群體感應(yīng)信號分子AI-2高產(chǎn)乳酸菌株篩選及特性研究
2018-02-28 10:05:49蔡針華程娜賈震虎逄曉陽
食品與發(fā)酵工業(yè) 2018年1期
蔡針華,程娜,賈震虎*,逄曉陽
1(山西師范大學 生命科學學院,山西 臨汾,041000) 2(中國農(nóng)業(yè)科學院農(nóng)產(chǎn)品加工研究所,北京,100193)
乳酸菌屬于革蘭氏陽性菌,是一類能利用碳水化合物產(chǎn)生乳酸,并對人體健康有特殊作用的重要益生菌。進入宿主腸道后可以通過生物拮抗作用抑制病原菌的粘附定植[1]和生長繁殖[2-3],在宿主腸道發(fā)育過程中發(fā)揮著重要作用[4]。據(jù)報道乳酸菌具有促進食物消化與吸收、降低血清膽固醇、抑制病原菌和抗感染、抗腫瘤及預(yù)防衰老等作用[5]。迄今為止,對乳酸菌粘附和定植腸道的詳細機制仍在探索之中,但是目前學術(shù)界關(guān)于乳酸菌腸道黏附已經(jīng)達成的初步共識是,腸道菌群和宿主免疫系統(tǒng)通過相互作用減少炎癥反應(yīng),對維持機體微生態(tài)平衡發(fā)揮著重要作用[6]。隨著研究的不斷深入,有關(guān)乳酸菌的研究已涉及生物學和醫(yī)學的各個領(lǐng)域,包括乳酸菌的鑒定和分類、與粘膜免疫機制、益生作用及表達系統(tǒng)等[7]。研究者發(fā)現(xiàn)乳酸菌菌群的群體行為跟很多生物現(xiàn)象相關(guān),比如生物膜形成[8]、自溶[9]、腸道定植能力等,所以大量研究開始關(guān)注乳酸菌的群體行為,而細菌的群體感應(yīng)現(xiàn)象是研究菌群群體行為的突破口[10]。
細菌的群體感應(yīng)(quorum sensing,QS)系統(tǒng)是通過菌體向周圍環(huán)境中分泌群體感應(yīng)信號分子,同時菌體細胞膜上具有感知這種信號分子的受體,當菌群密度達到一定閾值的時候,菌體細胞膜上的信號分子受體感受到這種密度壓力后,通過磷酸化的方式將QS信號傳遞給調(diào)控基因表達的調(diào)控蛋白,通過調(diào)控蛋白對菌體的生理代謝進行調(diào)節(jié)。細菌種間交流主要是通過自體誘導(dǎo)劑-2(Autoinducer-2,AI-2)來完成的[11],AI-2主要是由保守的LuxS蛋白酶在甲基循環(huán)中催化合成,這種信號分子存在于大多數(shù)細菌中,在許多微生物進行種內(nèi)和種間的交流起到了關(guān)鍵的作用。目前已報道在超過80種細菌中存在luxS基因[12-13],不僅在致病菌中有l(wèi)uxS基因,許多益生菌中也擁有l(wèi)uxS基因并能夠生成AI-2[12,14-15]。研究表明,luxS介導(dǎo)的群體感應(yīng)系統(tǒng)對細菌多種生理現(xiàn)象具有重要的調(diào)控作用[16],因此深入研究乳酸菌luxS基因的功能將為后續(xù)解析乳酸菌群體感應(yīng)信號傳導(dǎo)通路、探索乳酸菌在宿主腸道內(nèi)粘附定植的機制奠定基礎(chǔ)。
1 材料與方法
1.1 菌株
8株乳酸菌均為實驗室保藏菌株。大腸埃希氏菌 DH5α 購自北京全式金生物技術(shù)有限公司。哈維弧菌 BB152 (VibrioharveyiBB152),哈維弧菌 BB170 (VibrioharveyiBB170)、均購自美國模式培養(yǎng)物集存庫(American Type Culture Collection,ATCC)。
1.2 培養(yǎng)基及試劑
LB液體培養(yǎng)基:10 g胰蛋白胨(TRYPTONE)(OXOID 公司)、5 g酵母提取物(YEAST EXTRACT)(OXOID公司)、10 g NaCl(生工生物),使溶質(zhì)完全溶解,調(diào)節(jié) pH值至7.5,定容至1 L,121 ℃,15 min滅菌,4 ℃保存?zhèn)溆谩?/p>
AB培養(yǎng)基:1 L培養(yǎng)基中加入0.05 mol/L MgSO4(AMRESCO 公司),0.3 mol/L NaCl(AMRESCO公司),0.2% vitamin-free Casamino Acids(BD公司), 用KOH(AMRESCO公司)調(diào)pH至7.5,121 ℃,15 min滅菌后添加10 mL 1 mol/L K2HPO4(AMRESCO公司)(0.22 μm濾器過濾),10 mL 0.1 mol/LL-精氨酸(AMRESCO 公司)(0.22 μm 濾器過濾),20 mL 50%甘油(0.22 μm濾器過濾)。
MRS液體培養(yǎng)基:北京陸橋技術(shù)股份有限公司;MB液體培養(yǎng)基、MB固體培養(yǎng)基:青島高科園海博生物技術(shù)有限公司;細菌基因組提取試劑盒、細菌RNA提取試劑盒、cDNA反轉(zhuǎn)錄試劑盒、RT-PCR試劑盒,北京全式金生物技術(shù)有限公司。
1.3 主要儀器
凝膠成像儀(GelDocXR+,美國伯樂公司);快速梯度PCR儀(TP600),日本TAKARA公司;實時熒光定量PCR儀(ABI7500),美國Perkin ELmer公司;全波長酶標儀(SPARK20M),瑞士TECAN公司;恒溫培養(yǎng)箱(OJ5-2012R),上海精密儀器有限公司;恒溫金屬浴(MB-102),杭州博日科技有限公司。
1.4 菌種保藏活化及各生長時期的確定
從70多株乳酸菌中經(jīng)過平板計數(shù)挑選8株進行實驗(表1)。

表1 實驗所用菌株
將凍存的8株乳酸菌分別活化3代。每代于37 ℃培養(yǎng)18 h,連續(xù)培養(yǎng)3代即可用于實驗。BB170和BB152凍干粉復(fù)蘇,劃線于MB平皿,28 ℃恒溫箱靜置至生長出大小適宜的單菌落。隨后挑單菌落于MB液體培養(yǎng)基中,每代于28 ℃振蕩培養(yǎng)24 h,連續(xù)活化培養(yǎng)3代即可用于實驗。取商品化DH5α,于LB
液體培養(yǎng)基,37 ℃,200 r/min振蕩培養(yǎng)10 h至指數(shù)期,每代于37 ℃振蕩培養(yǎng)10 h,連續(xù)培養(yǎng)3代即可用于實驗,每次使用按4%的接種量接種于新鮮MRS培養(yǎng)基中。取活化好的乳酸菌菌液以4%的接種量于新鮮的MRS液體培養(yǎng)基中混勻,取1 mL于24孔透明酶標板中,用多功能酶標儀每隔1 h測1次OD600 nm值,繪制生長曲線確定各菌株的指數(shù)期,平臺期。
1.5 生物學方法檢測信號分子AI-2
將已活化好的8株乳酸菌按4%接種至滅菌后的MRS液體培養(yǎng)基中,37 ℃繼續(xù)培養(yǎng)18 h(平臺期)大約109CFU/mL,將菌液6 000 r/min離心10 min,上清液用0.22 μm的滅菌濾菌器過濾分裝至2 mL離心管中,得到各乳酸菌的無菌上清液;按相同方法收集28 ℃振蕩培養(yǎng)13 h的V.harveyiBB152無菌上清液和37 ℃培養(yǎng)的DH5α無菌上清液分別作為陽性對照和陰性對照;所得無菌上清液分別貯藏于-80 ℃冰箱備用。
將報告菌株培養(yǎng)至菌體OD600 nm為0.8~1.2,然后用新鮮的AB培養(yǎng)基以1∶5 000的比例與報告菌株混勻。分別將各乳酸菌、V.harveyiBB152、DH5α無菌上清液作為待測樣品、陽性對照和陰性對照,經(jīng)條件優(yōu)化后按1∶100混合于稀釋后的報告菌株培養(yǎng)液中,置于30 ℃繼續(xù)振蕩培養(yǎng)。不加任何菌液的只有稀釋過的報告菌株培養(yǎng)液作為介質(zhì)對照,1~6 h內(nèi),每隔1 h取200 μL至96孔黑色酶標板中,用多功能酶標儀的生物發(fā)光模式檢測其熒光強度值,在1~6 h內(nèi),以陰性對照組熒光強度值降到最小的時間點為基準,此時陽性組相對熒光強度設(shè)為1,信號分子AI-2的強度用相對熒光表示。樣品與對照均做3個復(fù)孔,每次實驗重復(fù)3次。按照以下方式計算各菌株熒光強度:陰性對照組熒光強度為陰性對照的熒光強度值與陽性對照的熒光強度值之比;待測組相對熒光強度值為待測樣品的熒光值與介質(zhì)對照熒光值之比。
1.6 luxS基因的生物學鑒定
1.6.1 引物的設(shè)計
根據(jù) NCBI上luxS基因的DNA全長序列(No.1063754),應(yīng)用Oligo7設(shè)計內(nèi)參引物(16S rRNA-F和16S rRNA-R) 及特異性引物 (LuxS-F和LuxS-R)、(LDH-F和LDH-R)并由北京六合華大基因科技股份有限公司合成,具體引物序列見表2。
1.6.2 菌種RNA提取及cDNA合成
將4株乳酸菌按4%的接種量轉(zhuǎn)接入MRS液體培養(yǎng)基中,37 ℃培養(yǎng)18 h至平臺期,離心收集菌體細胞,按照常規(guī)Trizol法提取菌體總RNA。cDNA按如下體系進行合成:3μL RNA(1 μg)溶液中加入 4 μL Mix和 1 μL gDNA Remove,Rnase-free Water至20 μL,輕輕混勻后 42 ℃孵育15 min,85 ℃加熱5 s以獲得cDNA。
1.6.3 實時熒光定量PCR
PCR反應(yīng)體系(20 μL)為:Tip Green Qpcr SuperMix 10 μL,ddH2O 8.5 μL,上、下游引物(10 μmol/L)各0.4 μL,Passive Reference Dye(50×) 0.4 μL,模板1 μL。PCR 反應(yīng)條件為 95 ℃ 10 min;95 ℃ 5 s,60 ℃ 15 s,72 ℃ 34s,40個循環(huán);PCR產(chǎn)物經(jīng)1%瓊脂糖凝膠電泳檢測,將產(chǎn)物送至北京六合華大基因科技股份有限公司進行DNA測序。
1.7 乳酸菌種內(nèi)不同pH下信號分子AI-2與菌體密度的關(guān)系
將篩選出高產(chǎn)AI-2信號的乳酸菌AST18進行耐酸性與信號分子AI-2的研究,備好的菌液按4%接種到不同pH培養(yǎng)基中,37 ℃培養(yǎng)18 h,按1.3.2的方法檢測不同pH培養(yǎng)條件下乳酸菌AI-2的情況。樣品與對照均做3個復(fù)孔,每次實驗重復(fù)3次。
2 結(jié)果與討論
2.1 各菌株生長周期的確定
結(jié)合表1和圖1選取實驗室的8株乳酸菌是因為他們在連續(xù)活化3代培養(yǎng)18 h后,經(jīng)平板計數(shù)菌落數(shù)都是109CFU/mL,通過多功能酶標儀獲得各乳酸菌的生長曲線,它們有相同的生長周期,在培養(yǎng)18 h后都達到了對數(shù)末期,保證了幾株菌的菌群密度一致性,幾株菌又是實驗室研究較多的,遺傳背景比較清楚,故實驗選取這8株乳酸菌來進行后續(xù)實驗。相關(guān)研究已經(jīng)證實,乳酸菌產(chǎn)信號分子AI-2通常都在對數(shù)末期或者穩(wěn)定初期達到最高[17-18],這可能是由于大量 AI-2信號分子隨著菌體密度的增高而增多,但當其濃度達到一定閾值時,發(fā)生的內(nèi)化作用,菌體重吸收或附著在細胞膜表面,調(diào)控穩(wěn)定期的某些重要生理功能[18]。由圖1顯示,8株乳酸菌大概從3 h進入指數(shù)期,10 h到達指數(shù)末期,之后進入平臺期,OD600 nm在指數(shù)末期達到最大,在18 h時OD600 nm趨于穩(wěn)定,處于平臺期,因此實驗中選擇培養(yǎng)18 h時測定不同乳酸菌產(chǎn)信號分子 AI-2的熒光強度。
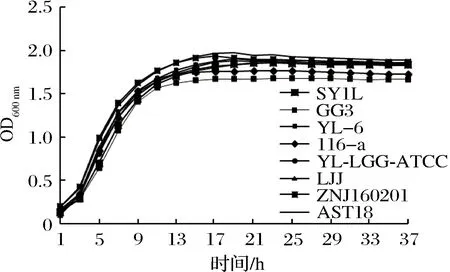
圖1 8株乳酸菌的生長曲線Fig.1 Growth curves of eight strains of lactic acid bacteria
2.2 菌株培養(yǎng)上清AI-2信號檢測
結(jié)合已研究的方法[17-20]進行優(yōu)化設(shè)計,實驗中用到的指示菌株V.harveyiBB170是標準菌株V.harveyiBB120的定向突變菌株,產(chǎn)信號分子AI-2的luxS基因是完整的,所以選它為報告菌株。不同細菌產(chǎn)生的信號分子AI-2,都能被報告菌株V.harveyBB170識別,誘導(dǎo)其熒光酶基因的表達,從而產(chǎn)生發(fā)光反應(yīng)[21]。V.harveyBB152也是突變體且只能產(chǎn)生信號分子AI-2,故作為陽性對照[17],大腸埃氏菌DH5α由于缺失luxS基因不能產(chǎn)生AI-2信號,故作為陰性對照。由圖2可知,在0~3 h內(nèi)所有對照組的熒光強度均隨時間的延長而在不斷下降,且陰性對照在3 h時熒光強度已經(jīng)達到最低,之后開始升高,由于陰性對照中只包含稀釋后的報告菌株,表明當培養(yǎng)時間達到3 h時,指示菌產(chǎn)生的信號分子AI-2濃度達到誘導(dǎo)其發(fā)光的閾值。因此,以3 h時的熒光強度值為標準計算各菌株相對熒光強度,表示信號分子AI-2活性。依據(jù)文獻[22]研究發(fā)現(xiàn),V.harveyiBB170構(gòu)成的AI-2檢測體系對pH值較敏感,過酸或者過堿均會對指示菌的發(fā)光性產(chǎn)生影響。從最優(yōu)角度出發(fā),按照1∶100的加樣比例添加,此時體系中既含有足夠量的AI-2信號分子足以誘導(dǎo)V.harveyiBB170產(chǎn)生熒光,同時又避免了培養(yǎng)上清液中酸度和抑菌物質(zhì)對指示菌的影響[19]。檢測結(jié)果如圖3所示,8株乳酸菌的相對熒光強度只有1株SY13小于陰性對照,說明其中7株乳酸菌均可以產(chǎn)信號分子AI-2,其中乳酸菌YL-6、AST18和ZNJ160201的相對熒光強度明顯高于其他幾株,乳酸菌LGG、YL-LGG-ATCC和LJJ的相對熒光強度較弱。
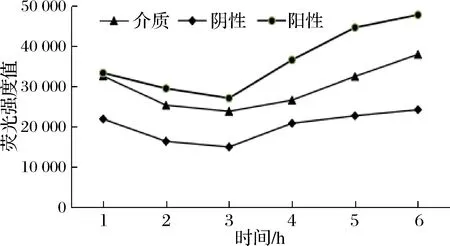
圖2 不同孵育時間各樣品的熒光強度值Fig.2 The luminescence of different incubation time of samples
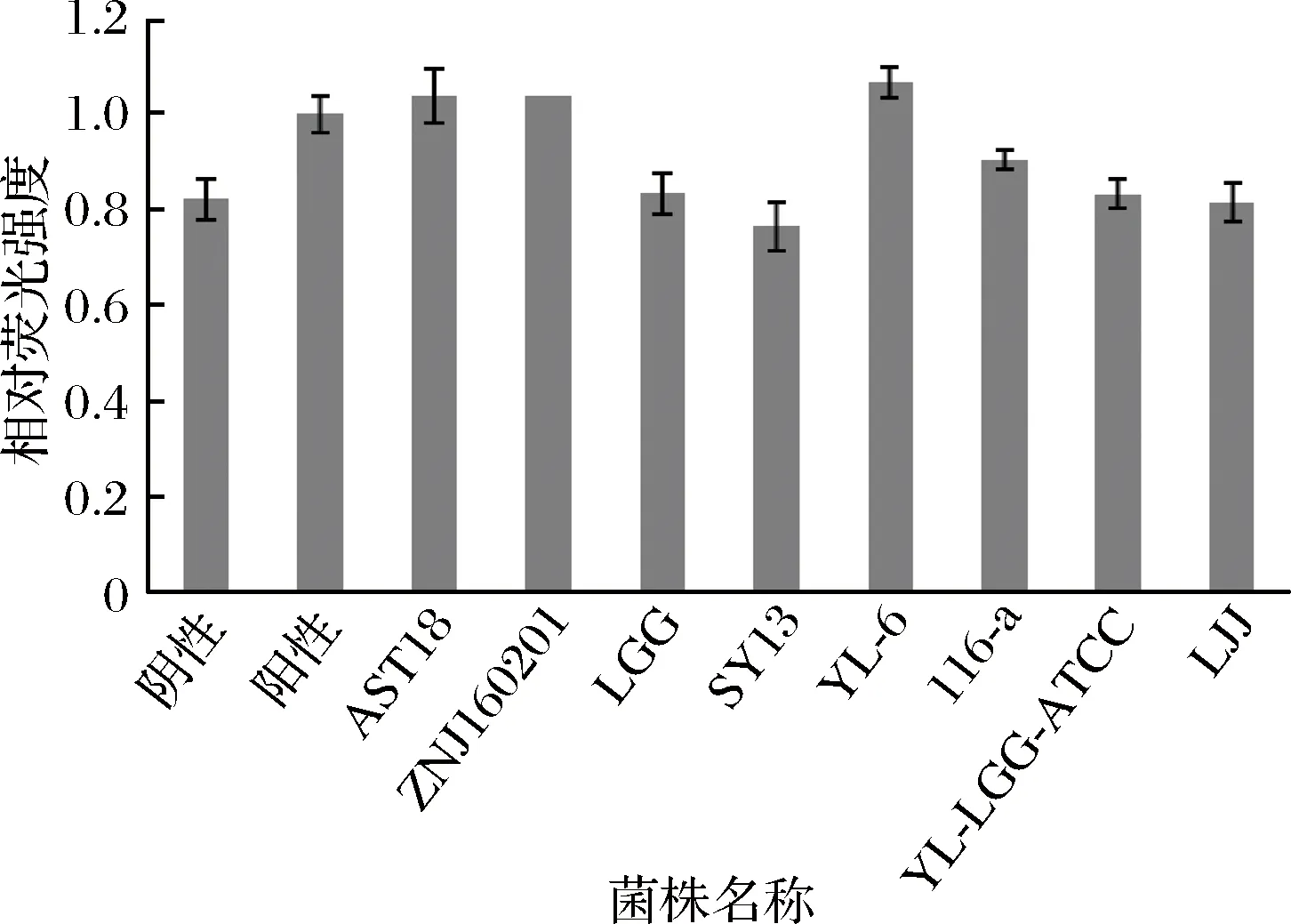
圖3 各乳酸菌培養(yǎng)18 h上清液的相對熒光強度Fig.3 The relative luminescence of lactic acid bacteria in incubation time 18 h
2.3 RT-PCR檢測luxS基因表達情況
已被證明[23]在細菌物種中導(dǎo)致產(chǎn)生AI-2的生物合成途徑是高度保守的,且已經(jīng)在不同乳桿菌的基因組中發(fā)現(xiàn)了LuxS同源物,同時早些時候已經(jīng)研究了一些乳桿菌屬物種產(chǎn)生AI-2的情況[22,24-25]。參與AI-2生物合成的重要酶是S-核糖基半胱氨酸酶,也稱為LuxS蛋白,基因組分析發(fā)現(xiàn)55種以上的細菌均含有AI-2合成酶luxS的同源保守序列[26]。選取其中乳酸菌AST18、ZNJ160201和SY13、YL-LGG-ATCC這4株菌用生物學方法來檢測AI-2熒光強度是否是由關(guān)鍵基因luxS所控制和熒光方法的準確性。對4株乳酸菌進行RT-PCR檢測,luxS基因的表達情況如圖4。luxS基因在乳酸菌AST18、ZNJ160201表達較強,在YL-LGG-ATCC表達較弱,而在SY13菌株中幾乎沒有表達,與圖2的熒光結(jié)果較一致,同時通過結(jié)合兩種檢測結(jié)果,說明AI-2熒光強度是由關(guān)鍵基因luxS所控制。
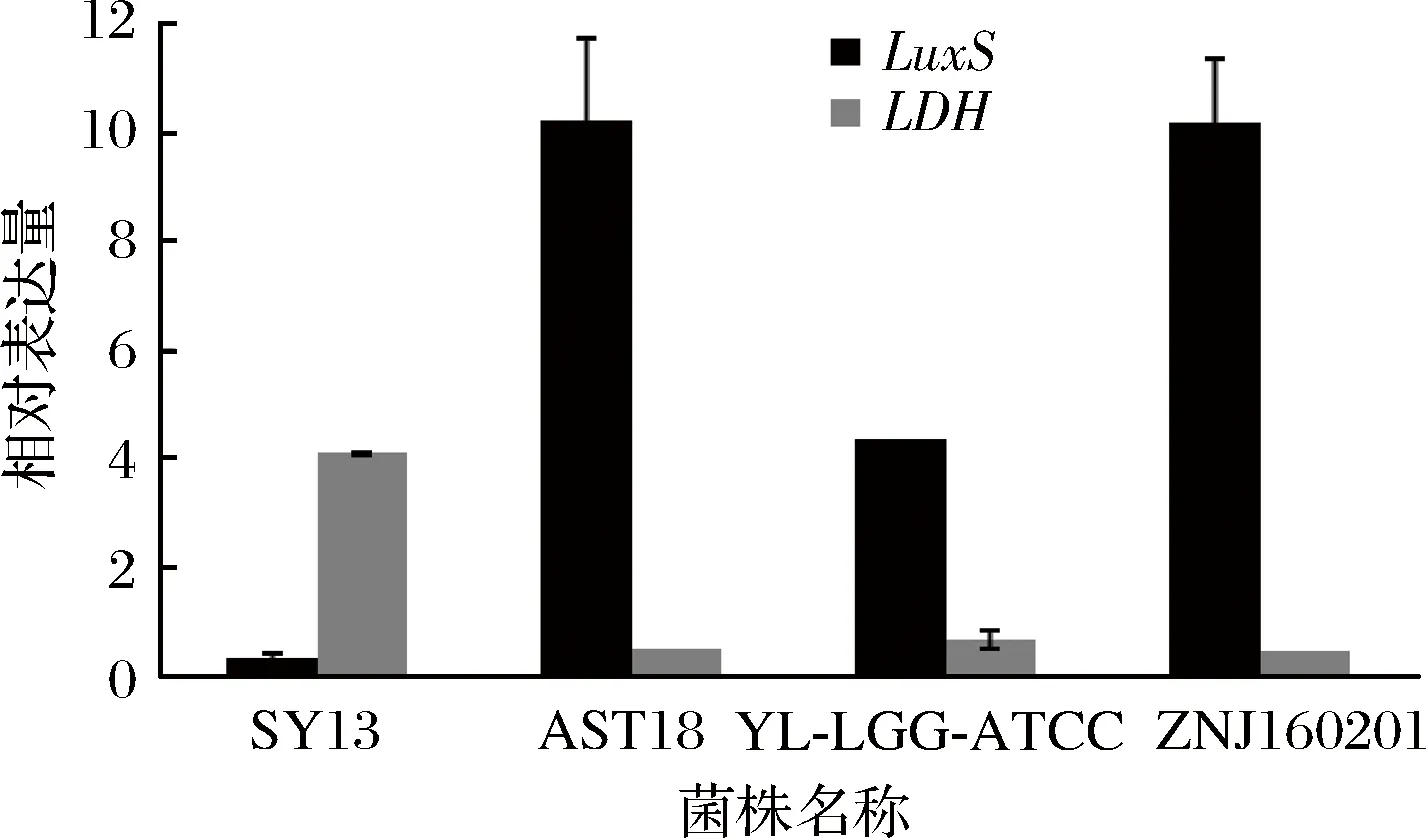
圖4 四株乳酸菌luxS基因和LDH基因的相對表達量Fig.4 Relative expression of luxS gene and LDH gene of four lactic acid bacteria
LEBEE[24]等人已經(jīng)研究得出luxS基因在乳酸菌中有中樞代謝作用。同時群體感應(yīng)必不可少可能伴隨著能量反應(yīng),luxS基因可能間接的參與了EMP,磷酸戊糖和TCA等循環(huán),因而實驗又選擇LDH基因來檢測QS系統(tǒng)與新陳代謝之間的關(guān)系。發(fā)現(xiàn)當luxS基因表達較強時,而LDH基因表達較弱,LDH基因編碼乳酸脫氫酶的生成,進而催化丙酮酸生成乳酸,乳酸是谷氨酸發(fā)酵的主要副產(chǎn)物,所以當luxS基因表達較強時,LDH基因相對較弱,乳酸生成相對較少,有利于宿主體內(nèi)腸道的酸堿平衡,繼而維持腸道菌群平衡,從而在腸道內(nèi)發(fā)揮益生作用。對這4株菌進行基因組提取,普通PCR擴增后,經(jīng)過聚丙烯酰胺凝膠電泳結(jié)果如圖5所示,與實時熒光聚合酶列式反應(yīng)結(jié)果一致,說明菌株AST18和ZNJ160201的AI-2信號強,luxS基因表達較強,AI-2信號強度是由關(guān)鍵基因luxS所主導(dǎo)。同時通過3個結(jié)果的一致性也說明了熒光檢測與RT-PCR檢測相結(jié)合是比較準確的方法用于群體感應(yīng)信號分子AI-2的檢測。
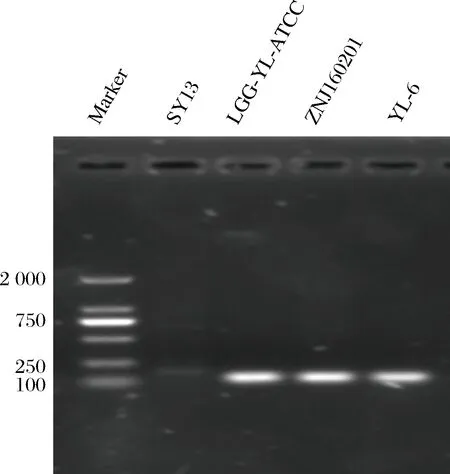
圖5 LuxS基因PCR產(chǎn)物電泳圖Fig.5 Electrophoresis of luxS gene amplified by PCR
2.4 生物發(fā)光與耐酸性特性研究
據(jù)報道[27],有機酸、蘋果酸對致病菌AI-2信號的產(chǎn)生有抑制作用。luxS基因參與環(huán)境調(diào)控脅迫,如酸性脅迫已經(jīng)在變形鏈球菌中被證明[28-29],但是該基因在乳酸桿菌中酸適應(yīng)的作用尚不清楚。實驗探究乳酸菌在強酸環(huán)境中生長后AI-2信號變化情況,結(jié)果如圖6,乳酸菌在不同pH的培養(yǎng)基中培養(yǎng)后,AI-2信號也大不相同。在菌體數(shù)達到一定閾值時,能夠激發(fā)報告菌株發(fā)光酶的活性,誘導(dǎo)報告菌株發(fā)光,酸性越強AI-2信號越強。當pH等于3.5時,菌體密度幾乎是最低的,但是AI-2信號是最強的。而當pH等于2.5時,過酸的環(huán)境不利于菌體的生長,可能是菌體數(shù)沒有達到閾值,所以AI-2信號較弱,上述所得結(jié)果與1.3中檢測的LDH基因和luxS基因的表達相符合。這樣的結(jié)果說明AI-2信號提高了乳酸菌酸脅迫作用,繼而發(fā)揮其腸道益生作用;同時也說明菌體密度并不是群體感應(yīng)變化的主要原因,可能與其所處的環(huán)境有極大的關(guān)系。為以后研究乳酸菌通過群體感應(yīng)系統(tǒng)調(diào)節(jié)腸道菌群平衡及發(fā)揮免疫功能提供了一定的幫助。
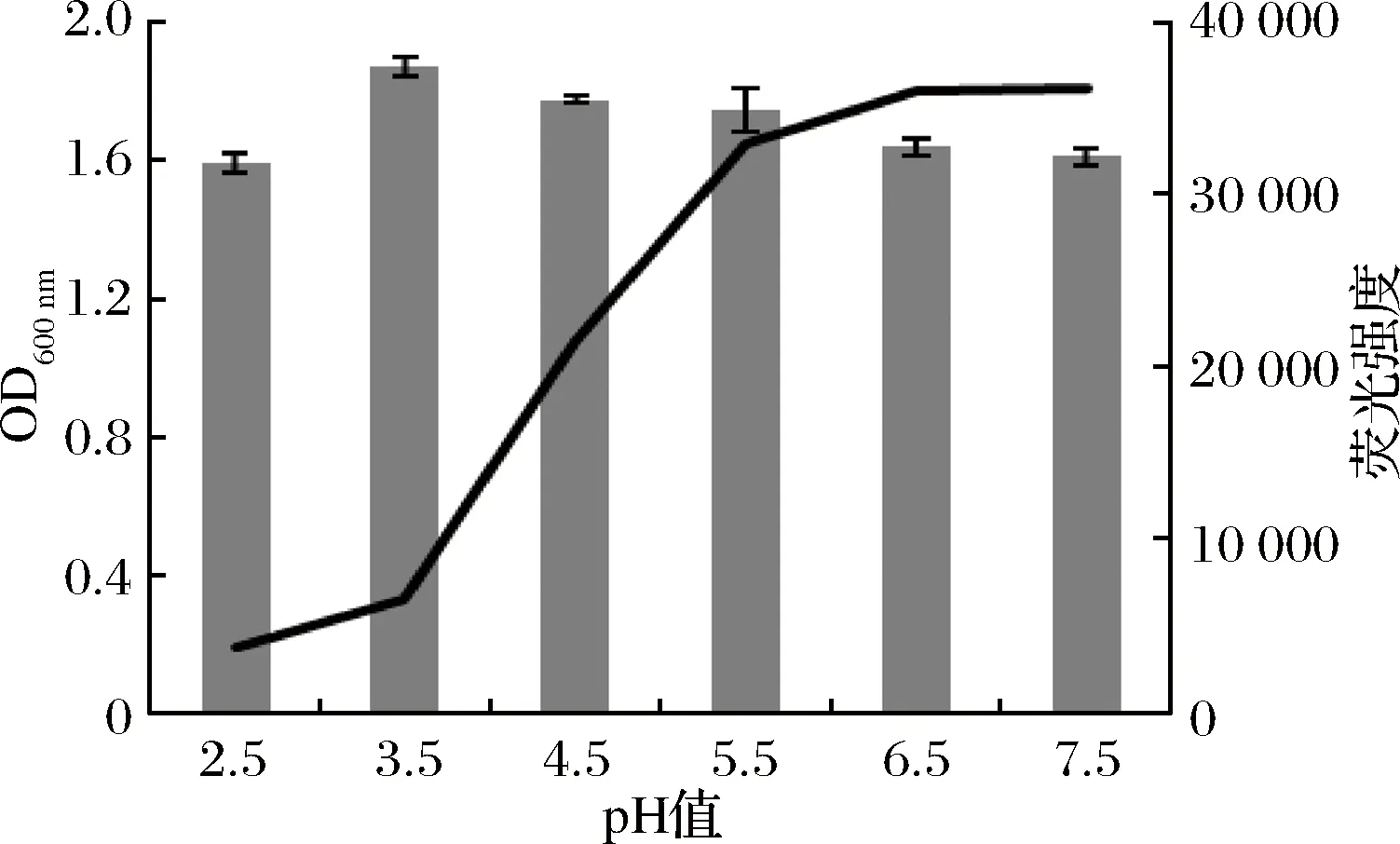
圖6 不同pH條件下熒光強度與OD600 nm的關(guān)系Fig.6 The relationship of light intensity and OD600 nm under unequal pH condition
3 結(jié)論
本研究篩選出3株高產(chǎn)群體感應(yīng)信號分子AI-2乳酸菌,通過RT-PCR檢測證明了熒光篩選方法的可靠性,同時確定了luxS基因編碼的LuxS蛋白是控制群體感應(yīng)信號分子AI-2的主要酶。同時得出在酸性環(huán)境中AI-2信號更強,說明乳酸菌能夠通過群體系統(tǒng)的調(diào)控在酸性腸道內(nèi)存活,并調(diào)控腸道菌群平衡,發(fā)揮其免疫功能。本研究為下一步研究luxS突變株與群體感應(yīng),免疫調(diào)節(jié)功能等關(guān)系奠定了基礎(chǔ)。
[1] BAUMGART D C ,SDIGNASS A U. Intestinal barrier function[J]. Curr Opin Clin Nutr Metab Care, 2002, 5(6): 685-694.
[2] LI X J,YUE L Y,GUAN X F,et al. The adhesion of putative probiotic lactobacilli to cultured epithelial cells and porcine intestinal mucus[J]. J Appl Microbiol, 2008, 104(4): 1 082-1 091.
[3] OHLAND C L,MACNAUGHTON W K. Probiotic bacteria and intestinal epithelial barrier function[J]. Am J Physiol Gastrointest Liver Physiol, 2010, 298(6): G807-819.
[4] SOMMER F, BACKHED F.The gut microbiota-masters of host development and physiology[J]. Nat Rev Microbiol, 2013, 11(4): 227-238.
[5] 代永剛,田志剛,南喜平. 乳酸菌及其生理功能研究的進展[J]. 農(nóng)產(chǎn)品加工(學刊), 2009(7): 24-26;29.
[6] 李紅娟.LactobacilluscaseiAST18抗真菌代謝產(chǎn)物分析及抑菌作用研究[D].北京:中國農(nóng)業(yè)科學院, 2014.
[7] 任大勇,李昌,秦艷青,等. 乳酸菌益生功能及作用機制研究進展[J]. 中國獸藥雜志,2011,45(2):47-50.
[8] 謝麗斯,張宏梅,劉學祿,等. 從腐敗食品中分離的乳酸菌生物被膜形成的影響因素[J]. 食品與發(fā)酵工業(yè), 2011, 37(3): 6-8.
[9] 孫潔. 乳酸菌發(fā)酵劑菌株的自溶特性及機理研究[D].北京:中國農(nóng)業(yè)科學院, 2010.
[10] 馮鎮(zhèn),張?zhí)m威. 乳酸桿菌LB-3自溶酶特性研究[J]. 中國乳品工業(yè), 2008, (9): 18-21.
[11] ASCENSO O S,MARQUES J C,SANTOS A R,et al. An efficient synthesis of the precursor of AI-2, the signalling molecule for inter-species quorum sensing[J]. Bioorg Med Chem, 2011, 19(3): 1 236-1 241.
[12] PARK H,YEO S,JI Y,et al.Autoinducer-2 associated inhibition byLactobacillussakeiNR28 reduces virulence of enterohaemorrhagicEscherichiacoliO157:H7[J]. Food Control, 2014, 45: 62-69.
[13] JIMENEZ J C,EDERLE M J. Quorum sensing in group A Streptococcus[J]. Frontiers in Cellular and Infection Microbiology, 2014, 4(4):127.
[14] CHRISTIAEN S E,CONNELL MOTHERWAY M O,BOTTACINI F,et al. Autoinducer-2 plays a crucial role in gut colonization and probiotic functionality ofBifidobacteriumbreveUCC2003[J]. PLoS One, 2014, 9(5): e98111.
[15] YEO S,PARK H, JI Y,et al. Influence of gastrointestinal stress on autoinducer-2 activity of twoLactobacillusspecies[J]. FEMS Microbiol Ecol, 2015, 91(7).DOI:10.1093/femsec/fivo65.
[16] MAN Li-li,MENG Xiang-chen,ZHAO Ri-hong. Induction of plantaricin MG under co-culture with certain lactic acid bacterial strains and identification ofLuxSmediated quorum sensing system inLactobacillusplantarumKLDS1.0391[J]. Food Control, 2012, 23(2): 462-469
[17] MOSLEHI-JENABIAN S,GORI K,JESPERSEN L.AI-2 signalling is induced by acidic shock in probiotic strains ofLactobacillusspp.[J]. International Journal of Food Microbiology, 2009, 135(3): 295-302.
[18] GORI K,MOSLEHI-JENABIAN S,PURROTTI M,et al. Autoinducer-2 activity produced by bacteria found in smear of surface ripened cheeses[J]. International Dairy Journal, 2011, 21(1): 48-53.
[19] 張騰,賀銀鳳. 堅強腸球菌SQ-3-2基于Lux S群體感應(yīng)系統(tǒng)信號分子AI-2的檢測及方法優(yōu)化[J]. 食品與發(fā)酵工業(yè), 2013, 39(2): 168-173.
[20] LI Z,NAIR S K.Quorum sensing: How bacteria can coordinate activity and synchronize their response to external signals?[J]. Protein Science, 2012, 21(10): 1 403-1 417.
[21] LEBEER S, CLAES I J J, VERHOEVEN T L A,et al. Impact ofluxSand suppressor mutations on the gastrointestinal transit ofLactobacillusrhamnosusGG[J]. Applied and Environmental Microbiology, 2008, 74(15): 4 711-4 718.
[22] DEKEERSMAECKER S C,VANDERLEYDEN J.Constraints on detection of autoinducer-2 (AI-2) signalling molecules usingVibrioharveyias a reporter[J]. Microbiology, 2003, 149(8): 1 953-1 956.
[23] WINANS S C,BASSLER B L. Mob psychology[J]. J Bacteriol, 2002, 184(4): 873-883.
[24] LEBEER S,DE KEERSMAECKER S C,VERHOEVEN T L. Functional analysis ofluxSin the probiotic strainLactobacillusrhamnosusGG reveals a central metabolic role important for growth and biofilm formation[J]. J Bacteriol, 2007, 189(3): 860-871.
[25] TANNOCK G W,GHAZALLY S,WALTER J,et al. Ecological behavior ofLactobacillusreuteri100-23 is affected by mutation of theluxSgene[J]. Applied and Environmental Microbiology, 2005, 71(12): 8 419-8 425.
[26] RAO R M,PASHA S,SOWDHAMINI R. Genome-wide survey and phylogeny of S-Ribosylhomocysteinase (LuxS) enzyme in bacterial genomes[J]. BMC Genomics, 2016, 17(1): 742.
[27] ALMASOUD A, HETTIARACHCHY N,RAYAPROLU S,et al. Inhibitory effects of lactic and malic organic acids on autoinducer type 2 (AI-2) quorum sensing ofEscherichiacoliO157:H7 andSalmonellaTyphimurium[J]. Lwt-Food Science and Technology, 2016, 66: 560-564.
[28] WEN Z T, BUME R A.LuxS-mediated signaling inStreptococcusmutans is involved in regulation of acid and oxidative stress tolerance and biofilm formation[J]. J Bacteriol, 2004, 186(9): 2 682-2 691.
[29] SZTAJER H,LEMME A,VILCHEZ R,et al. Autoinducer-2-regulated genes inStreptococcusmutansUA159 and global metabolic effect of theluxSmutation[J]. J Bacteriol, 2008, 190(1): 401-415.
猜你喜歡
中學生數(shù)理化·七年級數(shù)學人教版(2021年6期)2021-11-22 07:50:58
中學生數(shù)理化·七年級數(shù)學人教版(2021年6期)2021-11-22 07:50:58
中學生數(shù)理化·七年級數(shù)學人教版(2021年6期)2021-11-22 07:50:58
鴨綠江(2021年35期)2021-04-19 12:24:18
中學生數(shù)理化·七年級數(shù)學人教版(2020年12期)2021-01-18 06:57:46
中學生數(shù)理化·七年級數(shù)學人教版(2020年12期)2021-01-18 06:57:46
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
中國生殖健康(2019年3期)2019-02-01 06:12:26
海峽科技與產(chǎn)業(yè)(2016年3期)2016-05-17 04:32:12
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
