豚鹿幾種激素的季節性變化模式研究
2018-01-30 01:58:02嚴慧娟王巍易軍牛李麗屈羽陳昂蒲陽鄧家波鐘燕余建秋
四川動物 2018年1期
嚴慧娟, 王巍, 易軍, 牛李麗, 屈羽, 陳昂, 蒲陽, 鄧家波, 鐘燕, 余建秋*
(1.成都動物園,成都野生動物研究所,成都610081;2. 四川省畜牧科學研究院,動物遺傳育種四川省重點實驗室,成都610066)
豚鹿Axisporcinus隸屬于哺乳綱Mammalia偶蹄目Artiodactyla鹿科Cervidae斑鹿屬Axis,中國僅有印支亞種A.porcinusannamiticus,主要分布于云南西部靠近中緬邊境的耿馬、西盟兩縣的南丁河沿岸,1975年被《瀕危野生動植物種國際貿易公約(CITES)》列入附錄Ⅰ。近20年來,全球野生豚鹿數量急劇減少,世界自然保護聯盟(IUCN)在2008年將其收錄到受威脅物種紅色名錄并列為瀕危(EN)等級。在中國,豚鹿被列為國家Ⅰ級重點保護野生動物,是我國數量最稀少的鹿類之一。目前,我國豚鹿野生種群基本滅絕(汪松,1998),還保存有國家動物園系統的人工飼養群體,數量僅40只左右,已成為瀕危小種群。豚鹿的保種及擴繁一直受基礎群體數量有限、繁殖活動規律性不強和近親交配等因素制約。通過調查成都動物園豚鹿群體繁殖成活率,發現豚鹿的初生死亡率高達60%,主要因為其繁殖行為規律不明確,無法進行相應的機體調節和環境控制措施,導致初生幼崽發生環境應激性死亡(鄧家波等,2011)。因此,明確各季節豚鹿體內繁殖相關激素變化模式,確定其繁殖行為規律,對于提高繁殖成活率至關重要。
動物的繁殖行為涉及下丘腦-垂體-性腺軸系統復雜的神經內分泌過程(何世山,金小軍,2003)。光周期循環和溫度等一系列季節性節律變化因素能夠轉換為生理信號,刺激kisspeptin神經元(Silveiraetal.,2010),作用于下丘腦的GnRH神經元,改變GnRH和促性腺激素的分泌模式,調節生殖行為(Malpauxetal.,2002;Smithetal.,2008)。對反芻動物的研究發現,直接參與繁殖的激素主要包括促黃體素(LH)、外周血液促卵泡素(FSH)和催乳素(PRL),間接參與繁殖行為的激素為三碘甲腺原氨酸(T3)、四碘甲腺原氨酸(T4)(賴平等,2012;狄冉等,2015)。本研究通過監測2015年5月、8月和10月雌性豚鹿血液中繁殖相關激素含量的變化,結合溫度、濕度和光照的變化,粗略繪制春季、夏季和秋季豚鹿季節性內分泌調節趨勢圖。揭示圈養條件下季節變化與豚鹿繁殖的關系,明確豚鹿繁殖活動規律,以便根據季節調整養殖模式,有效提高豚鹿繁殖成活率,逐步擴繁種群。
1 材料與方法
1.1 試驗群體組建
在成都動物園的豚鹿群體中選擇16月齡左右的8頭雌鹿作為實驗動物,固定實驗群體。
1.2 溫濕指數及光照時間測定
在鹿舍中部距地面1.5 m處掛置干濕球溫度計,于2015年5—12月,記錄每日09∶00、12∶00、20∶00的溫度和濕度。按照Sandy(2000)提出的溫濕度指數(temperature-humidity index,THI):THI=0.8×溫度+相對濕度×(溫度-14.4)+46.4計算;根據國際上THI值劃分熱應激標準:THI<72,無熱應激;THI=72~79,溫和應激;THI=80~89,中等應激;THI=90~98,嚴重應激;THI>98,致死應激。同時記錄每天晝長時間。
1.3 血液樣本采集和生化指標測定
實驗期間,當連續3 d的THI值差異無統計學意義(P>0.05)時,于12∶00—14∶00采集8頭豚鹿的外周靜脈血,3 000 r·min-1離心10 min將血清和紅細胞迅速分離,分裝,于-20 ℃凍存。用T3、T4、LH、FSH和PRL的R & D上海卡麥舒ELISA試劑盒檢測。
取微孔平板,加入樣本10 μL,添加稀釋液40 μL,再加入HRP標記抗體100 μL,于37 ℃封閉水浴60 min。反復洗滌5次,加入底物各50 μL于37 ℃避光孵育15 min,加入50 μL終止液,于15 min內在450 nm波長處測定光密度值。
1.4 數據分析
實驗數據采用SPSS 17.0進行K-S檢驗,顯著性水平為α=0.05。利用R語言對差異性狀及群體進行層次聚類分析。
2 實驗結果
2.1 豚鹿圈舍THI值和光照趨勢
8月豚鹿圈舍內THI值為72~80,為溫和應激。其余月份的THI值均在72以下,無熱應激。晝長時間變化趨勢分析發現,6—8月的光照時間較長,日均達到13.63 h以上。10—12月的光照時間較短,日均為10.66 h(圖1)。
2.2 豚鹿激素水平季節性變化趨勢
分析發現,8月豚鹿血清中T3、T4的含量極顯著低于5月、10月(P<0.01)。LH、PRL的含量在8月均極顯著下降(P<0.01),在10月極顯著回升(P<0.01)。FSH在8月的含量極顯著低于5月(P<0.01),但與10月的差異無統計學意義(P>0.05)(圖2)。
2.3 各激素指標間相關性分析
血清中LH、FSH、PRL、T3、T4含量相關性分析顯示(表1),FSH與T3呈極顯著正相關(P<0.01,r=0.647),PRL與T4呈顯著正相關(0.01 圖1 豚鹿圈舍溫濕度指數和晝長時間 圖2 豚鹿血清中激素含量變化趨勢Fig. 2 The variation of hormone content in serum of Axis porcinus **P<0.01,*P<0.05; 下同the same below 表1 豚鹿血清中激素含量相關性分析(n=24)Table 1 Correlation analysis of hormone content in serum of Axis porcinus (n=24) 將5月、8月和10月各樣本中LH、FSH、PRL、T3、T4活性情況進行層次聚類分析,結果發現,T3、T4聚為一類,LH、PRL聚為一類,并且與FSH為二級聚類(圖3)。表明T3、T4活性的變化模式相似度較高,LH、PRL活性的變化模式相似度較高,但FSH的活性變化模式與其余4種激素有差異。 圖3 豚鹿血清各激素聚類圖Fig. 3 Cluster of hormones in serum of Axis porcinus T3、T4在血清中的含量變化與熱應激的時間和應激程度存在密切聯系(Yokusetal.,2006)。研究發現,T3、T4在各樣本中的變化模式相同;8月圈舍THI值達72~80、日照時間達13.63 h以上時,豚鹿血清中T3、T4的含量極顯著低于5月、10月,與同為反芻動物的奶牛在濕熱脅迫下的表現一致(Collieretal.,1981;魏學良等,2005;樊華等,2007)。對馬鹿Cervuselaphus的研究亦發現,T3、T4的分泌存在季節性變化(施振旦,1995)。推測較高的溫濕度和長日照所形成的脅迫造成了機體應激,使血漿中的T3、T4含量均顯著降低,從而調節自身體溫。大量研究表明,T3、T4含量的改變并不引起褐黑激素的變化,而通過影響下丘腦(Websteretal.,1991),調節kisspeptin神經系統和GnIH神經系統對動物的季節性繁殖起調節作用(Bernal,2002;Yasuoetal.,2006)。本研究通過激素間的相關性分析,證明了T3、T4的活性與繁殖相關激素具有相關關系。因此,推測高溫、高濕和長日照的脅迫可能引起血清中T3、T4含量的變化,間接影響豚鹿的繁殖表現。 LH是由垂體前葉促黃體素細胞嗜堿性細胞產生的一種糖蛋白激素,能夠作用于成熟的卵泡并促發排卵(Wuetal.,2000)。研究發現,在繁殖期不同階段,丘腦GnRH分泌的脈沖頻率不同。繁殖期開始時,LH在排卵期前達到最大值,在繁殖末期降至最低,隨后卵巢進入靜止期(Clarke,1993;王訓立等,2002)。FSH是由垂體前葉嗜堿性細胞分泌的一種糖蛋白激素,能夠在繁殖期促使卵泡內膜細胞分泌雌激素,激發卵泡的最后成熟,誘發排卵并使顆粒細胞變成黃體細胞(解美娜等,2004)。對灘羊Procapraprzewalskii的研究發現,FSH和LH含量變化具有顯著的季節性,且秋冬高于春夏(祁云霞等,2014)。另有研究發現,隨春季光照延長,哺乳動物達到FSH分泌上升的光照閾值,隨后LH也隨著光照的進一步延長而增加(楊海明,2014)。PRL作為一種由垂體前葉腺嗜酸細胞分泌的蛋白質激素,可以促進乳腺生長發育,刺激并維持泌乳(Bole-Feysotetal.,1998)。研究發現,隨光照和溫度的變化,馬崗鵝Bostaurus血漿中PRL的濃度呈周期性變化(黃運茂等,2007)。本研究發現,8月THI值較高時,豚鹿血清中的LH、PRL含量極顯著低于5月、10月,FSH含量極顯著低于5月,表明豚鹿繁殖相關激素的分泌與光照時長和THI值相關,季節變化能夠在一定程度上影響豚鹿的繁殖效率。豚鹿繁殖激素的變化模式符合短光照繁殖類型動物隨光照延長、繁殖激素表達降低的基本特征,推斷豚鹿屬于短光照繁殖類型動物。光照延長后,豚鹿GnRH活化的同時也激發雌激素的負反饋作用,使機體進入非繁殖期(Knightetal.,1983;賀建寧等,2013),另外,夏季的高溫高濕脅迫也是限制豚鹿繁殖表現的重要因素。 本研究發現,豚鹿屬于短光照繁殖類型動物,夏季的高溫、高濕和長光照會影響豚鹿繁殖相關激素的分泌。由于圈養環境外在遮擋物較少,豚鹿直接受到強光和高濕熱脅迫而產生應激,建議在飼料中添加抗應激類添加劑,或改善圈舍條件,如增加遮光設施、增加植被和噴淋設施降低溫度等,以便緩解應激對機體生殖激素的逆向調節,在一定程度上提高其繁殖效率。 鄧家波, 余建秋, 牛李麗, 等. 2011. 豚鹿繁殖成活率低的病因學調查研究[J]. 野生動物, 32(3): 120-122. 狄冉, 郭曉飛, 劉秋月, 等.2015.生殖激素對綿羊繁殖性能影響的研究進展[J]. 家畜生態學報, 36(9): 1-6. 樊華, 樊麗, 王峰, 等. 2007. 熱應激對泌乳奶牛血液生化指標的影響[J]. 中國畜牧獸醫, 34(4): 45-46. 何世山, 金小軍. 2003. 高溫對肉雞血液生化指標的影響[J]. 浙江大學學報(農業與生命科學版), 29(3): 311-314. 賀建寧, 王金鑫, 狄冉, 等. 2013. 常年發情和季節性發情綿羊在不同季節生殖激素變化規律[J]. 畜牧獸醫學報, 44(10): 1547-1553. 黃運茂, 施振旦, 李孝偉, 等. 2007. 光周期對馬崗鵝產蛋、PRL和LH分泌季節性變化的影響[J]. 華南農業大學學報, 28(3): 94-96. 賴平, 王憑青, 張寶云, 等. 2012.哺乳動物季節性繁殖的神經內分泌調節機制[J]. 遺傳, 34(3): 281-288. 祁云霞, 何小龍, 劉曉芳, 等. 2014. 巴美肉羊發情期血清FSH和LH濃度變化規律及其與產羔數關系分析[J]. 黑龍江畜牧獸醫, (7): 17-20. 施振旦. 1995. 甲狀腺功能對調節雄性馬鹿體重和繁殖活動季節性變化的研究[J]. 獸類學報, 15(1): 17-24. 汪松. 1998. 中國瀕危動物紅皮書[M]. 北京: 科學出版社. 王訓立, 周建華, 范春梅. 2002. 季節性變化對雌性恒河猴生殖功能的影響[J]. 中國實驗動物學報, 10(1): 21-26. 魏學良, 張家驊, 王豪舉, 等. 2005. 高溫環境對奶牛生理活動及生產性能的影響[J]. 中國農學通報, 21(5): 13-15. 解美娜, 張才喬, 米玉玲, 等. 2004. 卵泡刺激素和雌激素對培養的雞胚卵巢生殖細胞增殖的影響[J]. 動物學研究, 25(1): 53-56. 楊海明. 2014. 揚州鵝繁殖的光照調控及下丘腦基因差異表達研究[D]. 江蘇揚州: 揚州大學. Bernal J. 2002. Action of thyroid hormone in brain[J]. Journal of Endocrinological Investigation, 25(3): 268-288. Bole-Feysot C, Goffin V, Edery M,etal. 1998. Prolactin and its receptor: actives, signal transduction pathways and phenotypes observed in PRL receptor knockout mice[J]. Endocrine Reviews, 19(3): 225-268. Clarke IJ. 1993. Variable patterns of gonadotropin-releasing hormone secretion during the estrogen-induced luteinizing hormone surge inovariectomized ewes[J]. Endocrinology, 133(4): 1624. Collier RJ, Eley RM, Sharma AK,etal. 1981. Shade management in subtropical environment for milk yield and composition in holstein and jersey cows 1[J]. Journal of Dairy Science, 64(5): 844-849. Knight PG, Cunningham FJ, Gladwell RT. 1983. Concentrations of immunoreactive luteinizing hormone releasing hormone in discrete brain regions of the cockerel: effects of castration and testosterone replacement therapy[J]. Journal of Endocrinology, 96(3): 471. Malpaux B, Tricoire H, Mailliet F,etal. 2002. Melatonin and seasonal reproduction: understanding the neuroendocrine mechanisms using the sheep as a model[J]. Reproduction Supplement, 59(1): 167-179. Sandy S. 2000. Is your dairy management program ready for the summer heat[D]. Texas: The Texas A & M University System. Silveira LFG, Teles MG, Trarbach EB,etal. 2010. Role of kisspeptin/GPR54 system in human reproductive axis[J]. Frontiers of Hormone Research, 39: 13-24. Smith JT, Coolen LM, Kriegsfeld LJ,etal. 2008. Variation in kisspeptin and R Famide-related peptide (RFRP) expression and terminal connections to gonadotropin-releasing hormone neurons in the brain: a novel medium for seasonal breeding in the sheep[J]. Endocrinology, 149(11): 5770-5782. Webster JR, Moenter SM, Woodfill CJI,etal. 1991. Role of the thyroid gland in seasonal reproduction: Ⅱ. Thyroxine allows a season-specific suppression of gonadotropin secretion in sheep[J]. Endocrinology, 129: 176-183. Wu J, Nayudu PL, Kiesel PS,etal. 2000. Luteinizing hormone has a stage-limited effect on preantral follicle development in vitro[J]. Biology of Reproduction, 63(1): 320-327. Yasuo S, Nakao N, Ohkura S,etal. 2006. Long-day suppressed expression of type 2 deiodinase gene in the mediobasal hypothalamus of the Saanen goat, a short-day breeder: implication for seasonal window of thyroid hormone action on reproductive neuroendocrine axis[J]. Endocrinology, 147(1): 432-440. Yokus B, Cakir DU, Kanay Z,etal. 2006. Effects of seasonal and physiological variations on the serum chemistry, vitamins and thyroid hormone concentrations in sheep[J]. Journal of Veterinary Medicine, 53(6): 271-276.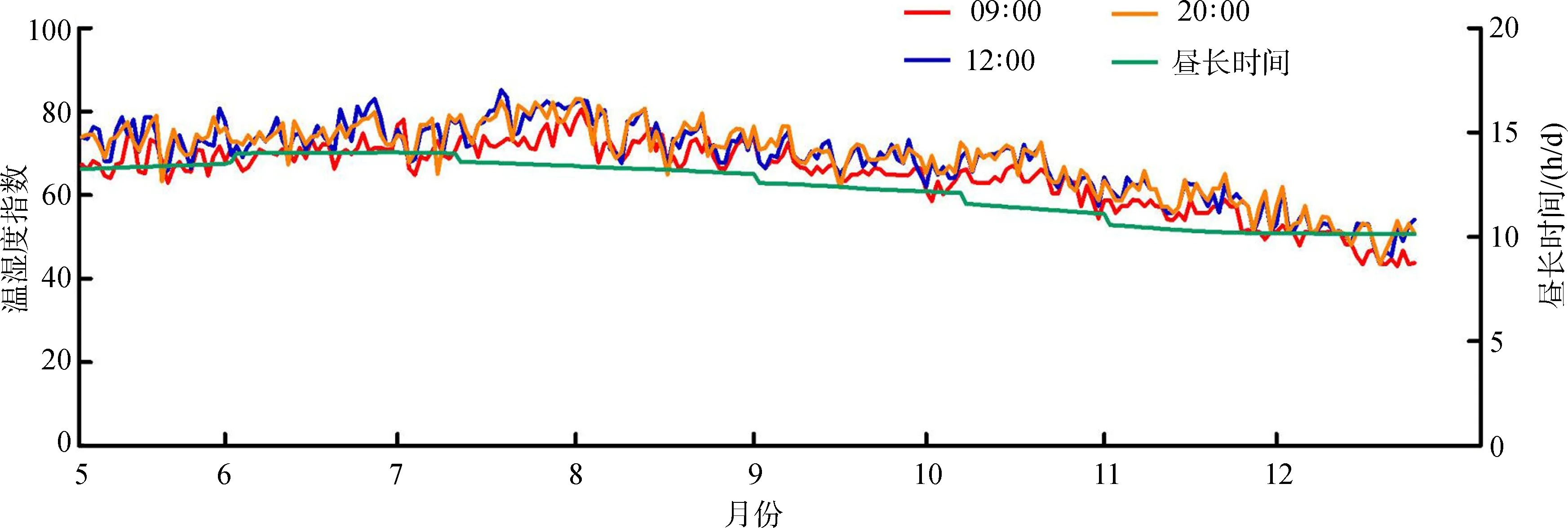
Fig. 1 The trend of temperature-humidity index in breeding houses ofAxisporcinusand daylength

2.4 各激素指標間層次聚類分析

3 討論
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
中老年保健(2021年3期)2021-08-22 06:50:04
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
昆明醫科大學學報(2021年1期)2021-02-07 01:06:36
現代臨床醫學(2021年1期)2021-01-26 00:56:02
中華養生保健(2020年4期)2020-11-16 01:31:40
中西醫結合肝病雜志(2020年2期)2020-10-27 02:18:50
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
