海水脅迫對白子菜生長及生理特性的影響
2018-01-12 11:39:17覃和業徐微風王永壯符少萍郭建春段瑞軍
江蘇農業科學 2017年24期
王 蕾, 覃和業, 徐微風, 劉 姣, 王永壯, 符少萍, 郭建春, 段瑞軍
(1.中國熱帶農業科學院/熱帶生物技術研究所,海南海口 571101;2.海南大學,海南海口 571101)
隨著環境條件的惡化,土壤鹽漬化逐漸成為一個全球性的生態問題。相關研究表明,鹽漬化土壤占全球耕地面積的7%,其中約有20%的耕地正受到次生鹽漬化的威脅[1-2]。土壤中過高的鹽分易造成植物細胞結構的損傷,嚴重抑制植物的生長,甚至會導致植物大面積死亡,極大地影響了現代農業的生產及可持續發展[3]。隨著全球性糧食、資源、生態環境問題的日趨嚴重,沿海灘涂植被顯示出越來越大的生態及經濟效益,海水蔬菜的開發及利用已經成為改良土壤鹽漬化、維持生態平衡的重要途徑,也是目前應對鹽漬化危機的主要方法[4-5]。目前,北美海蓬子、堿蓬、番杏、中亞濱藜等耐鹽植物已被報道用于鹽漬土的改良[6]。
白子菜[Gynuradivaricata(L.) DC.]別稱接骨丹、百子菜、茹童菜、雞菜、大肥牛白背菜、散血姜、明月草等,屬菊科三七屬,為多年生草本植物,主要分布于我國華南地區、云南及越南北部等地區[7]。白子菜根系發達,繁殖能力強,可扦插,扦插5~7 d即可成活,適宜在含鹽量高的沙地種植[8]。此外,白子菜抗逆性強、耐高溫,適宜在海南地區種植。白子菜藥用及營養價值較高,嫩莖可食用,味甘、淡,寒,根入藥有清熱瀉火之效[9]。可用于清熱解毒、活血通經、舒筋接骨、涼血止血,對支氣管肺炎、小兒高熱、百日咳、目赤腫痛、風濕關節痛、崩漏、跌打損傷、血瘀腫痛、骨折風濕骨痛、外傷出血癰瘡腫毒、乳腺炎、瘡瘍疔腫、燒燙傷等具有明顯的療效[10]。當前對白子菜的研究多集中在栽培技術、藥用價值等方面,關于其在鹽脅迫下生長及生理響應的研究國內外尚未有報道。本試驗通過海水長期處理白子菜,研究不同海水濃度處理對白子菜的生長及相關生理指標的影響,為進一步利用白子菜對濱海灘涂地、鹽荒地、海水倒灌農田的改良和利用提供理論基礎。
1 材料與方法
1.1 試驗材料
白子菜采自海南省陵水黎族自治縣分界洲島,將嫩莖扦插于塑料花盆中進行育苗,插后4~7 d即能發生不定根,12~15 d后定植于透氣的塑料花盆中。海水取自海南省海口市白沙門,鹽濃度為3.0%。
1.2 試驗方法
1.2.1 材料處理 本試驗于2016年3月在中國熱帶農業科學院熱帶生物技術研究所內進行,設置6個海水濃度(0、10%、20%、30%、40%、50%),3次重復。當扦插的白子菜長出4~6張新葉后進行移栽,選取長勢一致的幼苗定植于塑料花盆中,每盆3株,澆透水。試驗時在06:00—7:00間土壤溫度較低時進行海水處理,同時配以Hoagland 營養液,每隔5 d澆1次海水,每隔20 d澆1次Hoagland營養液,當表層沙土 1~2 cm發白時及時補澆自來水,使土壤含水量達到田間最大含水量的60%~70%。栽培基質選用篩過的細河沙。90 d后,測定其相關生理形態指標等。
1.2.2 形態指標的測定 各處理分別取白子菜3株,用直尺測量其株高(cm)、節間長(cm),用游標卡尺測定其莖粗(cm)、葉片厚度(cm),用紫光平臺掃描儀(LA2000)結合ImageJ軟件測定葉面積[11]。
1.2.3 生理指標的測定 超氧化物歧化酶(簡稱SOD)活性的測定參照李合生等的氮藍四唑光化還原法[12-13];過氧化物酶(簡稱POD)活性的測定參照朱廣廉等的愈創木酚法[14];過氧化氫酶(簡稱CAT)活性的測定參照彭志英等的方法[15];脯氨酸含量的測定參照鄭炳松的茚三酮法[16]并有改進;可溶性糖含量的測定參照鄭炳松的蒽酮比色法[16];細胞膜透性參照劉寧等的方法[17-18]測定質膜相對透性;丙二醛(簡稱MDA)含量測定參照Heath等的硫代巴比妥酸(簡稱TBA)比色法[19];根系活力的測定參照張憲政的2,3,5-三苯基氯化四氮唑(TTC)法[20];葉綠素含量的測定采用張憲政的丙酮乙醇混合提取法[20]。
1.2.4 數據分析 采用Excel 2003整理試驗數據、繪制圖表;用SPSS 13.0進行方差分析和多重比較,進行α=0.05水平的統計顯著性檢驗。
2 結果與分析
2.1 海水脅迫對白子菜生長的影響
為研究海水處理對白子菜生長的影響,本試驗測定不同濃度海水處理下白子菜的形態指標(株高、莖粗、節間數、葉片厚度、葉面積、地上部生物量)。由表1可知,白子菜在0~30%海水濃度處理下,各生長指標相較于對照均未受到明顯抑制,且在≤20%海水濃度處理下,大多數指標隨海水濃度的提高有不同程度的增加;株高、節間數、葉面積在海水濃度≥40%時均顯著減少;而地上部生物量在10%~20%海水濃度脅迫下有所增加,當海水濃度≥40%時,地上部生物量顯著減少,說明當海水濃度在0~20%時在一定程度上促進了白子菜的生長,當海水濃度≥40%時,白子菜生長受海水抑制作用顯著。
2.2 海水脅迫對白子菜細胞膜透性的影響
2.2.1 海水脅迫對白子菜細胞膜透性的影響 電導率是反映細胞膜透性的一個非常重要的指標。白子菜在未經海水處理下,其根、莖、葉中的細胞膜透性分別是40.25%、87.50%、85.64%。由圖1可知,在海水脅迫下,白子菜根莖葉中膜透性整體隨脅迫的增加而升高,但在莖中無顯著性升高;在40%海水濃度脅迫下葉片中電導率顯著增加,其余各濃度處理下無顯著性變化;根系中的變化尤為明顯,在≥10%海水處理下,電導率顯著增加。說明海水脅迫對根系的損傷遠遠大于莖、葉。
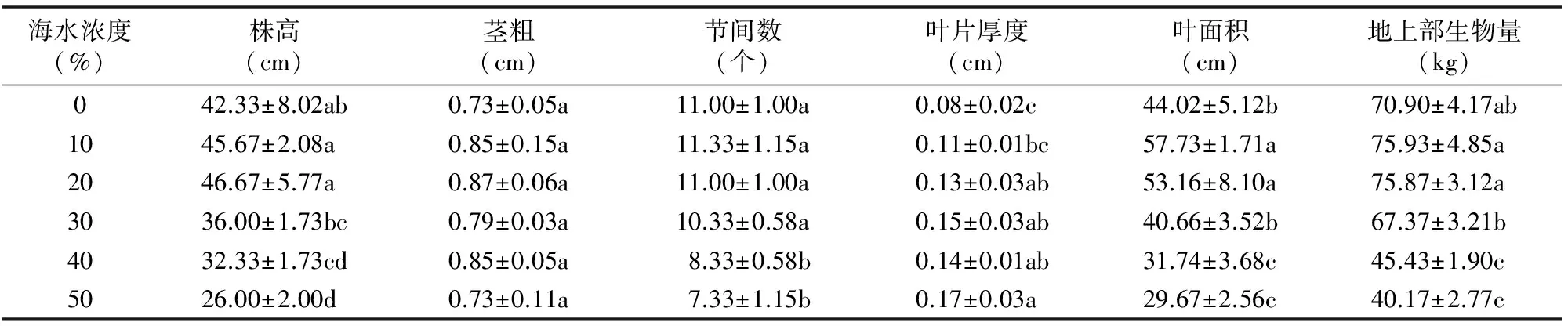
表1 海水處理對白子菜生長指標的影響
注:同列數據后不同小寫字母表示差異顯著(P<0.05)。
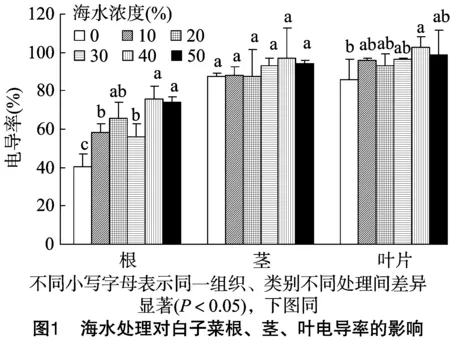
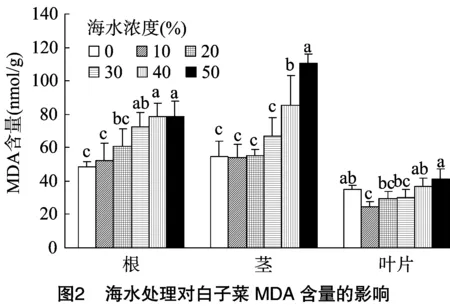
2.2.2 海水脅迫對白子菜MDA含量的影響 MDA是膜脂過氧化作用的主要產物之一,可間接表示細胞膜的受損程度,并兼有反饋的作用。在未經海水處理下,白子菜根、莖、葉片中MDA的含量分別為48.26、54.36、34.70 nmol/g。由圖2可知,白子菜MDA含量在根、莖中隨海水濃度的增加整體增加,分別在≥30%、≥40%海水處理下MDA含量顯著高于對照。在葉片中,MDA含量呈先減少后增加的趨勢,當海水處理濃度≤30%時,MDA含量較對照略有下降,但隨著海水濃度的升高MDA含量逐步上升,最大值達到41.20 nmol/g,較對照增加了18.7%。說明在低濃度下海水對白子菜細胞膜的損傷較小,而在高濃度下細胞膜受到海水毒害作用明顯增加。
心律失常患者選擇射頻導管消融術治療能夠有效的改善患者心功能,幫助患者緩解臨床癥狀,促進患者健康的恢復。但在使用射頻導管消融術治療中,手術中可能對患者和術者產生的輻射危害是一種潛在的風險,有可能對患者和醫生的身體造成不良影響,為了減輕治療過程中的輻射危害,在手術治療過程中合理采取防護措施幫助降低輻射十分關鍵[1]。為了具體分析心律失常射頻導管消融中的放射防護措施及效果,本文選取2016年12月—2017年12月在我院接受射頻導管的心律失常患者72例進行分組研究如下。
2.3 海水脅迫對白子菜滲透調節物質的影響
2.3.1 海水脅迫對白子菜脯氨酸含量的影響 植物細胞的脯氨酸含量增加,維持了細胞的正常膨壓,同時還可以保護酶的活性和使細胞膜系統免受傷害。白子菜在未使用海水處理下,其根、莖、葉中的脯氨酸含量分別為2.32、5.28、2.04 μg/g。由圖3可知,海水脅迫下白子菜根、莖、葉中脯氨酸含量的變化趨勢整體基本相同,都是隨著脅迫濃度的增加而增加,在低濃度脅迫下脯氨酸含量變化不顯著。在≥30%海水處理下,根中的脯氨酸含量顯著增加,海水濃度最高時其含量最高,是對照的15.37倍;在≥20%海水處理下,莖、葉片中的脯氨酸含量顯著增加,濃度最高時含量最高,分別是對照的6.21、1.95倍。說明白子菜在低濃度的海水脅迫下對脯氨酸含量變化無顯著性影響,而在高濃度海水脅迫下較為敏感。

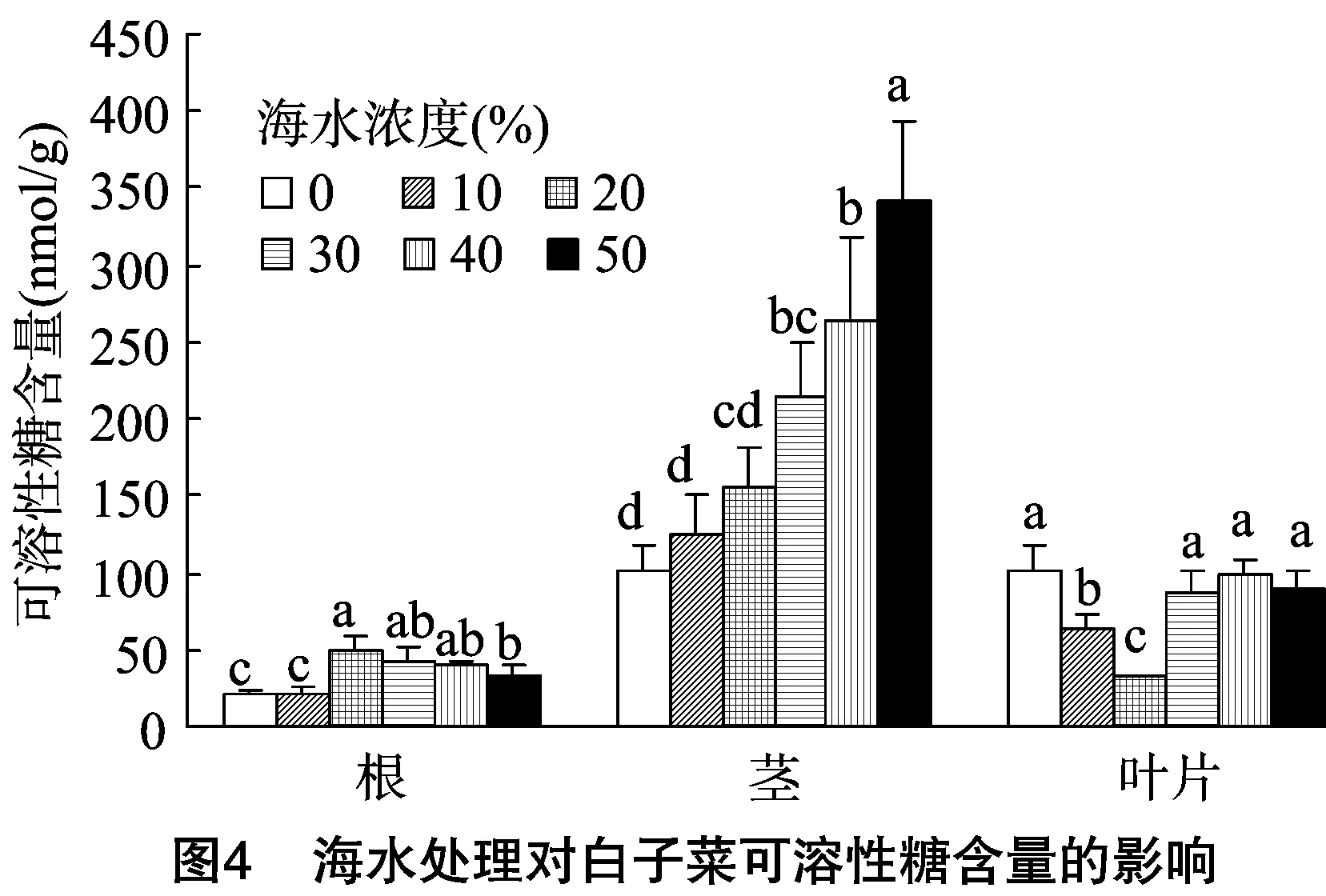
2.3.2 海水脅迫對可溶性糖含量的影響 可溶性糖作為植物重要的滲透調節物質,在逆境環境中其含量的增加有利于增強植物對環境的適應能力。由圖4可知,白子菜在未經海水脅迫處理下,根、莖、葉中可溶性糖含量分別為20.98、101.79、100.66 nmol/g。在葉片中,可溶性糖含量隨著海水濃度的升高呈先下降后上升的趨勢,在20%海水濃度處理下達到最低值,較對照減少了14.5%;在30%~50%海水處理下,其含量略有提高。在根中,可溶性糖含量隨著海水濃度的升高呈先增加后略有下降的趨勢,在20%海水濃度處理下含量最高,為對照的2.38倍;在30%~50%海水濃度處理下略有下降,分別為對照的2.00、1.95、1.61倍。在莖中可溶性糖含量呈穩定增長趨勢,在50%海水處理下達到最大值,為342.60 nmol/g,是對照的 3.37 倍,表明海水脅迫促進了可溶性糖在白子菜根莖中的積累。
2.4 海水脅迫對白子菜不同組織抗氧化酶系統的影響



2.4.2 海水脅迫對白子菜POD活性的影響 由圖6可知,在未經海水脅迫處理下,白子菜根、莖、葉中POD活性分別為208.14、476.85、220.93 U/g。白子菜POD活性對海水脅迫較敏感,根、莖中分別在10%、20%海水脅迫下,POD活性較對照組顯著增加,分別達到526.8、729.4 U/g,分別較對照增加了153.3%、50.8%,且隨海水濃度的增加POD 活性增加,根中POD活性在50%海水濃度處理下達到最大值。葉中POD活性呈先增加后減小的變化趨勢,除20%海水脅迫下顯著增加,其余各個濃度與對照組相比沒有顯著性變化。POD活性的增加,有助于清除體內的H2O2,從而增強白子菜的耐鹽性。

2.4.3 海水脅迫對白子菜CAT活性的影響 由圖7可知,在未經海水處理下,白子菜根、莖、葉片中CAT活性分別為86.93、28.60、72.21 U/g。白子菜根中CAT活性隨處理濃度的升高而升高,在10%~20%的海水處理下,其活性無明顯增加,在30%~50%濃度下,活性明顯增強,分別比對照增加了26.4%、26.2%、41.6%。莖、葉片中的變化趨勢相似,在10%~30%濃度處理下,CAT活性顯著增加,且30%處理濃度時達到最大值,分別是對照的1.66、1.93倍。在高濃度的海水脅迫下,莖、葉片中的CAT活性下降,但仍然略高于對照,說明白子菜中CAT活性能對海水脅迫作出響應。
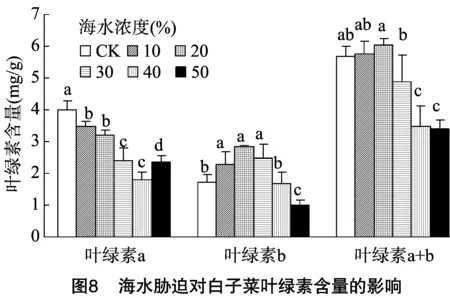
2.5 海水脅迫對白子菜葉綠素含量的影響

2.6 海水脅迫對白子菜根系活力的影響
根系活力是衡量植物根系吸收、合成、抵抗逆境能力大小的重要生理指標[23]。由圖9可知,在未經海水處理下,白子菜根系活力為350.251 μg/(g·h)。在海水脅迫≤30%濃度處理下,根系活力保持在較高水平,且在10%處理濃度下,根系活力較對照顯著增加了18.76%,在40%、50%濃度處理下,根系活力與對照相比下降,分別比對照減少了21.41%、12.03%。所以,低濃度的海水脅迫提高了白子菜的根系活力,且能忍受較高濃度的海水脅迫。

3 結論與討論
植物在逆境脅迫下細胞內自由基代謝平衡遭到破壞,導致膜脂過氧化,使細胞膜系統嚴重受損。MDA是膜脂過氧化產物,能與細胞內各種成分發生反應,進而引起膜的損傷,其含量的高低可間接表現植物受逆境脅迫的強弱[24-25]。前人研究發現,在中低鹽度脅迫下細胞膜透性增加不明顯,而在高鹽度脅迫下迅速增加,本研究也獲得了相似結果,白子菜在≤30%海水脅迫下,MDA含量較對照略有升高,增加不明顯,表明在此濃度范圍內,白子菜對海水脅迫具有一定的適應性,而在≥40%海水脅迫下,MDA含量在根、莖中顯著增加,其電導率的變化與MDA趨勢基本相同,說明高鹽脅迫下白子菜的細胞膜結構受到損傷,細胞內外的穩定性遭到破壞,造成植物細胞內的電解質滲漏增強,并隨脅迫濃度的增加而增加。
脯氨酸和可溶性糖是植物體內重要的滲透調節物,有研究者將其作為植物抗鹽的重要生理指標[26-27]。譚大海等發現,蘆葦受鹽脅迫時,能夠大量迅速地累積脯氨酸來降低細胞滲透勢,使植物體對鹽分具有一定的耐受性[28]。本研究中白子菜體內的脯氨酸含量隨鹽濃度的升高大量積累,并且濃度越高積累越迅速,根莖中的積累量大于葉片中的,表明游離脯氨酸的積累是白子菜適應鹽脅迫的重要途徑之一,根的適應性高于葉片,這是植物為了對抗鹽脅迫而采取的一種保護性滲透調節反應。可溶性糖為有機物質的合成提供碳架和能量來源,能穩定細胞膜,是重要的滲透調節物質[29]。在本研究中,白子菜根莖中的可溶性糖含量隨海水處理濃度的升高整體呈上升的趨勢,說明植物可通過可溶性糖的積累來增強滲透調節能力,而在葉片中表現為先減小后增加的趨勢,可能是由于低濃度的海水脅迫使白子菜的生長維持在較高的水平,并且海水中一些礦物元素促進了植物的吸收,而在高濃度(≥30%)脅迫下,其含量顯著增加。高鹽脅迫下,可溶性糖含量增加也有可能是由于高濃度鹽脅迫下植物生長受抑制,導致糖利用減少,可溶性糖含量增加[30]。白子菜中脯氨酸與可溶性糖含量的增加,有助于維持細胞正常的膨壓,有利于細胞生理生化過程的正常進行;保持細胞持續生長,以減輕逆境對細胞生長的抑制;對于保持原生質體與環境的平衡和膜結構的完整性具有重要的作用,此外,還可以維持生物膜的穩定性和某些酶的活性。
植物在逆境脅迫下,會產生大量對植物有強烈毒害作用的活性氧,破壞植物體的動態平衡,使膜透性增加,代謝紊亂,此時植物會啟動細胞內的保護酶(SOD、CAT、POD等)系統,從而及時清除體內由于氧化脅迫產生的大量自由基來維持正常的代謝[31-32]。SOD、POD和CAT是酶保護系統中的重要組成部分,可以清除過氧化物、過氧化氫和氧自由基,避免或減輕植物受到傷害,使植物正常生長代謝[33]。本研究中,在低濃度海水(≤30%)處理下,3種酶活性都有升高,表明植物對海水脅迫作出了反應。高濃度海水(≥40%)處理的白子菜根中SOD、POD、CAT活性維持在較高水平,而莖、葉中SOD活性較對照無明顯變化,說明白子菜各組分因海水的脅迫產生了大量的活性氧,抑制了活性氧清除酶活性,SOD、POD、CAT協同作用下降,活性氧的產生與清除失去平衡,保護酶活性的變化與MDA含量變化相一致。
葉綠素參與植物的光合作用,是光合作用的重要物質,逆境脅迫會抑制植物葉綠素合成的相關酶類,導致葉綠素生成受阻、含量下降,葉綠素功能結構受損、氧化分解增強。本研究中,葉綠素a與葉綠素b的變化不一致,說明二者對鹽害的適應性不同,在海水濃度≤30%時,葉綠素b含量增加,葉綠素a含量減少,而葉綠素總量無顯著減少,表現出較強的忍耐能力。這可能是由于低鈉鹽可以增加植物體內的葉綠素含量,而高濃度海水脅迫破壞了植物葉綠體結構,使體內葉綠素含量下降,引起植株光合能力減弱[34-37]。
植物的根系活力能夠客觀地反映根系生命活動力,可以衡量植物根系的吸收和合成能力也是判斷植物根系抵御逆境脅迫能力的重要生理指標,能夠反映植物抗鹽能力[38-40]。本研究中,在≤30%海水濃度處理下,白子菜根系能夠保持較高的活力,但隨著海水脅迫濃度(≥40%)的增高,根系受到損傷,根系活力較對照下降,但下降不明顯。說明白子菜具有較強的耐鹽性,這與張潤花等對黃瓜[41]、陳炳東等對油葵的研究結果[42]相似。
綜上,白子菜在海水濃度為0~30%時各項生理活動能正常進行,白子菜的生長未受到明顯抑制,能正常生長;海水中豐富的礦質營養使得在≤20%海水濃度脅迫下生物量等指標增加。通過對各項生理指標的綜合分析發現,白子菜在海水濃度≤30%時表現正常,其原因為(1)白子菜抗氧化酶(SOD、POD、CAT)的高活性,能最大限度地清除因海水脅迫產生的大量活性氧,減輕活性氧對白子菜細胞的損傷。(2)脯氨酸和可溶性糖積累,維持了白子菜的細胞滲透壓。(3)根系活力處于較高水平有助于營養和水分的吸收,保障了地上部的營養供應。(4)葉綠素含量無明顯減少,因而植物能進行正常的光合作用,維持正常的生理活動。當海水濃度≥40%時,白子菜葉綠素含量、根系活力降低抑制了白子菜的光合作用與營養物質的運輸,膜系統受到的毒害作用增加,MDA大量積累,保護酶協同清除活性氧作用下降,白子菜生長受到顯著抑制。
[1]Aroca R,Ruiz-Lozano J M,ZamarreoM,et al. Arbuscular mycorrhizal symbiosis influences strigolactone production under salinity and alleviates salt stress in lettuce plants[J]. Journal of Plant Physiology,2013,170(1):47-55.
[2]Li J G,Pu L J,Han M F,et al. Soil salinization research in China:advances and prospects[J]. Journal of Geographical Sciences,2014,24(5):943-960.
[3]Estrada B,Aroca R,Barea J M,et al. Native arbuscularmycorrhizal fungi isolated from a saline habitat improved maize antioxidant systems and plant tolerance to salinity[J]. Plant Science,2013,201(1):42-51.
[4]Lieth H. Concepts for different uses of halophytes[M]// Mangroves and halophytes:restoration and utilisation. Berlin:Springer Netherlands,2008:3-5.
[5]楊成龍,段瑞軍,李瑞梅,等. 鹽生植物海馬齒耐鹽的生理特性[J]. 生態學報,2010,30(17):4617-4627.
[6]王 蕾,吳朝波,徐微風,等. 海水脅迫對番杏生長、滲透調節物質和細胞膜透性的影響[J]. 江蘇農業科學,2016,44(7):248-251.
[7]昊 霞,張錢錢,王忠震,等. 白子菜葉片中酸性多糖的降血糖作用及其對相關指標的影響[J]. 植物資源與環境學報,2015,24(2):115-117.
[8]楊 暹,劉厚誠. 白子菜的特征特性及栽培技術[J]. 廣東農業科學,1998(5):16-17.
[9]關佩聰,吳琰芬. 8 種野菜的特征、生境、食法及保健作用[J]. 長江蔬菜,1999(11):28-30.
[10]李小軍,覃 歡,劉 歡. 菊三七屬植物的民族藥用和食用價值[J]. 中南民族大學學報(自然科學版),2015,34(4):62-67.
[11]戴志聰,杜道林,司春燦,等. 用掃描儀及 Image J 軟件精確測量葉片形態數量特征的方法[J]. 廣西植物,2009,29(3):342-347.
[12]李合生,孫 群,趙世杰,等. 植物生理生化實驗原理和技術[M]. 北京:高等教育出版社,2000.
[13]郝建軍,康宗利,于 洋. 植物生理學實驗技術[M]. 北京:化學工業出版社,2007.
[14]朱廣廉,鐘海文,張愛琴. 植物生理學實驗[M]. 北京:北京大學出版社,1990:37-40.
[15]彭志英,蔣 黎. 紫外速率直接法測定過氧化氫酶活性[J]. 華西醫學,1995,10(1):4-7.
[16]鄭炳松. 現代植物生理生化研究技術[M]. 北京:氣象出版社,2006:3-120.
[17]劉 寧,高玉葆. 滲透脅迫下多花黑麥草葉內過氧化物酶活性和脯氨酸含量以及質膜相對透性的變化J]. 植物生理學通訊,2000,36(1):11-14.
[18]陳建勛,王曉峰. 植物生理學實驗指導[M]. 2版. 廣州:華南理工大學出版社,2006:64-66.
[19]Heath R L,Parke L. Photoperitation in isolated diloroplastskinstics and stoichimertry fatty acid peroxidation [J]. Archbiophys,1968,75:189-198.
[20]張憲政. 作物生理研究法[M]. 北京:農業出版社,1992.
[21]陳少裕. 膜脂過氧化對植物細胞的傷害[J]. 植物生理學通訊,1991,27(2):84-90.
[22]張玉霞,李志剛,李美娟,等. 四種草地早熟禾抗鹽堿生理生化特性的研究[J]. 中國農學通報,2004,20(5):209-213.
[23]Yanagawa H A,Watanabe K,Nakamura M. Application of the feed ingredients for livestock to an artificial diet by using polyphagous strain of the silkworm[J]. The Journal of Sericultural Science of Japan,1989,58(5):401-406.
[24]華 春,周泉澄,王小平,等. 外源GA3對鹽脅迫下北美海蓬子種子萌發及幼苗生長的影響[J]. 南京師大學報(自然科學版),2007,30(1):82-87.
[25]孫海菁,王樹鳳,陳益泰. 鹽脅迫對6個樹種的生長及生理指標的影響[J]. 林業科學研究,2009,22(3):315-324.
[26]劉占彬,袁慶華,景啟美. 9個高羊茅品種苗期耐鹽性研究[J]. 中國草地學報,2007,29(6):81-85.
[27]馬宗仁,郭 博. 短芒披堿草和老芒麥在水分脅迫下游離脯氨酸積累的研究——Ⅰ. 植物的抗旱性與脯氨酸積累能力關系的標準[J]. 中國草地,1991,4:12-16.
[28]譚大海,沙 偉,張瑩瑩. 蘆葦鹽脅迫下滲透調節物質含量變化研究[J]. 齊齊哈爾大學學報(自然科學版),2006,22(2):84-86.
[29]李圓圓,郭建榮,楊明峰,等. KCl和NaCl處理對鹽生植物堿蓬幼苗生長和水分代謝的影響[J]. 植物生理與分子生物學學報,2003,29(6):576-580.
[30]Nixon P J,Mullineaux C W.Regulation of photosynthetic electron transport[M]//Regulation of Photosynthesis. Springer Verlag:Springer Netherlands,2001:533-555.
[31]Ouariti O,Boussama N,Zarrouk M,et al. Cadmium-and copper-induced changes in tomato membrane lipids[J]. Phytochemistry,1997,45(7):1343-50.
[32]Kanazawa S,Sano S,Koshiba T,et al. Changes in antioxidative enzymes in cucumber cotyledons during natural senescence:comparison with those during dark-induced senescence[J]. Physiologia Plantarum,2001,109(2):211-216.
[33]劉愛榮,張遠兵,陳登科. 鹽脅迫對鹽芥(Thellungiellahalophila)生長和抗氧化酶活性的影響[J]. 植物研究,2006,26(2):216-221.
[34]Munns R,Termaat A. Whole-plant responses to salinity[J]. Functional Plant Biology,1986,13(1):143-160.
[35]Matoh T,Murata S. Sodium stimulates growth of Panicum coloratum through enhanced photosynthesis[J]. Plant Physiology,1990,92(4):1169-1173.
[36]汪貴斌,曹福亮. 鹽分和水分脅迫對落羽杉幼苗的生長量及營養元素含量的影響[J]. 林業科學,2004,40(6):56-62.
[37]吳永波,薛建輝. 鹽脅迫對3種白蠟樹幼苗生長與光合作用的影響[J]. 南京林業大學學報(自然科學版),2002,26(3):19-22.
[38]王素平,郭世榮,胡曉輝,等. 鹽脅迫對黃瓜幼苗葉片光合色素含量的影響[J]. 江西農業大學學報,2006,28(1):32-38.
[39]宋海星,王學立. 玉米根系活力及吸收面積的空間分布變化[J]. 西北農業學報,2005,14(1):137-141.
[40]高福元,張吉立,劉振平,等. 鹽脅迫對樹錦雞兒葉綠素含量和根系活力的影響[J]. 貴州農業科學,2010,38(5):46-48.
[41]張潤花,郭世榮,李 娟. 鹽脅迫對黃瓜根系活力、葉綠素含量的影響[J]. 長江蔬菜,2006(2):47-49.
[42]陳炳東,黃高寶,陳玉梁,等. 鹽脅迫對油葵根系活力和幼苗生長的影響[J]. 中國油料作物學報,2008,30(3):327-330.
猜你喜歡
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
