模擬氮沉降對高寒草甸土壤纖毛蟲群落的影響
2018-01-29 06:36:19寧應之張惠茹王芳國楊永強杜國禎
生態環境學報 2018年1期
寧應之,張惠茹,王芳國,楊永強,杜國禎
1. 西北師范大學生命科學學院,甘肅 蘭州 730070;2. 蘭州大學生命科學學院,甘肅 蘭州 730000
大氣污染已經成為當今世界亟待解決的重要問題之一,其中由于化石燃料的燃燒、汽車尾氣的排放和農業化肥的使用等情況日益嚴重,含氮化合物排放量急劇上升,導致氮沉降增加(韓雪等,2014),隨之引發一系列生態環境問題,例如森林植被衰退、土壤和水體酸化、物種多樣性降低等(Lu et al.,2014),這引起了科學家和公眾的密切關注。目前國內外學者對氮沉降的研究主要集中在其對森林生態系統、溫室氣體通量以及土壤環境因子的影響等方面(王暉等,2006;張煒等,2008;李秋玲等,2013)。關于氮沉降對土壤動物的影響,則多見于徐國良(2004,2005)、林英華(2010)、榮海等(2011)、周丹燕等(2015)對農田和森林生態系統中土壤動物群落的研究,而有關草原生態系統中原生動物受氮沉降影響的調查鮮有報道。
甘南位于青藏高原東部,是青藏高原向黃土高原和內陸盆地過渡的典型地區,因其獨特的地理單元和氣候特征而具有豐富的生物資源(劉旻霞,2017)。高寒草甸作為當地最主要的草地類型(牛鈺杰等,2017),是氮沉降等環境變化的敏感區域。土壤纖毛蟲是最復雜和最高等的單細胞真核生物,因其具有獨特的形態結構、生理功能和能對周圍環境變化做出迅速反應等特點(寧應之等,2017),在生態學研究領域有著不容忽視的意義。大氣氮沉降以無機態及有機態兩種形式發生,無機態所占比重較大,約為70%,且硝態氮和銨態氮為無機氮素的主要形態(程云云等,2017),因此本研究選取硝酸銨人工模擬氮沉降,以甘南高寒草甸為研究樣區,旨在揭示不同濃度氮肥作用下,土壤纖毛蟲群落及土壤環境因子的變異,并探討其相關性,以闡明氮沉降對其所產生的影響,為土壤原生動物對氮沉降的響應研究提供科學依據,補充氮沉降對生態系統影響的評價內容,為草原生態系統的良好穩定發展提供合理施肥的科學依據。
1 研究地區與研究方法
1.1 研究區概況
甘南藏族自治州瑪曲縣位于甘肅省西南部(圖1),地處甘、青、川三省交界地帶,地理坐標為100°45′~102°29′E,33°06′~34°30′N,海拔 3500~3800 m。該地區地勢西高東低,氣候屬典型的高原大陸性氣候,無明顯四季之分,地貌復雜。年平均氣溫1.1~2.7 ℃,年降水量約611.9 mm(包延東等,2017)。樣區主要土壤類型為高山草甸壤,呈微酸性至堿性(戚登臣等,2007);主要植被類型為高寒草甸,以莎草科(Cyperaceae)和禾本科(Gramineae)為主,多分布矮生嵩草(Kobresia humilis)、線葉嵩草(K. capillifolia)和披堿草(Elymus dahuricus)等。
圖1 研究區地理位置Fig. 1 Location of study area
1.2 研究方法
1.2.1 樣地設置
2009年在甘南瑪曲縣阿孜實驗站選取實驗樣地,設置16個(4×4)面積為60 m2(6 m×10 m)的小樣方,為防止干擾,各樣方間設置1 m緩沖地帶(不施肥),同時僅冬春季放牧。結合當地氮沉降情況,參照國際相關研究的處理方法(Gundersen et al.,1998),設計 4個不同水平的處理,即對照組(CK,不施氮肥)、低氮處理組(T5,5 g?m-2?a-1)、中氮處理組(T10,10 g?m-2?a-1)和高氮處理組(T20,20 g?m-2?a-1),每個處理組重復4次,隨機分布,保證每列每行都有4個不同水平氮沉降處理組。于次年5月底進行一次性氮沉降處理,所用肥料為顆粒狀NH4NO3,其有效氮含量約46%。為避免氮沉降不均產生斑塊異質而造成實驗誤差,將NH4NO3溶于水并均勻灑入樣地,CK噴灑等量自來水,各樣方除氮施用量不同外,其他處理保持一致。
1.2.2 樣品采集與處理
于2011年分季節完成4次采樣。采用用梅花五點取樣法,揀去土層表面凋落物,用15 mL圓筒形采樣器采集0~5 cm土壤層土樣25份,混勻裝袋并做好標記。采樣時觀察樣地植被類型,并用GPS測定儀測得樣區海拔和地理坐標。所得新鮮土樣部分用于測土壤含水量及 pH,剩余土樣倒入白瓷盤中,覆蓋透氣草紙使其自然風干備用,緊閉門窗以防空氣中的纖毛蟲包囊進入土壤而污染土樣(寧應之等,2007)。
1.2.3 鑒定方法和數據測定
(1)纖毛蟲的培養和鑒定
定性研究采用“非淹沒培養皿法”(non-flooded petri dish method),即在25 ℃下將50 g風干土樣置于直徑為15 cm的培養皿中,加土壤浸出液令其在充分浸潤但不淹沒的狀態下培養(Foissner,1992),之后用生物顯微鏡進行活體鏡檢分類,并配合 Wilbert蛋白銀染色法(Wilbert,1975)進行觀察,根據實驗記錄,依據Lynn(2008)分類系統對纖毛蟲進行分類。定量研究采用“3級10倍環式稀釋法”,即將風干土樣稀釋至10-1、10-2、10-3這3種濃度,并將由0.5 g NaCl+1.2 g瓊脂+98 mL蒸餾水配制而成的培養基趁熱倒入直徑為9 cm的培養皿中,快速埋入5個小玻璃杯,取3級土壤稀釋液各1 mL接種于各玻璃杯內,置于25 ℃下培養,之后分別在培養第 4、7、11天時鏡檢分類,根據密度換算表計算纖毛蟲數量(Stout,1962)。
(2)土壤理化因子的測定
土壤含水量采用烘干法進行測定(李廣文等,2009);土壤溫度于采樣時使用曲管地溫計(溫度范圍:-10~60 ℃)進行測量;土壤pH使用TSS-851土壤濕度酸度計進行測量(四平市電子技術研究所);土壤速效氮使用全自動流動注射儀進行測量(蘭州大學生命科學學院)。
1.3 數據處理
統計纖毛蟲物種數量并進行類群劃分,將物種數量占全部物種總數比例最多的目(order)定義為優勢類群,次多的目為次優勢類群,剩余各目均為常見類群(寧應之等,1998)。
對不同氮沉降處理組的相關指標進行單因素方差分析,采用LSD法比較顯著差異性分析;對土壤纖毛蟲物種數據進行氮處理與季節間無重復雙因子方差分析;對土壤纖毛蟲物種數據與環境因子進行冗余分析及Pearson相關性分析。所有數據均運用SPSS 23.0軟件進行處理;運用Primer 5.0進行Cluster及MDS分析;運用Canoco 4.5進行冗余分析;運用Arcview gis 9.3及GPS獲得的經緯度數據繪制樣區地圖;運用Origin 8.0繪制柱狀圖。
土壤纖毛蟲群落多樣性采用 Gleason-Margalef多樣性指數 d(沈韞芬等,1990)進行評價,具體計算公式如下:

式中,S為類群數(50 g實驗土樣中觀察到的所有物種數);N為群落中所有類群個體總數(纖毛蟲密度×土樣質量,土樣質量為50 g)。
C/P系數指纖毛蟲中r-對策者與K-對策者的比值,即腎形綱(Colpodea)纖毛蟲與異毛綱(Heterotrichea)、瓶纖綱(Armophorea)和旋毛綱(Spirotrichea)纖毛蟲的比值。它能夠在一定程度上評價生態環境質量狀況,在一般生態環境中C/P系數≤1,且其值越小表明生境越穩定、越良好(Foissner et al.,2005)。
2 結果與分析
2.1 土壤纖毛蟲群落特征對氮沉降的響應
2.1.1 群落組成
本研究在 4個樣點共鑒定到土壤纖毛蟲 121種,隸屬于9綱16目28科46屬(表1)。其中,優勢類群為散毛目(Sporadotrichida),共 3科 10屬27種,占物種總數的22.31%;次優勢類群為刺鉤目(Haptorida),共4科8屬24種,占物種總數的19.83%;常見類群有腎形目(Colpodida)、尾柱蟲目(Urostylida)、籃環目(Cyrtolophosidida)、齒管目(Chlamydodontida)、前管目(Prorodontida)、帆口目(Pleuronematida)、排毛目(Stichotrichida)、四膜目(Tetrahymenida)、游仆蟲目(Euplotida)、瓶纖目(Armophorida)、固著目(Sessilida)、籃口目(Nassulida)、嗜污目(Sathrophilus)和異毛目(Heterotrichida)等14類,共計70種,占物種總數的57.85%。
各處理組纖毛蟲群落組成也存在顯著差異,如圖2所示,CK纖毛蟲類群數最為豐富,且隨著氮沉降量量的增加,各處理組纖毛蟲群落組成趨于簡單化。CK 及 T5優勢類群為散毛目(Sporadotrichida);T10優勢類群為散毛目和腎形目(Colpodida);而T20優勢類群僅有腎形目,這表明氮沉降處理在使纖毛蟲群落組成簡單化的同時,也使得分布其中的纖毛蟲物種也相應低等化,由高等復雜的腹毛類纖毛蟲變為低等適應性更強的腎形類纖毛蟲。
2.1.2 物種分布
經統計,各處理組土壤纖毛蟲物種分布情況為:CK共分布68種,占物種總數的56.20%,物種最為豐富;T5和T10次之,分別具53、43種,占物種總數的43.80%、35.54%;T20物種分布最少,僅24種,占物種總數的19.83%。回歸分析表明,隨氮沉降量增加,纖毛蟲物種數顯著減少,回歸方程為 ?=-0.4599x+30.364(R2=0.9881,P=0.006)。
應用 Primer 5.0對各處理組的物種分布進行Cluster和MDS分析,結果見圖3。在歐氏距離為0.8相似性水平上各處理組被分為三類,T5與 T10組合在一起,CK和T20各自獨成一組。在歐氏距離為1.3相似性水平上各處理組又被聚為兩類,T5、T10與CK聚為一組,T20獨成一組。MDS分析結果與 Cluster一致。這表明 T5與 T10物種分布最為相似,它們與CK物種分布較相似,而T20與其他處理組差異很大。
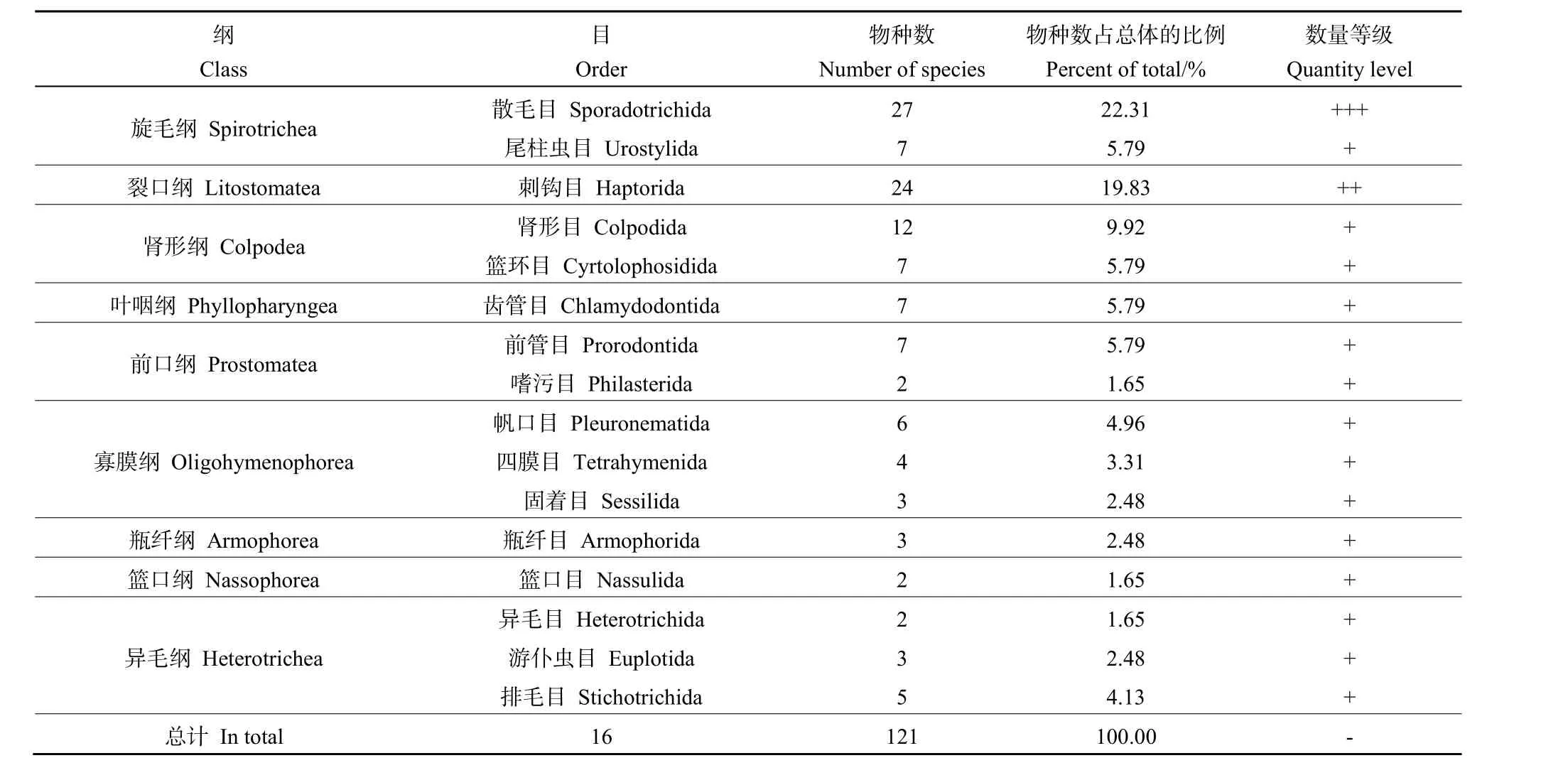
表1 纖毛蟲群落組成及數量等級Table 1 Ciliate community composition and quantity level
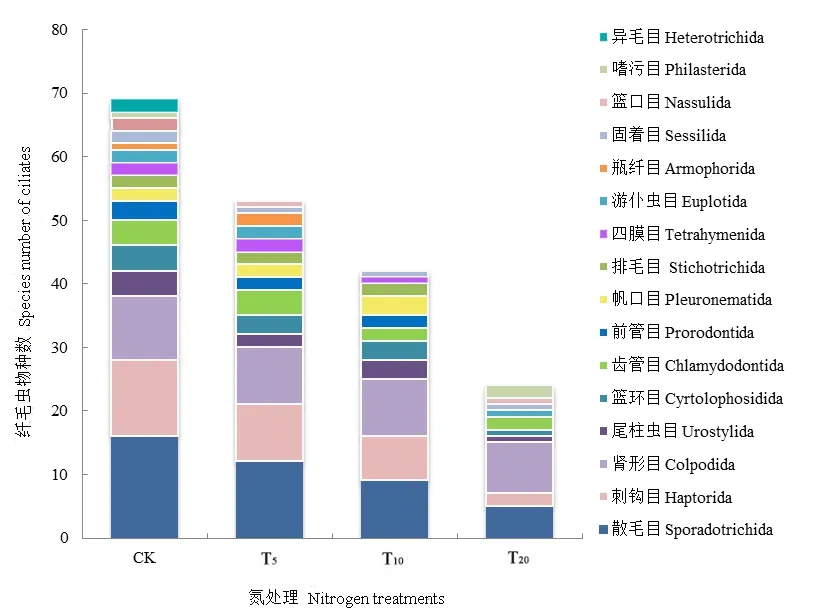
圖2 各處理組土壤纖毛蟲群落組成Fig. 2 The community composition of soil ciliates in various treatment group
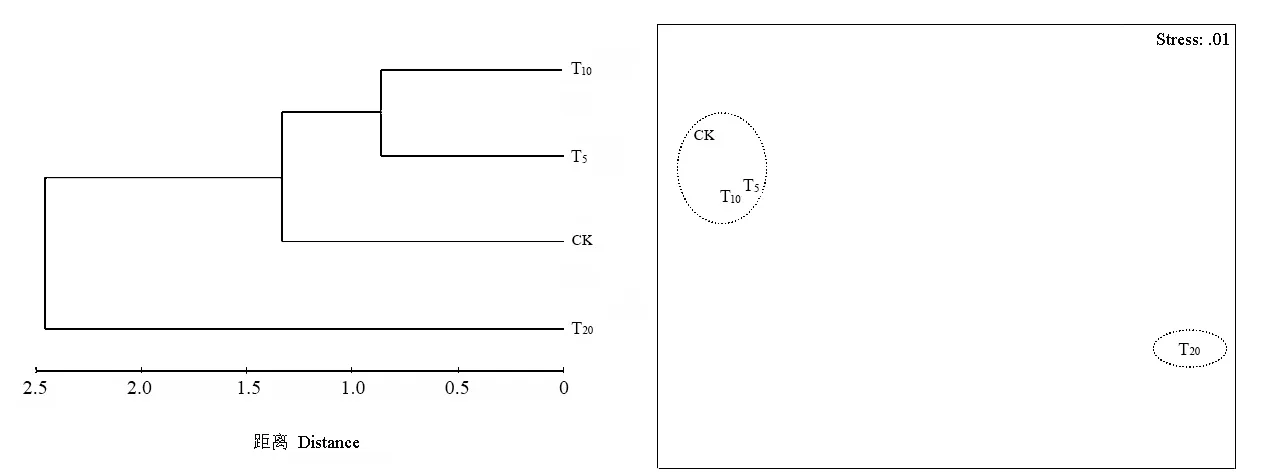
圖3 各處理組土壤纖毛蟲物種分布的Cluster和MDS分析Fig. 3 The cluster and MDS analysis on species distribution of soil ciliates in various treatment group
2.1.3 優勢種
將出現頻次較高的種定義為優勢種。就各處理組的物種分布而言,CK優勢種為小尖毛蟲(Oxytricha minor)、野生假膜蟲(Notohymena selvatica)、剛強殖口蟲(Gonostomum strenuum)、粘游仆蟲(Euplotes muscicola)、鈍單鐮蟲(Drepanomonas obtusa);T5優勢種為剛強殖口蟲(G. strenuum)、顆粒尖毛蟲(O. granulifera)和鈍單鐮蟲(D. obtusa);T10優勢種為顆粒尖毛蟲(O.granulifera)、僧帽腎形蟲(Colpoda cucullus)和粗糙腎形蟲(C. aspera);T20優勢種單一,為僧帽腎形蟲(C. cucullus)和土壤腎形蟲(C. edaphoni)。隨著氮沉降量增加,優勢種也從腹毛類纖毛蟲演替為腎形類纖毛蟲,且種類數逐漸減少。
2.1.4 廣布種和特有種
廣布種即為所有處理組中均出現的種,共 12種,占物種總數的9.92%。特有種指僅在一個處理組中出現的物種,CK中分布最多,共39種,占物種總數的 32.23%;T5分布 29種,占物種總數的23.97%;T10分布12種,占物種總數的9.92%;T20僅分布2種,占物種總數的1.65%。各處理組中特有種數目變化與優勢種一致,即 CK>T5>T10>T20,回歸分析也表明,隨著氮肥處理濃度的增加,特有種種類數顯著減少,回歸方程為?=-0.4976x+18.951(R2=0.9383,P=0.031)。
2.1.5 C/P系數
CK、T5、T10的 C/P系數分別為 0.54、0.63、0.86,而T20的C/P系數大于1,說明CK中以K-型腹毛類纖毛蟲占優勢,而 T20中以 r-型腎形類纖毛蟲為主,表明土壤環境狀況隨氮沉降量增加逐漸趨于惡劣,高氮處理嚴重破壞了土壤環境。
2.2 土壤環境因子對氮沉降的響應
如表2所示,氮沉降處理對土壤含水量、土壤溫度和速效氮的影響差異均不顯著,但對pH具有顯著影響(P<0.05)。各氮沉降處理組土壤含水量均高于對照組,且隨著氮沉降量增大呈上升趨勢,4個季節均以 T20土壤含水量最高。氮沉降處理對土壤溫度的影響與含水量呈相反趨勢,即隨著氮沉降量增加,土壤溫度下降,且其隨季節變化呈現出夏季最高、冬季最低的特征。與對照組相比,氮沉降處理使pH下降了0.03~0.83個單位,沉降量越高其下降幅度越大。氮沉降處理對速效氮的影響總體表現為隨沉降量的增加,速效氮含量也增加,而在各季節表現出不同的趨勢。
2.3 土壤纖毛蟲物種數、密度及多樣性指數對氮沉降的響應
土壤纖毛蟲物種數、密度及多樣性指數均隨不同水平氮沉降處理發生了不同程度的改變。如圖 4所示,土壤纖毛蟲物種數在各季節均表現為對照組(CK)顯著高于氮沉降處理(P<0.05),而在各氮沉降處理組中纖毛蟲物種數又隨著氮沉降量的增加呈下降趨勢,T20中觀察到的物種數最少。這與榮海等(2011)、徐國良等(2005)對單施氮肥對土壤動物生物量的影響研究結果相一致,即高濃度氮處理會引起土壤動物類群數的下降。
土壤纖毛蟲密度隨著氮沉降量增加表現出不同的趨勢,T5(春:138.50;夏:194.58;秋:269.58;冬:341.50)和T10(春:163.25;夏:195.58;秋:341.67;冬:384.92)纖毛蟲密度顯著高于CK(春:123.67;夏:165.58;秋:146.67;冬:198.08)(P<0.05),且各季節均以 T10纖毛蟲密度為最大。除夏季外,T20(春:143.67;夏:98.33;秋:156.42;冬:218.50)纖毛蟲密度均略大于 CK,但差異不顯著,這表明低水平氮處理對纖毛蟲密度表現為促進作用,而高水平下促進作用減弱,存在閾值效應。
不同水平氮處理對Gleason-Margalef多樣性指數的影響與物種數一致,即隨著氮沉降量的增大,多樣性指數顯著下降(P<0.05),CK(春:3.45;夏:3.99;秋:3.49;冬:4.58)多樣性指數最高,T5(春:2.95;夏:3.26;秋:2.96;冬:3.90)次之,T10(春:2.57;夏:2.73;秋:2.68;冬:3.35)再次,T20(春:1.58;夏:1.53;秋:1.80;冬:2.05)最低。
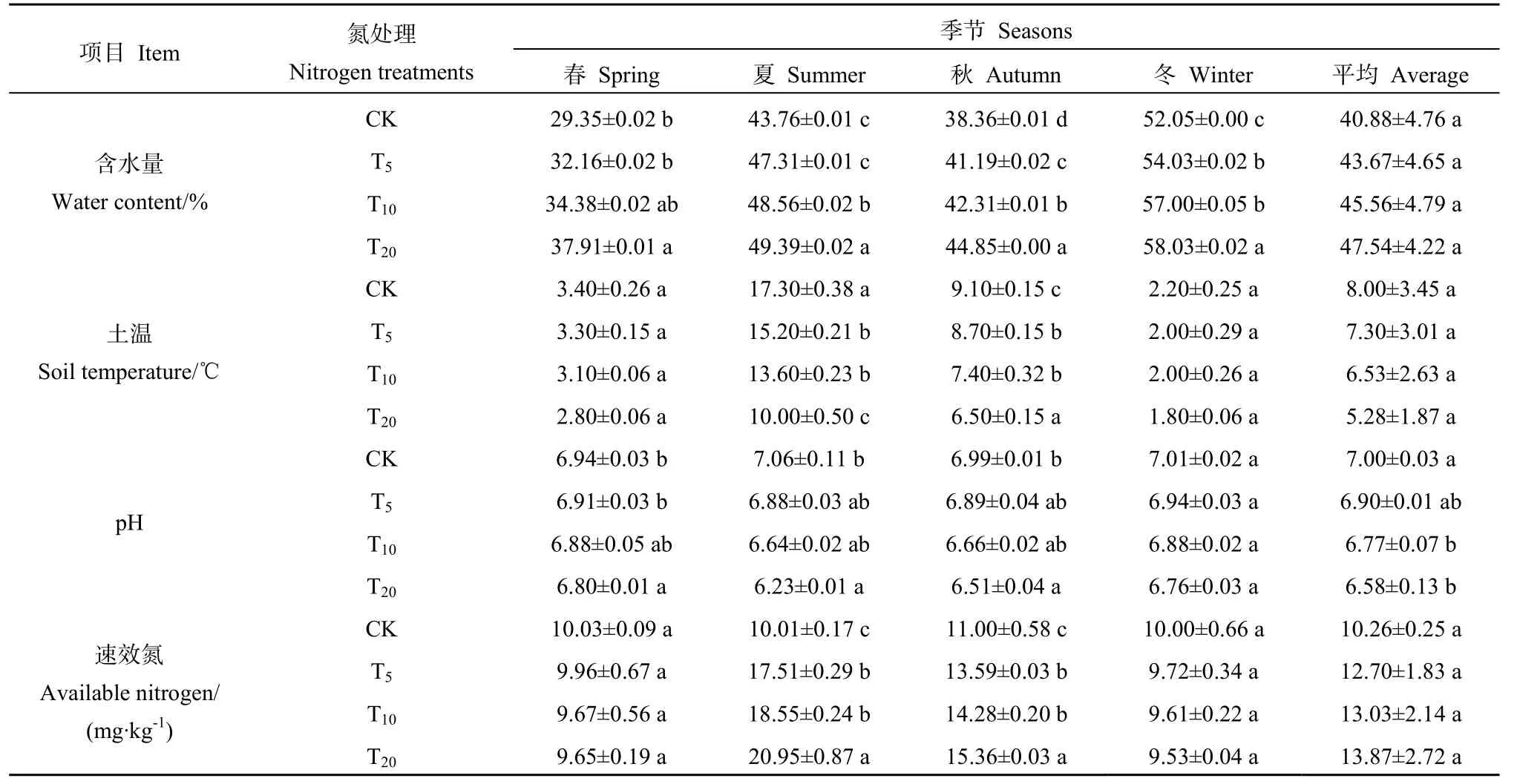
表2 各處理組環境因子Table 2 Environmental factors in various treatment group
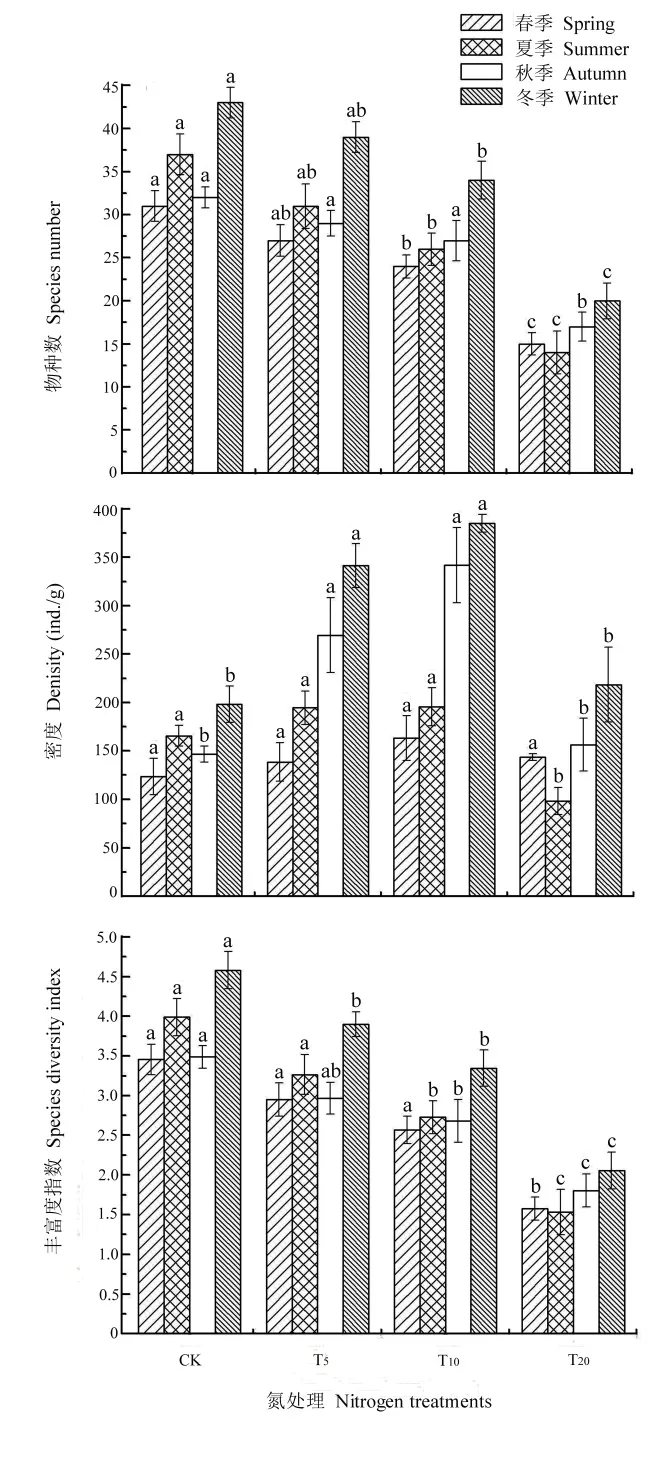
圖4 各處理組纖毛蟲物種數、密度和多樣性指數Fig. 4 Species number, density, species diversity index in various treatment group
對土壤纖毛蟲物種數、密度及多樣性指數進行氮處理與季節間的無重復雙因子方差分析,結果顯示,土壤纖毛蟲物種數對氮處理(P=0.000,F=12.424)和季節(P=0.000,F=16.168)的響應差異顯著;密度對氮處理(P=0.000,F=3.867)和季節(P=0.000,F=20.750)的響應差異也顯著;多樣性指數對氮處理(P=0.000,F=12.414)和季節(P=0.000,F=10.515)的響應同樣差異顯著。
2.4 土壤環境因子對不同氮沉降處理土壤纖毛蟲的影響
2.4.1 土壤纖毛蟲物種數據與土壤環境因子的RDA分析
為確定合適的模型并進行排序分析,首先對物種數據進行DCA分析,即非約束性排序-去趨勢對應分析,得知結果中 4個軸的最大梯度為 0.212,故采用線性模型-冗余分析(RDA)探討纖毛蟲物種數據與環境因子之間的關系。如表3所示,通過對環境因子進行蒙特卡羅置換檢驗(Monte Carlo Permutation Test),結果顯示,含水量與pH是對纖毛蟲群落具有顯著影響的環境因子,通過手動預選對其與纖毛蟲物種數據進行 RDA分析,提取到 2個有效排序軸(表 4),特征值分別為 0.5390和0.1140,累計貢獻率達到65.3%,因此RDA排序分析一定程度上可反映土壤纖毛蟲物種數據與環境因子之間的相關關系。第1排序軸與土壤含水量和土壤pH都呈正相關,第2排序軸與土壤含水量呈正相關,與土壤pH呈負相關。
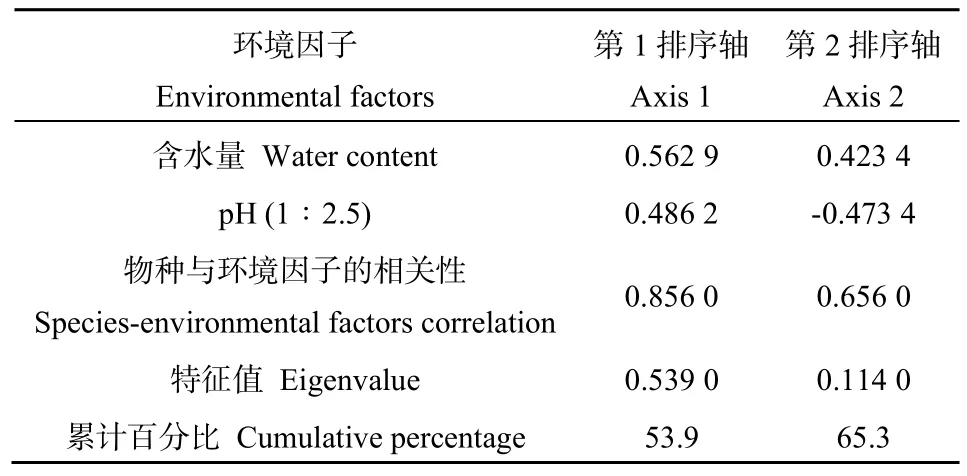
表4 環境變量與排序軸的相關系數Table 4 Correlation coefficients between environmental factors with ordination axes of RDA
圖5所示為反映土壤環境因子與纖毛蟲物種數據的RDA二維排序圖,圖中數字1~4代表氮沉降量依次增加的春季各處理組,同理 5~16代表氮沉降量依次增加的夏、秋、冬季各處理組。根據各射線夾角可知,纖毛蟲密度與土壤含水量、纖毛蟲物種數與土壤pH都呈正相關,相關性較強,纖毛蟲多樣性指數與土壤含水量和土壤pH均呈負相關。分析樣點與環境因子間的相互關系可知,對照組與低氮處理組樣點多分布于環境因子同側,與環境因子關系更為密切,因此,RDA排序圖可以很好地反映出纖毛蟲物種參數與環境因子之間的關系。
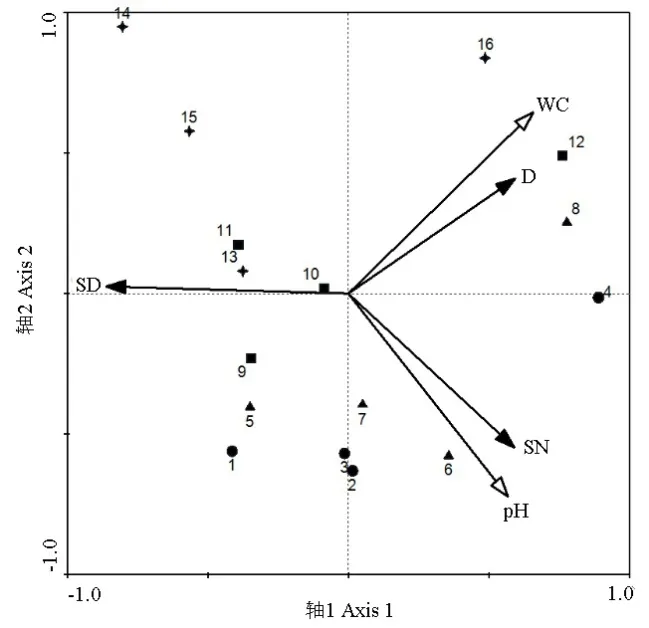
圖5 物種-環境因子相關RDA排序圖Fig. 5 RDA analysis ordination diagram of species-environment relationships
2.4.2 土壤纖毛蟲物種數據與土壤環境因子的相關性分析
運用SPSS 23.0對土壤纖毛蟲物種數據與環境因子進行Pearson相關性分析(Pearson correlation analysis),結果如表5所示,在氮處理下,土壤含水量與物種多樣性指數呈極顯著負相關(P=0.004),與密度呈顯著正相關(P=0.036);土壤 pH與物種數呈極顯著正相關(P=0.001);其他各因子之間相關性不顯著。這與上述 RDA冗余分析的結果相吻合。綜上所述,通過對各樣點進行不同水平氮處理,該研究樣區的環境因子發生了不同程度的改變,其中含水量與pH為起主導作用的因素,可以對纖毛蟲的群落特征參數產生不同程度的影響。
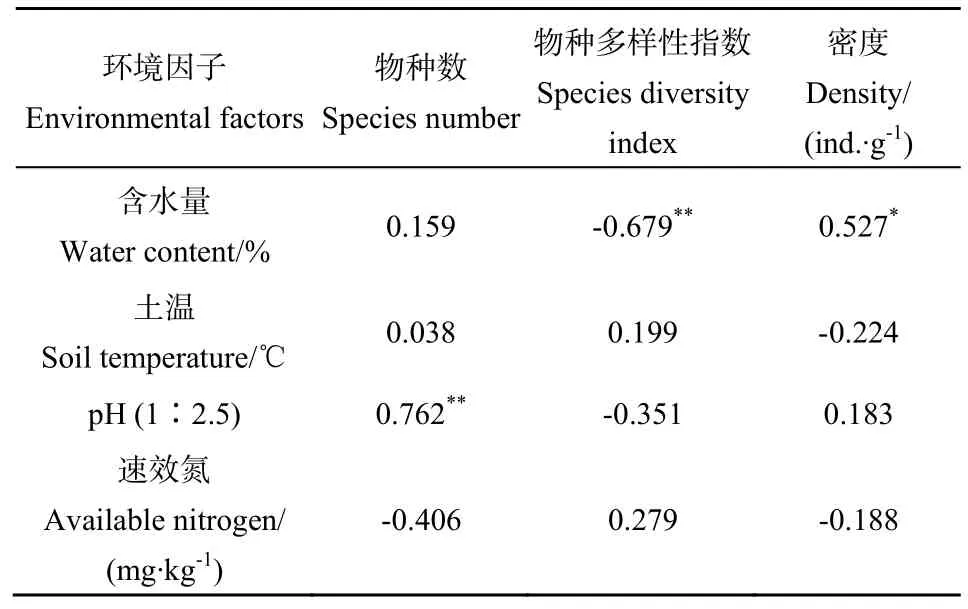
表5 纖毛蟲物種數據與環境因子的Pearson相關性分析Table 5 Relationships between species and environmental factors using Pearson?s correlation coefficient
3 討論
3.1 氮沉降對土壤環境因子的影響
人工模擬氮沉降對研究區土壤環境因子產生了不同程度的影響,結果顯示,氮處理對土壤 pH的影響最為顯著(P<0.05),對土壤含水量、土壤溫度和速效氮的影響均不顯著。氮沉降處理使土壤pH顯著下降,且氮沉降量越高,下降幅度越大,這與齊莎等(2010)、周丹燕等(2015)研究結果相一致。究其原因,過量的NO3-除通過反硝化作用或淋溶作用散失外,剩余部分被滯留在土壤中(肖輝林,2001),這可能使土壤處于氮“過飽和”狀態,從而發生強烈的酸化作用;其次,Foster et al.(1989)研究表明,土壤中Ca2+的凈損失會導致土壤酸化,而由氮沉降引起的土壤中 NO3-的過量會導致土壤中 Ca2+和 Mg2+等陽離子的淋失加重,故土壤隨氮沉降量的增加呈明顯弱酸性。各氮沉降處理組土壤含水量隨氮沉降量增加呈上升趨勢,這主要是由于各處理組土壤速效氮成分隨著氮沉降量增大而升高,且植被覆蓋率隨植被總生物量的升高有所增加(周丹燕等,2015),從而提高了土壤保水性;而各氮沉降處理土壤含水量的季節變化均表現為冬季>夏季>秋季>春季,這與研究區所處的自然環境密切相關。氮沉降處理對土壤溫度的影響與含水量呈相反趨勢,即氮沉降量越高,土壤溫度越低,但差異不顯著;土壤溫度也隨季節變化呈現夏季最高、冬季最低的特征。氮沉降處理對土壤速效氮含量的影響在各季節間表現出不同趨勢:春季各處理組速效氮含量無顯著變化,這是由于研究區春季氣溫較低,植被生長及微生物活動相對較弱,單施氮肥對土壤速效氮含量影響并不顯著;夏、秋兩季速效氮含量隨氮沉降量加大而增大,這是由于隨著植被生長逐漸旺盛,微生物對腐殖質的分解等作用加強,且人為添加硝酸銨,使土壤中的硝態氮和氨態氮含量上升;而在冬季,氮沉降處理組速效氮含量相較于對照組有所下降,這是由于集中降雨期(5—10月)過后,淋溶作用加強,且植被的生長消耗了大量速效氮。
3.2 氮沉降對土壤纖毛蟲群落的影響
隨氮沉降量增大,各處理組土壤纖毛蟲物種數呈線性顯著減少(P<0.05),這與已有研究結果相一致,齊莎等(2010)認為,連續5年氮處理后,土壤生物量及其多樣性均顯著降低,在土壤原生動物中,土壤纖毛蟲對氮處理的響應最為積極。根據前文對土壤纖毛蟲物種數據與環境因子之間的 RDA分析和Pearson相關性分析可知,土壤pH與纖毛蟲物種數呈極顯著正相關(P<0.01)。人工模擬氮沉降后,土壤環境發生變化,其中最為顯著的是土壤pH降低,導致土壤酸化,氮沉降量越高,pH下降幅度越大,T20處理組土壤pH均值為6.58,而最適宜纖毛蟲生長的土壤 pH在 6.7~7.0之間(沈韞芬,1999),因此高氮處理組中纖毛蟲物種數最低。此外,徐國良等(2003)指出氮沉降物性質也可對土壤動物的生長和繁殖產生直接作用,如酸沉降物與土壤動物直接接觸可致其死亡。
氮沉降處理對纖毛蟲密度的影響表現為 CK<T20<T5<T10,其中,T10纖毛蟲密度最大,這表明氮沉降對纖毛蟲密度的影響具有閾值效應,當氮沉降量控制在一定范圍內時,對纖毛蟲密度表現為促進作用,但當氮沉降量過高時,促進作用減弱。RDA分析與 Pearson相關性分析表明,土壤含水量與物種密度呈顯著正相關(P<0.05),隨氮沉降量的增加,各處理組土壤含水量呈上升趨勢,因此纖毛蟲密度在低氮及中氮處理后增加,但在高氮處理后有所下降,這是由于過量的氮肥使土壤中速效氮含量上升,降低了水分的有效性;再者,邱波等(2004)研究表明,單施氮肥會降低植被多樣性,作為土壤原生動物最重要的棲息地,植被根系多樣性的破壞也會使其密度降低。
纖毛蟲多樣性指數表現為 CK>T5>T10>T20,即隨著氮沉降量增加,各處理組纖毛蟲群落組成趨于簡單化,優勢種和特有種單一且都為具有良好適應能力的腎形蟲,纖毛蟲由較為高等的K-型纖毛蟲演替為簡單低等的r-型纖毛蟲。究其原因,腹毛類纖毛蟲屬于纖毛蟲中較為高級的階元,個體大,只有當環境良好時才可以大量生存,而高氮處理使其受到環境脅迫,因此在 T20處理組中腹毛類纖毛蟲分布最為罕見;其次,該類群主要攝食細菌、藻類及其他原生動物,氮沉降量處理在使土壤環境發生變化的同時,也會使土壤團聚體及土壤溶液滲透壓發生改變,從而導致土壤微生物活動量減弱,轉入休眠狀態甚至死亡(宇萬太等,2008),最終改變土壤微生物群落結構(張海芳等,2017)。腎形類纖毛蟲是土壤纖毛蟲中最常見的三大類群之一,除對攝食要求較低外,該類群適應能力極強,具有完善而迅速的脫包囊和形成包囊機制,因而能存活于氮沉降后較為惡劣的環境中,甚至經過長期適應,擁有較強的耐受性而成為高氮處理組的優勢類群。隨氮沉降量增加,纖毛蟲物種數降低而密度增加,這也是纖毛蟲多樣性指數呈下降趨勢的直接原因。
4 結論
本研究表明,在甘肅甘南高寒草甸人工模擬氮沉降后,高氮處理對纖毛蟲群落組成具有消極影響,即高濃度氮肥不利于土壤纖毛蟲群落物種多樣性的提高,且對土壤纖毛蟲密度的影響具有閾值效應。氮沉降處理通過影響土壤環境因子進而改變土壤纖毛蟲群落結構,其主要制約因子為土壤含水量和pH。為維持穩定良好的生態系統功能,推薦5~10 g?m-2?a-1氮肥施用量作為高寒草甸最佳施肥水平參考值。氮沉降對生態系統的影響是一個長期漸進的過程,由于本試驗研究周期較短,其對高寒草甸土壤纖毛蟲群落的長期作用有待對多個影響因素、多地點及多時間點展開更深入的研究。
致謝:英國自然歷史博物館 Alan Warren(E-mail:a.warren@nhm.ac.uk)研究員為本文的英文摘要進行了潤色,特此致謝!
FOISSNER W, BERGER H, XU K, et al. 2005. A huge, undescribed soil ciliate (Protozoa: Ciliophora) diversity in natural forest stands of Central Europe [J]. Biodiversity & Conservation, 14(3): 617-701.
FOISSNER W. 1992. Estimating the species richness of soil protozoa using the “non-flooded petridish method” [C]//Lee J J, Soldo ATods.Protocols in Protozoology. Lawrence: Allen Press: 10.1-10.2.
FOSTER N W, HAZLETT P W, NICOLSON J A, et al. 1989. Ion leaching from a sugar maple forest in response to acidic deposition and nitrification [J]. Water, Air, & Soil Pollution, 48(1-2): 251-261.
GUNDERSEN P, EMMETT B A, KJONAAS O J, et al. 1998. Impact of nitrogen deposition on nitrogen cycling in forests: a synthesis of NITREX data [J]. Forest Ecology and management, 101(1): 37-55.
LU X, MAO Q, GILLIAM F S, et al. 2014. Nitrogen deposition contributes to soil acidification in tropical ecosystems [J]. Global change biology,20(12): 3790-3801.
LYNN D. 2008. The ciliated protozoa: characterization, classification, and guide to the literature [M]. Berlin: Springer Science & Business Media:1-605.
STOUT J D. 1962. An estimation of microfaunal populations in soils and forest litter [J]. European Journal of Soil Science, 13(2): 314-320.
WILBERT N. 1975. An improved technique of protargol impregnation for ciliates [J]. Mikrokosmos, 64(6): 171-179.
包延東, 劉長仲. 2017. 甘南州天然草原生態環境現狀及恢復治理對策[J]. 甘肅畜牧獸醫, 47(3): 108-109.
程云云, 孫濤, 王清奎, 等. 2017. 模擬氮沉降對溫帶森林土壤線蟲群落組成和代謝足跡的影響[J]. 生態學報, 38(2): 1-10.
韓雪, 王春梅, 藺照蘭. 2014. 模擬氮沉降對溫帶森林凋落物分解的影響[J]. 生態環境學報, 23(9): 1503-1508.
李廣文, 王永潔, 羅金明, 等. 2009. 融凍時期扎龍濕地土壤含水量變化研究[J]. 高師理科學刊, 29(6): 71-74.
李秋玲, 肖輝林, 曾曉舵, 等. 2013. 模擬氮沉降對森林土壤化學性質的影響[J]. 生態環境學報, 22(12): 1872-1878.
林英華, 黃慶海, 劉驊, 等. 2010. 長期耕作與長期定位施肥對農田土壤動物群落多樣性的影響[J]. 中國農業科學, 43(11): 2261-2269.
劉旻霞. 2017. 甘南高寒草甸植物元素含量與土壤因子對坡向梯度的響應[J]. 生態學報, 37(24): 1-10.
寧應之, 沈韞芬. 1998. 中國典型地帶土壤原生動物: I. 區系特征和物種分布[J]. 動物學報, 44(1): 5-9.
寧應之, 王娟, 劉娜, 等. 2007. 甘肅天水麥積山風景名勝區土壤纖毛蟲的物種多樣性[J]. 動物學研究, 28(4): 367-373.
寧應之, 楊元罡, 蘇萇, 等. 2017. 放牧對沼澤化草甸土壤纖毛蟲群落的影響[J]. 生態學雜志, 36(6): 1614-1621.
牛鈺杰, 周建偉, 楊思維, 等. 2017. 基于地形因素的高寒草甸土壤溫濕度和物種多樣性與初級生產力關系研究[J]. 生態學報, 37(24): 1-12.
戚登臣, 李廣宇. 2007. 黃河上游瑪曲濕地退化現狀, 成因及保護對策[J]. 濕地科學, 5(4): 341-347.
齊莎, 趙小蓉, 鄭海霞, 等. 2010. 內蒙古典型草原連續5年施用氮磷肥土壤生物多樣性的變化[J]. 生態學報, 30(20): 5518-5526.
邱波, 羅燕江, 杜國禎. 2004. 施肥梯度對甘南高寒草甸植被特征的影響[J]. 草業學報, 13(6): 65-68.
榮海, 范海蘭, 李茜, 等. 2011. 模擬氮沉降對農田大型土壤動物的影響[J]. 東北林業大學學報, 39(1): 85-88.
沈韞芬, 章宗涉, 龔循矩, 等. 1990. 微型生物監測新技術[M]. 北京: 中國建筑工業出版社: 1-524.
沈韞芬. 1999. 原生動物學[M]. 北京: 科學出版社: 1-656.
王暉, 莫江明, 薛璟花, 等. 2006. 氮沉降增加對森林凋落物分解酶活性的影響[J]. 熱帶亞熱帶植物學報, 14(6): 539-546.
肖輝林. 2001. 大氣氮沉降對森林土壤酸化的影響[J]. 林業科學, 37(4):111-116.
徐國良, 莫江明, 周國逸, 等. 2003. 土壤動物與N素循環及對N沉降的響應[J]. 生態學報, 23(11): 2453-2463.
徐國良, 莫江明, 周國逸, 等. 2004. 模擬氮沉降對苗圃地土壤動物群落的影響[J]. 生態環境, 13(4): 487-492.
徐國良, 莫江明, 周國逸. 2005. 模擬氮沉降增加對南亞熱帶主要森林土壤動物的早期影響[J]. 應用生態學報, 16(7): 1235-1240.
宇萬太, 姜子紹, 周樺, 等. 2008. 不同土地利用方式對潮棕壤微生物量碳及其周轉率的影響[J]. 生態學雜志, 27(8): 1302-1306.
張海芳, 劉紅梅, 趙建寧, 等. 2017. 貝加爾針茅草原土壤真菌群落結構對氮素和水分添加的響應[J]. 生態學報, 38(1): 1-11.
張煒, 莫江明, 方運霆, 等. 2008. 氮沉降對森林土壤主要溫室氣體通量的影響[J]. 生態學報, 28(5): 2309-2319.
周丹燕, 卜丹蓉, 葛之葳, 等. 2015. 氮添加對沿海不同林齡楊樹人工林土壤動物群落的影響[J]. 生態學雜志, 34(9): 2553-2560.
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
媽媽寶寶(2017年3期)2017-02-21 01:22:28
中國塑料(2016年3期)2016-06-15 20:30:00
通信電源技術(2016年3期)2016-03-26 07:13:38
