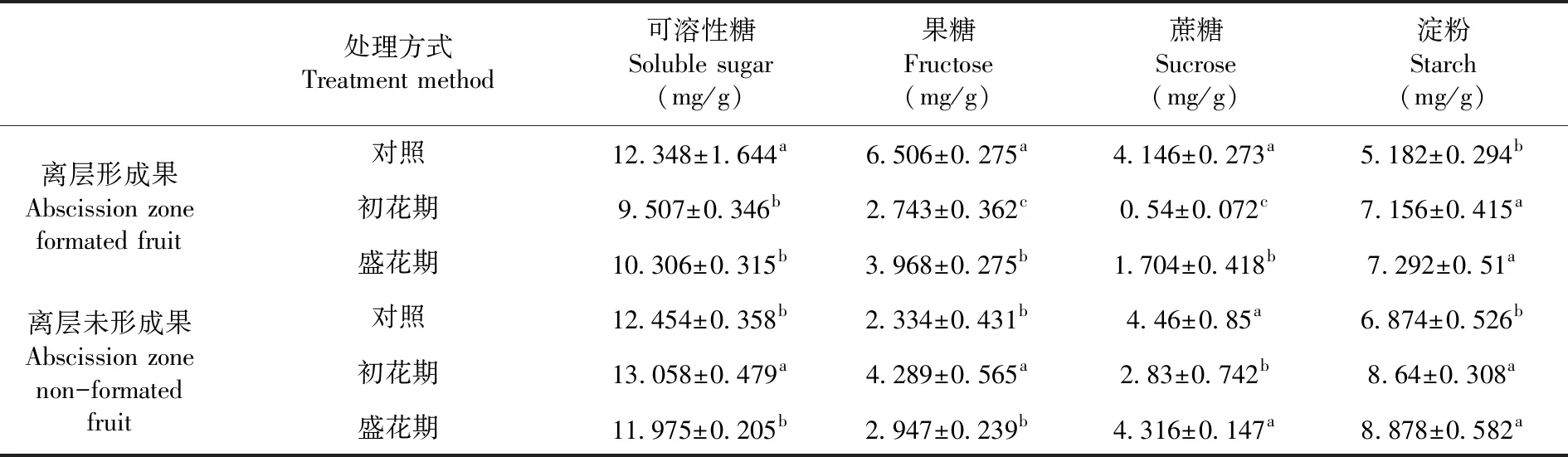
處理方式 Treatment method可溶性糖Soluble sugar(mg/g)果糖Fructose(mg/g)蔗糖Sucrose(mg/g)淀粉Starch(mg/g)離層形成果Abscission zone formated fruit對照12.348±1.644a6.506±0.275a4.146±0.273a5.182±0.294b初花期9.507±0.346b2.743±0.362c0.54±0.072c7.156±0.415a盛花期10.306±0.315b3.968±0.275b1.704±0.418b7.292±0.51a離層未形成果Abscission zone non-formated fruit對照12.454±0.358b2.334±0.431b4.46±0.85a6.874±0.526b初花期13.058±0.479a4.289±0.565a2.83±0.742b8.64±0.308a盛花期11.975±0.205b2.947±0.239b4.316±0.147a8.878±0.582a
注:不同小寫字母分別表示差異顯著(P<0.05)
Note: The small letters represent significant difference at 0.05
表3 花期增溫處理下萼片脫落期香梨果實同化物積累變化
Table 3 Effects of warming treatments on assimilate accumulation of fragrant pear fruit during the calyx abscission stage
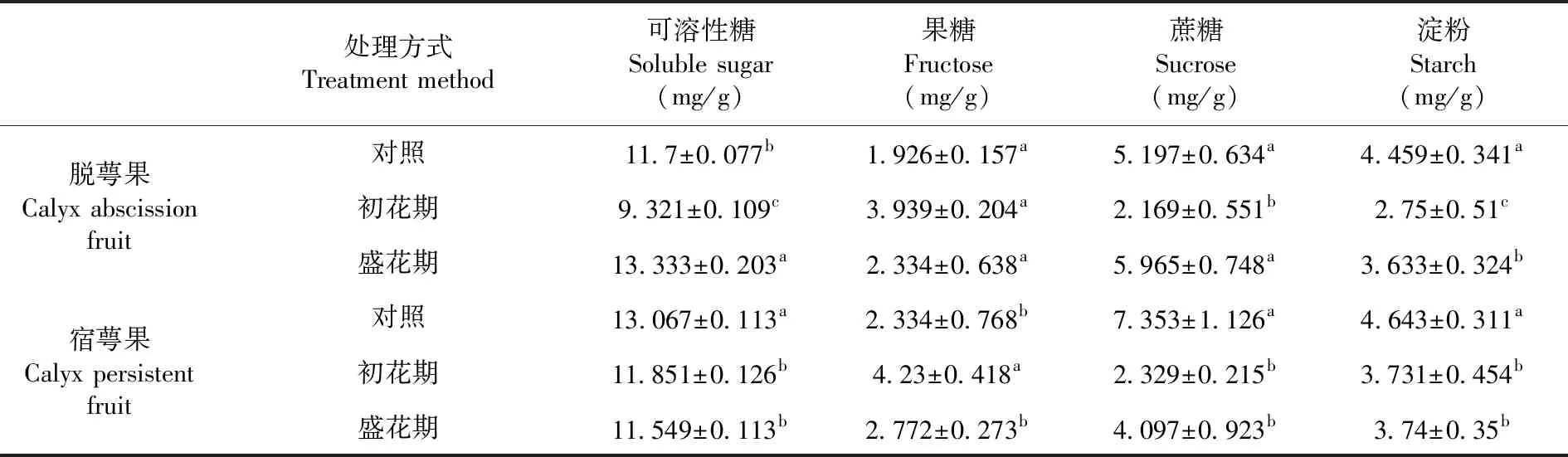
處理方式 Treatment method可溶性糖Soluble sugar(mg/g)果糖Fructose(mg/g)蔗糖Sucrose(mg/g)淀粉Starch(mg/g)脫萼果Calyx abscission fruit 對照11.7±0.077b1.926±0.157a5.197±0.634a4.459±0.341a初花期9.321±0.109c3.939±0.204a2.169±0.551b2.75±0.51c盛花期13.333±0.203a2.334±0.638a5.965±0.748a3.633±0.324b宿萼果Calyx persistent fruit對照13.067±0.113a2.334±0.768b7.353±1.126a4.643±0.311a初花期11.851±0.126b4.23±0.418a2.329±0.215b3.731±0.454b盛花期11.549±0.113b2.772±0.273b4.097±0.923b3.74±0.35b
注:不同小寫字母分別表示差異顯著(P<0.05)
Note: The small letters represent significant difference at 0.05
3 討 論
植物器官(葉、花和果實等)的脫落除了受環境因素影響外,還受植物體內在因素如相關基因表達、內源激素代謝和養分、能量供應等綜合影響[22]。馬宏超等[23]研究表明,GA3處理的庫爾勒香梨幼果萼筒具有發育良好的導管和篩管,萼筒內側基本組織中還形成了大量的含有單寧的異細胞,保證了花萼中養分和水分的供應,使幼果萼片細胞發育正常,后期不會形成離層;而pp333處理的幼果萼筒在發育過程中維管束只有導管,未見有篩管和異細胞的形成,維管束結構不完整且面積小,使花萼中水分和養料的供給缺乏,導致幼果萼筒在發育后期出現了離層,形成了脫萼果。研究結果表明,與對照相比,香梨不同花期增溫處理均能降低脫萼率,盛花期增溫處理的脫萼率差異極顯著,這與Choi J J等[6]的研究結果基本一致。在坐果始期,果實可溶性糖含量在盛花期處理后極顯著增加,初花期處理后無顯著變化,而在蔗糖含量在初花期和盛花期增溫處理后均上升,并且在萼片離層形成期離層形成的果實可溶性糖,蔗糖和淀粉含量均低于未形成離層果,在萼片脫落期宿萼果淀粉含量均高于脫萼果。說明盛花期處理可能增加運輸到果實的糖含量,引起果實同化物積累再往萼片運輸,使萼片繼續生長不掉落,而對照較低溫度條件下同化物供應不能滿足萼片需要,進而導致脫落[10]。
蘇柳蕓等[24]表明,花期氣溫高,梨花開放的快,花期短,受粉時間相應縮短;由于空氣干燥,柱頭上粘液里的水分迅速蒸發,不利于花粉附著和花粉粒萌發,從而不能受精導致坐果差。研究結果表明,初花期處理花朵坐果率降低,而盛花期處理花朵坐果率增加,但各處理組間坐果率差異均不顯著。同時從萼片離層形成期前后同化物含量變化動態得知,可溶性糖、果糖和淀粉含量總體上呈下降趨勢,說明香梨葉片還未完全展開,樹體儲存營養已被果實和葉片生長發育所消耗,這可能是引起生理落果的主要原因,而且初花期處理后坐果始期,萼片離層形成期和萼片脫落期可溶性糖含量總體上均低,同時初花期處理坐果率也均低,說明同化物含量的匱乏引起坐果率降低,該結果與蘇柳蕓等[24]研究報道相似,其研究結果認為,若碳水化合物充足則坐果率高。
木合塔爾·扎熱[25]研究發現,長勢較強的庫爾勒香梨樹的花序花朵數量的多少與萼片宿存和脫落之間沒有任何相關性,無論是人工授粉還是未人工授粉,疏花或未疏花,全部處理的果實均為宿萼果。研究結果發現,在庫爾勒香梨萼片離層形成期前期同化物充足雖然導致宿萼果的形成,但提高坐果率,研究并能實現提高脫萼率和坐果率是在香梨高質栽培的關鍵問題。玉山·庫爾班等[26]研究證明盛花期噴施復合試劑M1,能夠顯著提高庫爾勒香梨脫萼率和坐果率,改善果實品質。
4 結 論
不同花期增溫處理對果實同化物積累的影響程度不同。初花期增溫處理對庫爾勒香梨果實坐果始期同化物的積累和脫萼率無明顯差異,但總體上同化物含量較低,且坐果率無明顯降低,而盛花期增溫處理后坐果始期果實同化物的累積明顯增加并脫萼率顯著降低,但坐果率增加,與對照和初花期處理沒有顯著差異。在萼片離層形成期未形成離層果實可溶性糖、蔗糖和淀粉含量均高于離層形成果。盛花期溫度升高促進離層形成前期同化物積累,進一步導致萼片宿存,但對坐果率影響不明顯。在生產中可以通過合理栽植、施肥與噴施生長調節劑、修剪、疏花疏果和合理安排防護林等措施改變樹體養分狀況和果園微氣候條件,進而能提高庫爾勒香梨脫萼率和坐果率,減少氣候變化帶來的經濟損失。
