一種新的真核DNA表觀遺傳標記
——6mA
2018-03-04 06:47:06涂艷陽空軍軍醫(yī)大學(xué)唐都醫(yī)院實驗外科陜西西安710038
轉(zhuǎn)化醫(yī)學(xué)電子雜志 2018年12期
關(guān)鍵詞:生物
祁 婧,涂艷陽 (空軍軍醫(yī)大學(xué)唐都醫(yī)院實驗外科,陜西西安710038)
0 引言
基因在某種程度上決定大多數(shù)常見疾病的風(fēng)險[1-3],同時也不排除環(huán)境因素等一系列其他的因素,而其中表觀遺傳學(xué)在個體的遺傳因素和環(huán)境之間提供了一個機械的交叉點[4-6],那么通過找出該交叉口控制分子機制就有可能探究出疾病發(fā)生的秘密,以及采取具體的直接的治療方案來影響這一過程并改變其中的疾病風(fēng)險[7]。目前所報道的,與表觀遺傳學(xué)相關(guān)的疾病主要有腫瘤、心血管病、精神病和自身免疫系統(tǒng)性病等[8-9]。
DNA甲基化屬于表觀遺傳學(xué)[1-2]范疇,是其研究中最深入的方向。DNA甲基化是最常見的復(fù)制后及轉(zhuǎn)錄后修飾方式之一,它不改變DNA一級結(jié)構(gòu),在細胞生長分化、基因表達模式以及基因組的穩(wěn)定性中起著至關(guān)重要的作用。DNA甲基化是目前研究最為廣泛的表觀遺傳現(xiàn)象。在真核生物體內(nèi),DNA甲基化是一種酶介導(dǎo)的化學(xué)修飾,在DNA甲基轉(zhuǎn)移酶(DNA methyltransferase,DNMTs)的作用下,以 S-腺苷甲硫氨酸(S-adenosyl methionine,SAM)為甲基供體,將甲基基團轉(zhuǎn)移到胞嘧啶和鳥嘌呤雙核苷酸(CpG島或稱CpG)的胞嘧啶上,當然生物亦能通過屏蔽甲基轉(zhuǎn)移酶的作用實現(xiàn)去甲基化[10-11]。
2015年,三項研究發(fā)現(xiàn)在三種不同的真核生物中6mA獨立存在,揭示了這種甲基化修飾在真核生物中的功能[12-14]。通過利用和開發(fā)幾種高通量測序方法在Chlamydomonas基因組DNA中繪制6mA,顯示6mA不僅富含轉(zhuǎn)錄起始位點(TSS)周圍的ApT二核苷酸,還標記了活性轉(zhuǎn)錄基因,并標記了接頭相鄰核小體之間的DNA區(qū)域,表明衣藻基因組中腺嘌呤甲基化的潛在基因激活功能(圖 1B)[15-16]。
1 m6A的發(fā)現(xiàn)
最初N6-甲基腺嘌呤(6mA或 m6dA)在細菌DNA中被發(fā)現(xiàn),參與DNA修復(fù)、復(fù)制和細胞防御。20世紀50年代Dunn等[17]在探究胸腺胞嘧啶(T)的結(jié)構(gòu)類似物對需T菌株的影響時,將DNA水解后利用二維紙層析方法進行堿基分離時,發(fā)現(xiàn)了第5個紫外吸收光點 X——6mA。然而6mA目前在萊茵衣藻、秀麗隱桿線蟲、果蠅、真菌等真核生物中發(fā)現(xiàn)了DNA-6mA的存在。然而,真核系統(tǒng)中6mA的功能仍然沒有得到完全揭示。m6A的廣泛存在得到研究人員越來越多的關(guān)注,從而針對其生物學(xué)意義和重要性進行更深入的研究。
隨著免疫沉淀、限制酶切、LC-MS/MS以及生物測序等生物技術(shù)的發(fā)展,研究人員在秀麗隱桿線蟲中發(fā)現(xiàn)6mA,其去甲基化酶NMAD-1和潛在的甲基轉(zhuǎn)移酶DAMT-1改變了之前的觀點,即秀麗隱桿線蟲缺乏DNA甲基化,提出了6mA作為DNA甲基化標記而不是5mC的可能性。nmad-1和damt-1缺失的表型以及腺嘌呤甲基化和組蛋白修飾之間的串擾表明6mA 的潛在基因激活作用(圖1C)[18]。
通過敲除果蠅基因組中的去甲基化酶候選物并監(jiān)測6mA水平,Zhang等發(fā)現(xiàn)果蠅Tet同源物可能是果蠅基因組中6mA去甲基化的原因。鑒定的DNA 6mA去甲基化酶(DMAD)在胚胎發(fā)生和組織穩(wěn)態(tài)過程中調(diào)節(jié)6mA的水平。進一步的測序分析表明,動態(tài)去甲基化與轉(zhuǎn)座子表達相關(guān),并在發(fā)育中起關(guān)鍵作用(圖 1D)[16]。
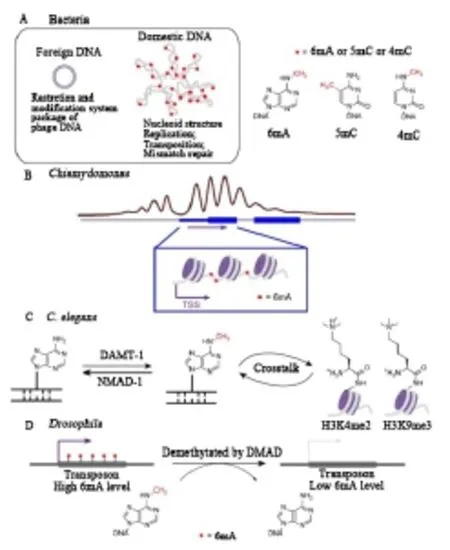
圖1
2 m6A的普遍
近年來,隨著測序技術(shù)的不斷發(fā)展,人們已經(jīng)在萊茵衣藻、秀麗隱桿線蟲、果蠅、真菌等真核生物中發(fā)現(xiàn)了DNA-6mA的存在,并且發(fā)現(xiàn)6mA甲基化參與了調(diào)控基因和轉(zhuǎn)座子的表達。此外,已有大量研究表明RNA-m6A修飾在人類mRNA中廣泛存在,與RNA剪接、mRNA穩(wěn)定性和基因表達有關(guān)。但有關(guān)DNA-6mA是否在基因調(diào)控和疾病致病機制中發(fā)揮作用等問題仍未得到深入研究。
大多數(shù)真核生物6mA研究都集中在單細胞原生生物上,包括纖毛蟲四膜蟲、草履蟲以及綠藻。6mA占這些基因組中總腺嘌呤的約0.4%~0.8%,這比哺乳動物和植物中5mC的普遍率低幾倍。有趣的是,也有極少數(shù)的報道表明在最近進化的生物體中存在6mA,包括蚊子、植物,甚至哺乳動物。
3 m6A的生物學(xué)功能
目前表觀遺傳學(xué)在癌癥的發(fā)展中的關(guān)鍵作用越來越受到人們的重視。單靠遺傳學(xué)并不能解釋沒有家族史的個體的散發(fā)性癌癥和癌癥的發(fā)展。表觀遺傳學(xué)為解釋這種情況下癌癥的發(fā)展提供了一種機制。表觀遺傳學(xué)特征的改變可以提供對癌癥的病因和發(fā)生發(fā)展的重要見解。由于幾種表觀遺傳變化發(fā)生在組織病理學(xué)改變之前,它們可以作為癌癥診斷和風(fēng)險評估的生物標志物。根據(jù)表觀基因組信息,已經(jīng)確定了一些可作為診斷工具的表觀遺傳標記物;其中一些生物標記物也可能有助于預(yù)后以及治療效果的判斷[26]。腺嘌呤甲基化對DNA結(jié)構(gòu)的直接影響及其在原核生物學(xué)中的作用已被充分表征[27-28]。6mA是否在真核生物中發(fā)揮保守的功能作用還有待觀察[29-30],但討論其在原核生物中的功能效應(yīng)提出了幾項的潛在功能,需要在真核生物中進一步探索。
3.1 轉(zhuǎn)錄 腺嘌呤的甲基化可通過修飾轉(zhuǎn)錄因子結(jié)合或改變?nèi)旧|(zhì)結(jié)構(gòu)來影響轉(zhuǎn)錄。在細菌中,6mA已經(jīng)被證明可以調(diào)節(jié)轉(zhuǎn)錄[31-32],這提高了真核生物中保留類似功能的可能性。有趣的是,已顯示N6腺嘌呤甲基化對哺乳動物和植物中的轉(zhuǎn)錄因子結(jié)合具有相反的作用。在腺嘌呤的N6位甲基化的轉(zhuǎn)染DNA降低了哺乳動物細胞中轉(zhuǎn)錄因子的DNA結(jié)合親和力[27-28]。 在植物中,鋅指蛋白 AGP1與 DNA 的結(jié)合親和力可以通過靶序列的N6腺嘌呤甲基化來增強[33]。此外,在大麥中使用瞬時表達系統(tǒng),發(fā)現(xiàn)轉(zhuǎn)錄在6mA修飾的報告質(zhì)粒中增加,而5mC對轉(zhuǎn)錄效率幾乎沒有或沒有影響[33]。
在C.reinhardtii中,6mA在TSS周圍富含超過14 000個基因,其中大多數(shù)是活躍轉(zhuǎn)錄的,并且這些6mA位點中的大多數(shù)在幾乎每個分析的細胞中都被甲基化[34]。相反,沉默基因在其TSS周圍具有較低的6mA水平。因此,似乎6mA是活性基因的一般標記,盡管目前還不清楚它是否在基因表達的動態(tài)調(diào)節(jié)中起作用。相比之下,5mC也存在于C.reinhardtii的基因組中,但位于基因體并且與轉(zhuǎn)錄抑制相關(guān)。尚不清楚在線蟲中轉(zhuǎn)錄期間6mA具有什么作用。有趣的是,突變體中6mA水平升高,組蛋白H3賴氨酸4二甲基化(H3K4me2)水平升高,這是與活躍轉(zhuǎn)錄相關(guān)的標志[35]。這表明6mA也可以標記秀麗隱桿線蟲中的活性基因,就像在C.reinhardtii中一樣。
在D.melanogaster中,6mA被提議用于促進轉(zhuǎn)座子表達。6mA-IP-seq測定顯示轉(zhuǎn)座子富集6mA,并且假定的脫甲基酶DMAD的喪失導(dǎo)致轉(zhuǎn)座子表達增加[36]。因此,三個進化上遙遠的生物中的最新發(fā)現(xiàn)表明6mA與基因表達升高的相關(guān)性,盡管仍然有待觀察這種機制的一般性以及6mA是否已進化為在不同物種和/或不同背景中具有不同功能。未來重要的實驗將涉及將DNA甲基轉(zhuǎn)移酶或去甲基化酶引導(dǎo)至特定基因座并分別檢查添加或去除6mA對轉(zhuǎn)錄的影響[37]。
3.2 DNA甲基化和核小體定位 嗜熱鏈球菌的先前研究表明,核小體DNA中的6mA水平低于接頭DNA[38]中的6mA水平。秀麗隱桿線蟲和黑腹果蠅都沒有發(fā)現(xiàn)6mA[38]的核小體定位偏倚。然而,秀麗隱桿線蟲研究是在混合組織上進行的,特定的基因組模式可能只在更詳細的組織特異性檢查中出現(xiàn)。另一方面,在萊茵衣藻中,發(fā)現(xiàn)6mA優(yōu)先位于TSS-近端接頭DNA上。這可以通過有利于接頭DNA的甲基化機制來解釋,可能是由于物理上易于接近,或者通過6mA調(diào)節(jié)核小體定位。有趣的是,某些藻類物種的CG重復(fù)中的甲基化發(fā)生在極端密度下,這不利于核小體組裝,從而決定核小體定位[39]。因此,DNA甲基化可以提供控制單細胞真核生物中核小體定位的一般機制。
3.3 表觀遺傳標記 在原核生物中,6mA甲基轉(zhuǎn)移酶在特定序列基序的情況下甲基化腺嘌呤[40],但真核生物6mA似乎不強烈依賴于序列基序識別。在萊茵衣藻中,6mA發(fā)生在多個序列中,主要位于ApT二核苷酸,類似于哺乳動物中CpG二核苷酸的5mC甲基化[41]。在多個裂變單元循環(huán)期間,穩(wěn)定地保持6mA水平。單堿基圖譜表明大多數(shù)單個6mA位點在變異培養(yǎng)條件下忠實保守,強化了6mA作為可遺傳的表觀遺傳標記[34]的推測。盡管尚未在萊茵衣藻中鑒定出6mA甲基轉(zhuǎn)移酶,但很可能存在從母細胞到子細胞繼承6mA特征的機制。在秀麗隱桿線蟲中,H3K4me2去甲基化酶spr-5的缺失導(dǎo)致跨代的生育力逐漸喪失[42]。這種生育缺陷與H3K4me2的逐漸積累和抑制標記H3K9me365[43]的水平下降同時發(fā)生。在spr-5突變體蠕蟲中,6mA也會在幾代人中增加;6mA去甲基化酶nmad-1的缺失加速,而潛在的6mA甲基轉(zhuǎn)移酶damt-1的缺失抑制了spr-5突變體蠕蟲的進行性生育缺陷。此外,刪除damt-1抑制了H3K4me2的積累,表明N6-腺嘌呤甲基化和H3K4二甲基化可能是共調(diào)節(jié)的并相互增強[44]。雖然6mA或H3K4me2本身是否可以跨代傳輸表觀遺傳信息仍有待確定,但當表觀遺傳信息被不正確地遺傳時,這些修飾會累積。這些發(fā)現(xiàn)提高了6mA可能作為表觀遺傳標記的可能性,其在真核生物中攜帶可遺傳的表觀遺傳信息。
4 展望
在幾十年前的真核基因組中發(fā)現(xiàn)了5mC和6mA都是如此[45]。由于其在哺乳動物和植物中的豐富性,研究集中于表征5mC已經(jīng)揭示了5mC的多種功能,而6mA的研究在真核生物中非常有限。眾所周知,5mC可以自發(fā)脫氨基,導(dǎo)致 C-to-T突變[46];因此,由于甲基化胞嘧啶的脫氨作用,具有5mC的生物體往往會失去CpG二核苷酸[47]。相反,6mA不傾向于自發(fā)突變,這顯然可能有利于基因組穩(wěn)定性。
最近顯示真核mRNA中的腺嘌呤甲基化對基因表達具有深遠的影響[48]。6mA DNA在塑造基因表達中的潛在作用仍然很大程度上未知。RNA N6-甲基腺嘌呤的甲基可以使Watson-Crick堿基配對不穩(wěn)定~1.0 kcal/mol[49-50],這顯示出誘導(dǎo)“m2A-轉(zhuǎn)換”機制,其改變RNA結(jié)構(gòu)并因此通過蛋白質(zhì)結(jié)合[51]。DNA中的相同特性可能影響轉(zhuǎn)錄,復(fù)制和其他需要鏈分離或DNA彎曲的過程。此外,甲基可以促進或抑制DNA結(jié)合蛋白的識別和結(jié)合,以調(diào)節(jié)基因表達。在哺乳動物細胞中,FTO催化的RNA N6-甲基腺嘌呤的去甲基化產(chǎn)生兩種新發(fā)現(xiàn)的中間體,N6-羥甲基腺苷(hm6A)和N6-甲酰腺苷(f6A),在非編碼RNA和mRNA[52]中。由于hm6A和f6A是非常穩(wěn)定的修飾,它們可能不僅僅是中間體,而且具有調(diào)節(jié)功能。還有待觀察是否在DNA 6mA的氧化去甲基化反應(yīng)期間產(chǎn)生類似的修飾,并且如果它們存在,是否它們具有任何生物學(xué)作用。這些修飾可以進一步使DNA雙鏈體失穩(wěn)或產(chǎn)生新的蛋白質(zhì)結(jié)合位點。
值得注意的是,秀麗隱桿線蟲和黑腹果蠅在其基因組DNA中幾乎沒有或沒有5mC,類似于其他一些含 6mA 的生物,如 P.aurelia[51]和 T.thermophila[52]。然而,萊茵哈迪在其基因組中含有6mA和5mC,但5mC的相對總體低水平加上其在外顯子上的不尋常富集表明與最近進化的植物和動物相比,5mC具有不同的功能,其中它主要是標記。6mA和5mC豐度和功能的相對變化可能具有不同的進化后果。正如最近的研究[52]所揭示的,N6-腺嘌呤甲基化可能是影響某些真核生物中基因表達的主要DNA甲基化機制。在其他系統(tǒng)中,它可能在功能上更顯著的5mC DNA甲基化中起補充作用。不同物種中6mA-甲基轉(zhuǎn)移酶同源物的存在提高了6mA可能存在于更多生物體中的可能性,包括哺乳動物。6mA的相對生物學(xué)重要性是否隨著其在最近進化的生物中的較低豐度而下降仍有待觀察。我們推測6mA可能在這些物種中進化出更為特殊的功能。隨著新的,靈敏的檢測技術(shù)的出現(xiàn),目前的研究聚焦于詳細探究這種DNA修飾在整個生命樹中的功能和精確的基因組分布。
猜你喜歡
天天愛科學(xué)(2022年9期)2022-09-15 01:12:54
天天愛科學(xué)(2022年4期)2022-05-23 12:41:48
當代水產(chǎn)(2022年3期)2022-04-26 14:26:56
科學(xué)大眾(2021年9期)2021-07-16 07:02:54
軍事文摘(2020年20期)2020-11-28 11:42:50
航空世界(2020年10期)2020-01-19 14:36:20
小學(xué)科學(xué)(學(xué)生版)(2018年3期)2018-04-18 12:34:19
知識經(jīng)濟·中國直銷(2017年10期)2017-11-07 02:39:52
知識經(jīng)濟·中國直銷(2017年3期)2017-04-16 03:08:01
文學(xué)少年(有聲彩繪)(2017年2期)2017-03-06 08:10:54
