雷帕霉素對自閉癥仔鼠神經、社交行為及海馬區BDNF、Bcl-2蛋白表達的影響
2018-01-06 02:32:10黨媛媛倪金鳳劉國玉張潔商艷朝
山東醫藥 2017年48期
黨媛媛,倪金鳳,劉國玉,張潔,商艷朝
(唐山市婦幼保健院,河北唐山063000)
雷帕霉素對自閉癥仔鼠神經、社交行為及海馬區BDNF、Bcl-2蛋白表達的影響
黨媛媛,倪金鳳,劉國玉,張潔,商艷朝
(唐山市婦幼保健院,河北唐山063000)
目的探討雷帕霉素對自閉癥仔鼠神經、社交行為及海馬區腦源性神經營養因子(BDNF)、Bcl-2蛋白表達的影響。方法將12只Wistar孕鼠分為對照組、丙戊酸鈉組(模型組)和丙戊酸鈉聯合雷帕霉素組(觀察組),每組4只。模型組和觀察組妊娠12.5天時腹腔注射丙戊酸鈉(600 mg/kg),對照組同期腹腔注射等量生理鹽水;觀察組腹腔注射丙戊酸鈉(600 mg/kg)后,每天給予雷帕霉素(4 mg/kg)灌胃直至仔鼠斷奶,每組仔鼠20只。仔鼠出生35天時,分別檢測三組仔鼠神經行為(在中央區的活動時間和直立次數)、社交行為(社交行為時間及非社交行為時間)及海馬區BDNF、Bcl-2表達。結果與對照組仔鼠比較,模型組及觀察組仔鼠在中央區活動時間均增加,站立次數均減少;與模型組仔鼠比較,觀察組仔鼠在中央區活動時間減少,站立次數增加;組間比較P均<0.05。與對照組仔鼠比較,模型組及觀察組仔鼠社交行為時間均減少,非社交行為時間均增加;與模型組仔鼠比較,觀察組仔鼠社交行為時間增加,非社交行為時間減少;組間比較P均<0.05。模型組仔鼠海馬BDNF蛋白相對表達量低于對照組,Bcl-2蛋白相對表達量高于對照組;觀察組仔鼠海馬BDNF蛋白相對表達量高于模型組,Bcl-2蛋白相對表達量低于模型組;組間比較P均<0.05。結論雷帕霉素可改善丙戊酸鈉自閉癥仔鼠模型神經行為表現,提高其社交能力;促進海馬區BDNF表達、抑制Bcl-2表達,調節神經元異常發育、促進細胞凋亡可能是其作用機制。
自閉癥;雷帕霉素;神經行為;社交行為;腦源性神經營養因子;Bcl-2;仔鼠
自閉癥又稱孤獨癥,是一種廣泛性發育障礙疾病,一般在嬰幼兒期發病,主要表現為言語溝通障礙、社會交往(簡稱社交)障礙以及行為刻板等,發病機制尚未明確,其發病率為2/1 000~6/1 000[1]。腦源性神經營養因子(BDNF)屬于神經營養因子家族成員,在各腦區均有分布,可調控神經系統發育,其生理功能紊亂或電信號傳導障礙均可引起精神發育遲緩、自閉癥、腦萎縮和阿爾茨海默病等腦部疾病[4,5]。神經細胞凋亡障礙可能是自閉癥的發病機制之一[6,7]。目前臨床上缺乏治療自閉癥的有效藥物。研究發現,雷帕霉素對外傷性腦組織損傷及帕金森病等神經退行性疾病動物模型的神經元具有保護作用。2015年6月~2016年6月,我們觀察了免疫抑制劑雷帕霉素對自閉癥大鼠神經、社交行為及海馬區BDNF、凋亡蛋白Bcl-2表達的影響。現報告如下。
1 材料與方法
1.1 材料 成年清潔級Wistar大鼠24只,雌雄各半,雄性體質量為300~350 g、雌性體質量為200~250 g,均購自我院實驗動物中心,許可證號:SYXK(冀)2015-0038。丙戊酸鈉、BDNF、Bcl-2一抗及二抗均購自美國Sigma公司,雷帕霉素購自上海前塵生物科技有限公司。
1.2 動物分組及處理 將Wistar大鼠按雌、雄配對分養過夜,于次日清晨檢查雌鼠有無陰栓,有陰栓者記為妊娠第1天,并將懷孕母鼠單獨飼養。按照隨機數字表法將孕鼠分為對照組、丙戊酸鈉組(模型組)和丙戊酸鈉聯合雷帕霉素組(觀察組),每組4只孕鼠。模型組和觀察組于妊娠12.5天時腹腔注射丙戊酸鈉(600 mg/kg),注射后每天予雷帕霉素4 mg/kg灌胃直至仔鼠斷奶;對照組同期腹腔注射等量生理鹽水。每組取仔鼠20只,均于出生23天時斷奶。
1.3 仔鼠神經行為觀察 在仔鼠出生后35天時,每組選擇6只仔鼠進行神經行為學檢測。被測試仔鼠的出生時間相差在24 h以內,體質量差別在15 g以內,且均為雄性,分籠飼養。測試前1天將仔鼠放入檢測室內適應環境。測試時將仔鼠放入25 cm×25 cm×38 cm封閉箱內中央區(12 cm×12 cm)。對仔鼠行為進行攝像記錄,應用Any-Maze軟件統計仔鼠5 min內在中央區的活動時間和直立次數。
1.4 仔鼠社交行為觀察 仔鼠出生35天時,每組選擇6只仔鼠進行社交行為測試(被測試仔鼠選擇標準同1.3)。測試前1天將仔鼠放入檢測室內適應環境。測試時將仔鼠放入60 cm×60 cm×60 cm透明三室箱的中間小室內適應10 min。在左側小室放入陌生(不來自上述三組)仔鼠,并用鐵絲籠罩住;右側小室放置空的鐵絲籠;打開中間小室進入兩小室的通道。對仔鼠行為進行攝像記錄,應用Any-Maze軟件統計仔鼠10 min內與陌生仔鼠交往時間(社交行為時間)及自梳理時間(非社交行為時間)。
1.5 仔鼠海馬區BDNF、Bcl-2蛋白表達檢測 采用Western blotting法。將出生35天的仔鼠(每組6只,仔鼠選擇標準同1.3)斷頸處死,立即分離左右側海馬,并將其迅速轉移至預先液氮冷卻的勻漿器皿中,將腦組織研磨均勻;滴加含PMSF(60 μL/mg)的蛋白裂解液,冰上裂解30 min;4 ℃ 12 000 r/min離心30 min,取上清進行SDS-PAGE電泳;將電泳分離后的蛋白轉移至PVDF膜上,采用5%脫脂奶粉將轉有蛋白的PVDF膜封閉2 h;分別加入BDNF、Bcl-2一抗,4 ℃過夜;PBS洗滌3次、每次5 min,加入二抗,室溫孵育1 h;PBS洗滌3次、每次10 min,將PVDF膜放入顯影液中顯影,應用成像分析儀對PVDF膜上的蛋白條帶進行成像,檢測各條帶的灰度值。以目的蛋白條帶與內參(β-actin)條帶灰度值的比值表示該樣本蛋白的相對表達量。

2 結果
2.1 各組仔鼠神經行為比較 見表1。
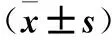
表1 三組仔鼠在中央區活動時間和站立次數比較
注:與對照組比較,*P<0.05;與模型組比較,△P<0.05。
2.2 各組仔鼠社會交往行為比較 見表2。

表2 三組仔鼠社交與非社交行為時間比較
注:與對照組比較,*P<0.05;與模型組比較,△P<0.05。
2.3 各組仔鼠海馬區BDNF、Bcl-2蛋白表達比較 見表3。
3 討論
動物模型是研究自閉癥的主要對象。目前常用的自閉癥動物模型主要有病毒感染模型、Tsc1基因敲除模型和丙戊酸鈉模型等[8]。丙戊酸鈉動物模型在結構效度、表現效度、可信度以及重復性等方面均較好,且能較好地呈現自閉癥的社交行為障礙、交流障礙以及行為重復刻板等臨床特點,是公認的一種較為理想的自閉癥動物模型[9]。本研究結果顯示,模型組仔鼠在中央區活動時間明顯長于對照組仔鼠,站立次數明顯少于對照組仔鼠;模型組仔鼠社會交往能力明顯低于對照組仔鼠,且不愿接觸新事物,不愿與陌生鼠交往;提示丙戊酸鈉制備的自閉癥仔鼠神經行為學表現和社交行為能力均符合自閉癥兒童的行為特點,模型制備成功。

表3 三組仔鼠海馬區BDNF、Bcl-2蛋白相對表達量比較
注:與對照組比較,*P<0.05;與模型組比較,△P<0.05。
雷帕霉素是一種從鏈酶菌中提取的大環內酯類免疫抑制劑,臨床主要應用于真菌感染、腫瘤以及器官移植排斥反應的治療。研究顯示,哺乳動物神經細胞內存在雷帕霉素靶蛋白(mTOR),雷帕霉素與mTOR特異性結合后可抑制mTOR的磷酸化和活性[12],保護神經細胞,減少神經細胞損害[10]。研究提示,mTOR通路與神經元的異常活動可能有關[11]。敲除小鼠模型PTEN基因可激活mTOR通路,導致神經元胞體增大、電活動異常;應用雷帕霉素干預后,神經元電活動明顯改善[13]。此外,雷帕霉素也可抑制TSC基因敲除小鼠星形膠質細胞的過度增殖,并可維持神經元微環境中興奮性谷氨酸鹽的穩定性[14]。本研究結果顯示,給予雷帕霉素干預的仔鼠神經行為和社交能力明顯改善,提示雷帕霉素可能用于預防或治療自閉癥。
BDNF主要由星形膠質細胞和神經元以自分泌和旁分泌的形式分泌,在腦組織的海馬區、扣帶前回、杏仁核以及皮質等區域含量較多[15]。研究顯示,自閉癥患者或動物模型海馬區結構和功能的變化可能與BDNF表達下調有關[16]。海馬區BDNF通過與其受體酪氨酸激酶B(TrkB)結合,導致該受體自磷酸化和包括絲裂原活化蛋白激酶(MAPK)的信號轉導途徑激活,從而促進與學習記憶相關的神經遞質的合成和釋放。另外,BDNF還可以結合位于突觸后膜的TrkB受體,導致NMDA和AMPA谷氨酸受體激活,增加NMDA受體NR1和NR2B單元磷酸化水平以及AMPA受體亞單位GluR1和GluR2/3表達,增強突觸傳遞效能[17]。BDNF表達下調的小鼠海馬長時程增強(LTP)以及海馬依賴性的學習與記憶能力都會受到一定損害[18]。有研究顯示,雷帕霉素可促進過氧化氫(H2O2)及離子霉素聯合去鐵胺損傷作用下的星形膠質細胞存活,對線粒體膜電勢具有保護作用,降低H2O2損傷作用下星形膠質細胞活性氧(ROS)的產生,并可維持細胞內線粒體ROS含量在較低水平[19]。本研究結果顯示,模型組仔鼠海馬區BDNF水平低于對照組,觀察組仔鼠海馬區BDNF水平高于模型組,提示雷帕霉素可上調自閉癥大鼠海馬區BDNF表達水平。
Bcl-2家族是調控細胞凋亡的主要基因家族,可抑制由多種細胞毒素所引起的細胞死亡。自閉癥大鼠海馬區神經細胞凋亡受到抑制,神經細胞增殖、分化和凋亡之間的平衡被打破,進而導致神經元數量增多、體積增大[20]。本研究顯示,模型組仔鼠海馬區Bcl-2蛋白表達明顯高于對照組,觀察組仔鼠海馬區Bcl-2表達明顯低于模型組,提示雷帕霉素可下調海馬區Bcl-2表達,促進海馬區神經元凋亡。雷帕霉素可抑制磷脂酰肌醇3激酶(PI3K)/絲氨酸蘇氨酸蛋白激酶(AKT)/mTOR信號通路。P13K/AKT/mTOR信號通路在體內參與細胞增殖、分化和凋亡等過程,此信號通路激活可抑制細胞凋亡,mTOR活化能夠使Bax表達降低,減弱其對Bcl-2蛋白抗凋亡和促增殖效應的拮抗作用,導致細胞增殖能力增強[21]。雷帕霉素可下調Bcl-2蛋白水平,通過與mTOR結合,抑制Bcl-2活性。
綜上所述,雷帕霉素可改善丙戊酸鈉自閉癥仔鼠模型神經行為表現,提高其社交能力;促進海馬區BDNF表達、抑制Bcl-2表達,調節神經元異常發育、促進細胞凋亡可能是其作用機制。
[1] Napoli E, Dueas N, Giulivi C. Potential therapeutic use of the ketogenic diet in autism spectrum disorders[J]. Front Pediatr, 2014(2):69.
[2] 劉振寰,謝巧玲,林青梅,等.自閉癥兒童的生存質量及影響因素[J].中華實用兒科臨床雜志,2013,28(12):902-904.
[3] Poopal AC, Schroeder LM, Horn PS. Increased expression of the P13K catalytic subunit p110 d underlies elevated S6 phosphorylation and protein synthesis in an individual with autism from a multiplex family[J]. Mol Autism, 2016,7(3):1186-1196.
[4] Yap CC, Winckler B. Acid indigestion in the endosome: linking signaling dysregulation to neurodevelopmental disorders[J]. Neuron, 2013,80(1):4-6.
[5] Rodrigues TM, Jerónimo-Santos A, Sebasti?o AM, et al. Adenosine A(2A) Receptors as novel upstream regulators of BDNF-mediated attenuation of hippocampal Long-Term Depression (LTD)[J]. Neuropharmacology, 2014(79):389-398.
[6] Dutta S, Guhathakurta S, Sinha S, et al. Reelin gene polymorphisms in the Indian population: a possible paternal 5′UTR-CGG-repeat-allele effect on autism[J]. Am J Med Genet B Neuropsychiatr Genet, 2007,144B(1):106-112.
[7] Krajewski S, Krajewska M, Shabaik A, et al. Immunohistochemical determination of in vivo distribution of Bax, a dominant inhibitor of Bcl-2[J]. Am J Pathol, 1994,145(6):1323-1336.
[8] Schneider T, Przewlocki R. Behavioral alterations in rats prenatally exposed to valproic acid: animal model of autism[J]. Neuropsychopharmacology, 2005,30(1):80-89.
[9] Silverman JL, Yang M, Lord C,et al. Behavioral phenotyping assays for mouse models of autism[J]. Nat Rev Neurosci, 2010,11(7):490-502.
[10] Takei N, Nawa H. mTOR signaling and its roles in normal and abnormal brain development [J]. Front Mol Neurosci, 2014(7):28.
[11] Liu F, Zhang H, Zhang K, et al. Rapamycin promotes schwann cell migration and nerve growth factor secretion[J]. Neural Regen Res, 2014,9(6):602-609.
[12] El-Chemaly S, Goldberg HJ, Glanville AR. Should mammalian target of rapamycin inhibitors be stopped in women with lymphangioleiomyomatosis awaiting lung transplantation[J]. Expert Rev Respir Med, 2014,8 (6):657-660.
[13] Ljungberg MC, Sunnen CN, Lugo JN, et al. Rapamycin suppresses seizures and neuronal hypertrophy in a mouse model of cortical dysplasia[J]. Dis Model Mech, 2009,2(7-8):389-398.
[14] Zeng LH, Xu L, Gutmann DH, et al. Rapamycin prevents epilepsy in a mouse model of tuberous sclerosis complex[J]. Ann Neurol, 2008,63(4):444-453.
[15] Zhang JC, Yao W, Hashimoto K. Brain-derived neurotrophic factor(BDNF)-TrkB signaling in inflammation-related depression and potential therapeutic targets[J]. Curr Neuropharmacol, 2015,33(6):668-672.
[16] Gao J, Wang X, Sun H, et al. Neuroprotective effects of docosahexaenoic acid on hippocampal cell death and learning and memory impairments in a valproic acid-induced rat autism model[J]. Int J Dev Neurosci, 2015,26(4):357-361.
[17] Poo MM. Neurotrophins as synaptic modulators[J]. Nat Rev Neurosei, 2001,2(1):24-32.
[18] 尹樂樂,蘇運欽,黃秀艷,等.雷帕霉素對小鼠星形膠質細胞體外誘導凋亡的影響[J].中國病理生理雜志,2015,31(4):652-658.
[19] 晉翔,張雅紅,劉鵬.迷迭香酸對新生大鼠海馬星形膠質細胞BDNF表達的影響及其機制探討[J].山東醫藥,2014,54(12):26-28.
[20] Yu ZW, Ju YH, Yang CL, et al. Antitumor effect of recombinant human endostatin combined with cisplatin on rats with transplanted Lew is lung cancer[J]. Asian Pac J Trop Med, 2015,8(8):664-667.
[21] Junmin S, Hongxiang L, Zhen L, et al. Ginsenoside R93 inhibits coion cancer cell migration by suppressing nuclear factor kappa B activity[J]. J Tradit Chin Med, 2015,35(4):440-404.
10.3969/j.issn.1002-266X.2017.48.012
R729
A
1002-266X(2017)48-0040-03
2017-05-14)
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
作文周刊·小學二年級版(2022年20期)2022-05-05 01:33:06
文萃報·周五版(2021年14期)2021-06-08 22:33:19
中國生殖健康(2020年7期)2021-01-18 03:02:02
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
創新作文(小學版)(2019年10期)2019-09-25 08:12:28
海峽姐妹(2017年5期)2017-06-05 08:53:17
小學生學習指導(低年級)(2017年5期)2017-05-04 04:14:38
丹青少年(2017年2期)2017-02-26 09:11:05
