外源硫化氫對加工番茄種子耐鹽性及抗氧化酶的影響
2017-12-18 06:18:54鄭州元林海榮崔輝梅
干旱地區農業研究 2017年5期
鄭州元,林海榮,崔輝梅
(1.石河子大學農學院,新疆石河子832003;2.特色果蔬栽培生理與種質資源利用兵團重點實驗室,新疆石河子832003)
外源硫化氫對加工番茄種子耐鹽性及抗氧化酶的影響
鄭州元1,2,林海榮1,崔輝梅1,2
(1.石河子大學農學院,新疆石河子832003;2.特色果蔬栽培生理與種質資源利用兵團重點實驗室,新疆石河子832003)
為明確硫化氫對鹽脅迫下加工番茄種子萌發的緩解作用,用NaCl半致死濃度為85 mmol·L-1的KT-63(耐鹽性弱)和半致死濃度為120 mmol·L-1的KT-18(耐鹽性強)為材料,用各自的半致死濃度與不同濃度的NaHS(0.2,0.4,0.6,1.0mmol·L-1)分別配成混合液處理種子。結果表明,隨著 NaHS濃度的增加,種子的發芽率、發芽勢、發芽指數、活力指數、SOD、POD、CAT活性呈先上升后下降的趨勢,其中與鹽脅迫單獨處理相比,用0.4 mmol·L-1NaHS處理,種子的發芽率、發芽勢、發芽指數、活力指數、SOD、POD、CAT活性在 KT-63中分別提升70%、250%、120%、130%、6.7%、150%和180%,在 KT-18中分別提升 50%、250%、60%、80%、4.3%、160%和 200%,而MDA的含量顯著降低,并且對KT-63的緩解效應強于KT-18。綜上所述,H2S(0.4 mmol·L-1)有效緩解了加工番茄萌發過程中鹽脅迫的抑制作用,促進了種子的萌發。
硫化氫;鹽脅迫;加工番茄;種子萌發;抗氧化酶
土壤鹽漬化和次生鹽漬化對農業的威脅已成為一個全球性問題,土壤中過高的鹽分會引起植物生理干旱,植物感受刺激后通過Na+受體作用激活Ca2+通道以及活性氧,如過氧化氫(H2O2)和羥自由基(HO·)在細胞中的積累,影響植物正常營養吸收,阻礙植物正常生長發育,引起植株氧化脅迫,質膜受損,最終引起對植物的傷害甚至死亡[1-2]。在中國絕大部分加工番茄種植在西北地區鹽漬土壤中[3],而新疆又是我國最大的加工番茄種植和加工基地。新疆地處內陸干旱地區,降水稀少蒸發量大,地表積鹽較重,最容易引起土壤鹽漬化[4]。加工番茄是中度耐鹽植物,土壤鹽漬化嚴重影響加工番茄種子萌發率、幼苗和根系的生長發育以及果實的產量和品質。因此,研究與加工番茄抗鹽性及其調控相關的抗氧化酶活性,對指導加工番茄生產中的抗鹽性具有實際意義。
近年來的研究發現H2S是繼NO、CO之后的第三種內源氣體信號分子,參與多種植物的生理過程[5]。已有的結果表明,外源H2S能促進小麥種子在正常情況下和 PEG,Cu2+,Cr,Al3+脅迫下的萌發[6-10],同時外源H2S能促進鹽脅迫下紫花苜蓿種子的萌發[11],以及根的發生[12],氣孔的運動[13]。然而,因為氣候、地域影響,有關H2S對鹽脅迫下新疆加工番茄種子萌發影響以及種子萌發過程中相關抗氧化酶活性及膜質過氧化方面的研究并不清楚。本試驗以新疆兩種加工番茄為材料,研究外源H2S處理對NaCl脅迫下加工番茄種子萌發和相關抗氧化酶(SOD,POD,CAT)活性及丙二醛含量的影響,揭示外源H2S調控NaCl脅迫下加工番茄種子萌發特性及抗氧化酶活性機制,以期為提高加工番茄耐鹽性及揭示其耐鹽性機理提供理論依據。
1 材料與方法
1.1 供試材料
供試番茄材料為:KT-63,KT-18兩個自交品系,由石河子大學農學院實驗室提供;外源硫化氫供體為NaHS,購自sigma公司(中國)。種子實驗在恒溫培養箱內進行,實驗于2015年8月在石河子大學農學院生理實驗室進行。
1.2 試驗設計
挑選大小一致且飽滿的加工番茄種子用37℃~40℃的溫水浸種3 h,之后用10%的高錳酸鉀溶液浸種10min,用蒸餾水沖洗3次。用前期預實驗篩選的NaCl半致死濃度為 85 mmol·L-1耐鹽性弱的KT-63和半致死濃度為 120 mmol·L-1耐鹽性強的KT-18用于實驗,用各自的半致死濃度NaCl和不同濃度NaHS配成混合溶液,試驗處理分別為蒸餾水處理(CK1),85 mmol·L-1NaCl處理(CK2),0.2 mmol·L-1NaHS+85 mmol·L-1NaCl(T1),0.4 mmol·L-1NaHS+85 mmol·L-1NaCl(T2),0.6 mmol·L-1NaHS+85mmol·L-1NaCl(T3),1.0 mmol·L-1NaHS+85mmol·L-1NaCl(T4);蒸餾水處理(CKn),120mmol·L-1NaCl處理(CKp),0.2mmol·L-1NaHS+120mmol·L-1NaCl(M1),0.4 mmol·L-1NaHS+120 mmol·L-1NaCl(M2),0.6 mmol·L-1NaHS+120 mmol·L-1NaCl(M3),1.0mmol·L-1NaHS+120mmol·L-1NaCl(M4)處理。每個處理3個重復,每個重復挑選30粒種子置于鋪有10層濾紙含有不同處理液的培養皿中,6天換一次濾紙。將培養皿置于25℃,16 h/8 h光暗交替的恒溫培養箱中培養12 d,每天觀察種子的萌發情況并記錄發芽數,以胚根長度達到種子自身長度一半為發芽標準。
1.3 測試指標及方法
采用常規方法測定發芽率、發芽勢、發芽指數、活力指數,具體參見孟紅梅等[14]的方法。用直尺測定胚軸和胚根長,用稱重法測定10棵幼苗鮮重。用高俊鳳[15]的硫代巴比妥酸(TBA)比色法測定丙二醛(MDA)含量;用氮藍四唑(NBT)還原法測定超氧化物歧化酶(SOD)活性,以抑制光化還原50%為1個酶活性單位;用愈創木酚法測定過氧化物酶(POD)活性;參照Chance和Maehly[16]的方法測定過氧化氫酶(CAT)活性。
1.4 數據統計分析
數據均取3次重復的平均值,采用Microsoft Excel處理數據,用SPSS 19.0軟件對數據進行單因素方差分析,并運用Duncan’s檢驗法對顯著性差異(P<0.05)進行多重比較,結果以平均值 ±標準誤(means±SD)表示,用 Microsoft Excel作圖。
2 結果與分析
2.1 外源H2S對NaCl脅迫下加工番茄種子發芽的影響
不同濃度的NaHS處理對NaCl脅迫下兩種番茄種子的萌發有一定的影響。隨著NaHS濃度的增加,兩種加工番茄品系的發芽相關指標呈現先上升后下降的趨勢(表 1)。用 0.2~1.0 mmol·L-1NaHS+NaCl處理較NaCl單獨處理兩個品系發芽率顯著增加,在 0.4~1.0mmol·L-1時達到與空白對照相近的水平,都表現為差異不顯著,并且兩個品系都在0.4mmol·L-1時發芽率、發芽勢、發芽指數和活力指數在脅迫條件下達到最高,在KT-63中,NaHS濃度為 0.4mmol·L-1時發芽率、發芽勢、發芽指數和活力指數較 CK2分別提升 0.7倍、2.5倍、1.2倍和 1.3倍,差異顯著。在KT-18中,NaHS濃度為0.4mmol·L-1時發芽率、發芽勢、發芽指數和活力指數較CKp分別提升 0.48倍、2.5倍、0.6倍和 0.8倍,其中發芽率、發芽指數和活力指數都達到顯著水平。結果表明 0.4~1.0 mmol·L-1NaHS都能緩解 NaCl脅迫對番茄種子萌發的抑制作用,其中以0.4 mmol·L-1效果最好,同時對發芽指標的緩解效果看,0.4 mmol·L-1的NaHS對耐鹽性弱的KT-63緩解效應更強。
2.2 外源H2S處理對NaCl脅迫下發芽加工番茄種子生長的影響
由圖1可以看到NaCl脅迫顯著降低了KT-63和KT-18的胚軸長,在添加不同濃度的NaHS處理之后,隨著NaHS濃度增加胚軸長度都呈現由上升到下降的趨勢。在 KT-63中,T2-T4條件下較CK2相比胚軸長度都達到了顯著水平,其中NaHS濃度為 0.4mmol·L-1胚軸最長,較 CK2相比提升了12.2%。在KT-18中,M1-M3比起 CKp胚軸長度顯著增加,在 NaHS濃度為 0.4 mmol·L-1時達到最大,較 CKp增加了 6.0%,其中 M4比 CKp顯著降低。說明低濃度的NaHS能促進鹽脅迫下加工番茄萌發期幼苗胚軸的伸長,在耐鹽性不同的兩個品系中,NaHS濃度為 0.4 mmol·L-1時處理效果最好,對耐鹽性強的 KT-18而言,1.0mmol·L-1的濃度抑制了胚軸伸長。
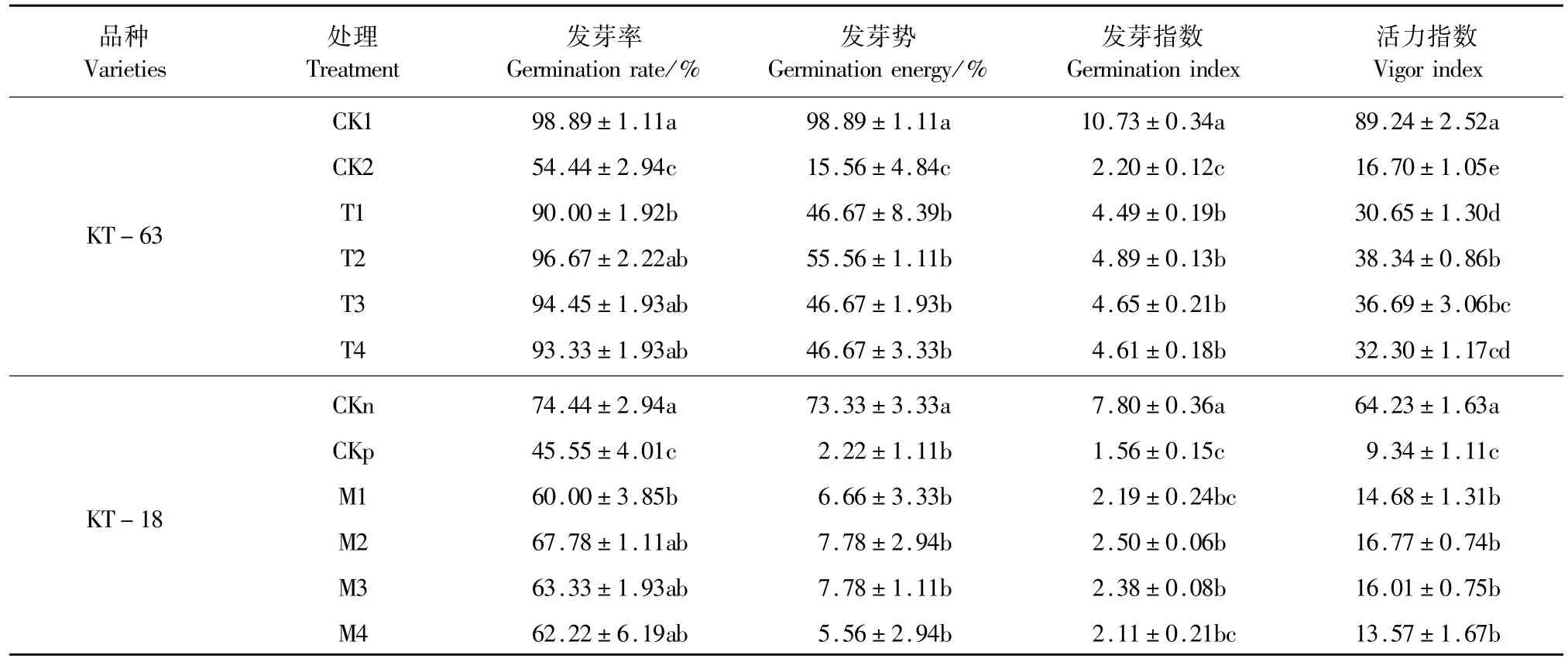
表1 NaHS對NaCl脅迫下加工番茄種子萌發的影響Table 1 Effect of exogenous NaHSon seed germination of processing tomato under NaCl stress
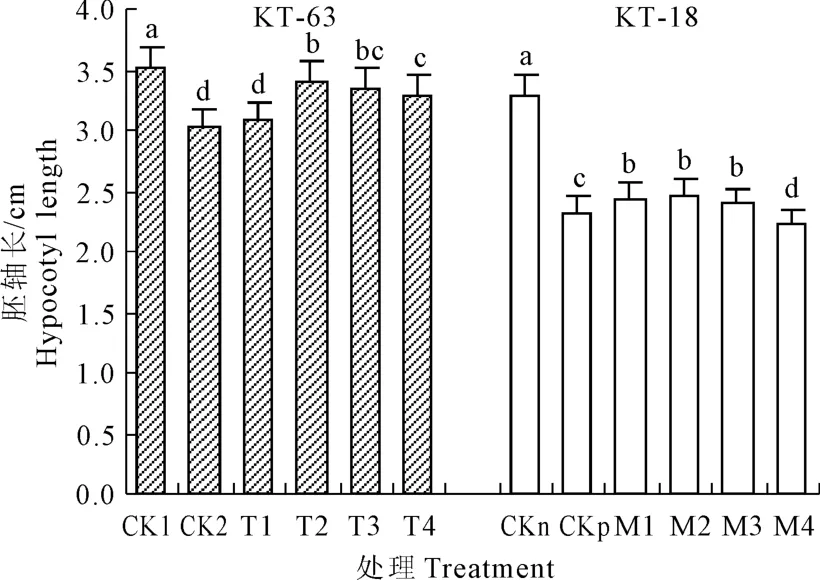
圖1 NaHS對NaCl脅迫下番茄幼苗胚軸長的影響Fig.1 Effectof exogenous NaHSon hypocotyl length of tomato seedlings under NaCl stress
不同濃度的NaHS處理對NaCl脅迫下加工番茄的胚根長有一定影響。圖2表明,與空白對照相比,NaCl脅迫使兩種番茄胚根長都顯著降低。隨著NaHS處理濃度的增加總體上胚根長呈現先升后降的趨勢,在KT-63中,T1-T3較CK2處理有明顯的促進作用,當濃度為 1.0 mmol·L-1時迅速下降。在KT-18中,NaHS處理也促進了鹽脅迫下胚根的伸長,但沒有達到顯著水平,在M3-M4時緩慢下降。當 NaHS濃度為 1.0mmol·L-1時較 0.6 mmol·L-1時胚根長在KT-63和KT-18中分別下降11.4%和3.0%。這個結果說明低濃度的NaHS可以促進鹽脅迫下加工番茄萌發期胚根的伸長,緩解鹽脅迫對胚根伸長的抑制作用,當濃度為 1.0mmol·L-1時,緩解效應下降,對于耐鹽性強的KT-18,NaHS的效應隨濃度的變化較緩慢。
圖3表明,T2-T3和M1-M3處理與鹽脅迫單獨處理相比,分別顯著增加了KT-63和KT-18在鹽脅迫下的側根數目,但在T4和M4時側根數目又迅速減少了,KT-63和KT-18兩品系側根數目都在 NaHS濃度為 0.4mmol·L-1時達到了最多。說明NaHS可以促進鹽脅迫下側根的形成,但高濃度的NaHS可能還會抑制側根的形成。

圖2 NaHS對NaCl脅迫下番茄幼苗胚根長的影響Fig.2 Effect of exogenous NaHSon root length of tomato seedlings under NaCl stress
圖4 表明,在T2-T4和M2-M4分別能顯著增加KT-63和KT-18在鹽脅迫下胚軸鮮重,并且NaHS濃度為 0.4 mmol·L-1時兩種在鹽脅迫下的加工番茄胚軸鮮重達到最重,分別較CK2和CKp增加了20.7%和23.5%。在KT-18中,M4胚軸鮮重與CKp相比有所下降。說明因耐鹽性不同,NaHS對兩個品系胚軸鮮重的影響程度也不一樣,對于KT-18來說高濃度的NaHS沒有緩解作用還加劇了脅迫。

圖3 NaHS對NaCl脅迫下番茄幼苗側根數的影響Fig.3 Effectof exogenous NaHSon lateral root number of tomato seedlings under NaCl stress
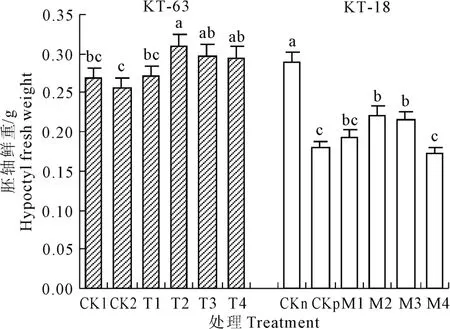
圖4 NaHS對NaCl脅迫下番茄幼苗胚軸鮮重的影響Fig.4 Effect of exogenous NaHSon hypocotyl fresh weightof tomato seedlings under NaCl stress
圖5 表示,T1-T4處理顯著增加了KT-63在鹽脅迫下胚根的鮮重,但是在KT-18中NaHS處理使鹽脅迫下胚根鮮重有所增加卻沒有達到顯著水平,兩個加工番茄品系都是在 NaHS濃度為 0.4 mmol·L-1時對胚根鮮重的作用效果最好,分別比CK2和CKp增加了40.9%和13.3%。以上結果表明,NaHS緩解鹽脅迫抑制胚軸鮮重增加的效應在耐鹽性弱的品系中更加明顯,表現為對鹽敏感對緩解物質NaHS的作用效果也敏感。

圖5 NaHS對NaCl脅迫下番茄幼苗胚根鮮重的影響Fig.5 Effect of exogenous NaHSon root fresh weight of tomato seedlings under NaCl stress
2.3 外源H2S處理對NaCl脅迫下加工番茄種子發芽抗氧化酶活性的影響
圖6表明,兩個品系加工番茄種子在NaCl脅迫(CK2,CKp)下較空白對照(CK1,CKn)相比 SOD活性顯著下降。而外源NaHS處理有效緩解了NaCl對番茄種子發芽過程中SOD活性的降低,鹽脅迫下隨著NaHS處理濃度的增加,SOD活性呈現先上升后下降的趨勢。在鹽脅迫下兩個品系中SOD活性都在NaHS濃度為 0.4mmol·L-1時達到了最高,SOD活性分別比CK2和CKp提高了6.7%和4.3%。以上結果說明,在這兩個品系中 0.4 mmol·L-1的 NaHS濃度為緩解NaCl脅迫下SOD活性降低的最適宜濃度,并且NaHS對耐鹽性弱的KT-63緩解效應表現的更強。
由圖7可以看出,在兩個番茄品系中鹽脅迫顯著降低了萌發過程中幼苗中POD的活性。加入外源NaHS后,隨NaHS濃度的增加,POD活性總體呈先升后降的趨勢。在 KT-63中,T2與 CK2相比POD活性顯著增加,活性提升了1.5倍,而 T4時POD活性比 CK2還要低。在 KT-18中,從 M1-M4,與CKp相比POD活性都顯著增加,并且在M2時達到最高,比CKp增加1.6倍。以上結果表明,NaHS對幼苗NaCl脅迫下POD活性的影響效果與濃度和品系有關,耐鹽性弱的KT-63對不同濃度的NaHS也較為敏感,高濃度的NaHS還會進一步降低POD的活性,而對于耐鹽性較強的KT-18,POD活性隨NaHS濃度增加變化相對較為緩和,但這兩個品系中最佳 NaHS作用濃度都是 0.4mmol·L-1。

圖6 NaHS對NaCl脅迫下番茄幼苗SOD活性影響Fig.6 Effect of exogenous NaHS on SOD activity of tomato seedlings under NaCl stress
如圖8所示,在各處理中,CAT活性表現出與POD相似的變化趨勢。在 KT-63中,T1-T3較CK2的 CAT活性都顯著增加,并且在 T2和 T3時CAT活性比CK1還要高,都達到顯著水平,分別是CK2的1.77倍和 1.78倍。在 KT-18中,M2-M4水平下CAT活性與CKp相比顯著升高,并且在M2時CAT活性最高,是CKp的2倍。結果表明,在加入NaHS后,對CAT活性影響效應在兩個品系中不完全一樣,在KT-63中NaHS處理后較空白對照相比CAT活性顯著提升,但在KT-18中沒有這樣明顯的效應。說明NaHS效應在這兩個抗鹽性不同品系間有差異,表現為鹽脅迫下KT-63對NaHS的作用效果更敏感。

圖7 NaHS對NaCl脅迫下番茄幼苗POD活性影響Fig.7 Effectof exogenous NaHSon POD activity of tomato seedlings under NaCl stress

圖8 NaHS對NaCl脅迫下番茄幼苗CAT活性影響Fig.8 Effect of exogenous NaHS on CAT activity of tomato seedlings under NaCl stress
2.4 外源H2S處理對NaCl脅迫下加工番茄種子發芽丙二醛含量的影響
由圖9可知,CK2與CK1相比,CKp與CKn相比丙二醛(MDA)含量均顯著升高。添加NaHS后,隨著NaHS濃度的升高MDA含量在兩個番茄品系中都表現先降低后升高的趨勢。在KT-63中,從T1-T4的MDA含量都顯著比CK2低,并且在T3降到最低。在KT-18中,NaHS處理組中MDA含量較CKp也顯著降低,在 M2降到了最低。這說明用NaHS處理鹽脅迫下加工番茄可以緩解萌發期由鹽脅迫所引起的MDA的積累。在不同濃度NaHS處理中,0.6mmol·L-1和 0.4mmol·L-1NaHS可以使鹽脅迫下KT-63和KT-18中MDA含量分別降到最低,比CK2和CKp分別下降36.0%和26.0%。說明NaHS可以顯著緩解萌發期兩種耐鹽性不同番茄幼苗的膜脂過氧化程度。
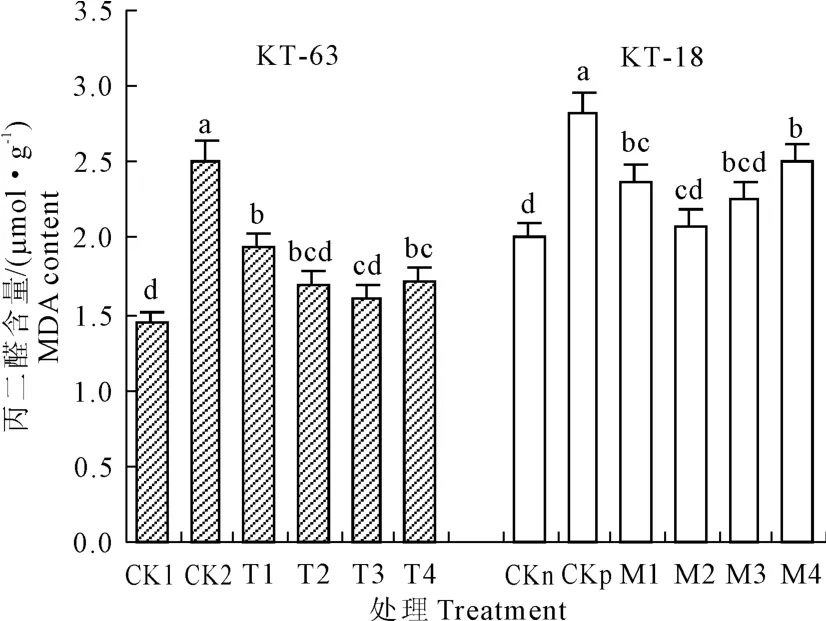
圖9 NaHS對NaCl脅迫下番茄幼苗MDA含量影響Fig.9 Effectof exogenous NaHSon MDA content of tomato seedlings under NaCl stress
3 討論與結論
種子萌發是植物生長發育的重要階段,但是近年來的研究結果表明鹽脅迫對種子的萌發具有顯著的抑制作用。由于高濃度的鹽脅迫使水勢降低,造成種子吸水困難,同時還會破壞細胞質膜的完整性,進而影響芽和根的伸長。在植物生育期中,種子萌發期是對鹽脅迫較為敏感的時期,利用外源物質提高種子萌發期的抗鹽性是很重要的方法。
外源物質H2S被報道參與植物的生長發育以及脅迫耐性的獲得[17]。NaHS處理能改善幼苗的生長狀況,可以提高能源植物小桐子種子的發芽率和發芽勢[18]。本實驗中H2S供體NaHS可以提高鹽脅迫下兩種耐鹽性不同加工番茄種子發芽指標,說明H2S能緩解鹽脅迫對加工番茄種子萌發的抑制,這個結果與Wang等[11]H2S可以緩解鹽脅迫對紫花苜蓿種子萌發的抑制相一致。NaHS處理對鹽脅迫下萌發期幼苗胚軸的影響,說明NaHS能促進鹽脅迫下加工番茄萌發期幼苗胚軸和胚根的伸長,胚軸和胚根鮮重的增加,還能促進側根的形成,這個結果與Li等[19]的一致。不同濃度的 NaHS處理對 NaCl脅迫下加工番茄種子發芽和生長的影響說明低濃度的NaHS能促進鹽脅迫下加工番茄種子的萌發,而高濃度的NaHS對脅迫的緩解作用降低,甚至加重脅迫作用,這種表現在耐鹽性弱的KT-63中更為明顯。這可能與高濃度的NaHS產生的滲透脅迫有關,同時對于環境變化感受較為明顯的品系來說,是不是因為品系間基因型存在差異進而對環境的感受也存在差異,有待進一步研究。
SOD,POD,CAT在清除超氧自由基、過氧化物、過氧化氫等方面具有重要作用[20]。在本實驗中NaHS處理使鹽脅迫下SOD,POD,CAT活性在兩種番茄中都有所增加,這個結果與何慶元[21]在大豆中的結論相似。在正常環境下植物體內活性氧的產生與清除是處于動態平衡的,當植物遭受逆境脅迫時,這種動態平衡就會被打破,使 SOD、POD、CAT活性發生變化[22]。本實驗中耐鹽性弱的 KT-63對NaHS的作用效果也相對敏感,而耐鹽性強的KT-18對NaHS濃度的變化表現遲鈍。說明NaHS對番茄萌發過程中鹽脅迫產生的氧化損傷具有一定的緩解效應,番茄幼苗鹽脅迫的緩解作用可能與品系有關,同時這三種抗氧化酶在耐鹽性不同的兩個品系中的升降幅度不同,可能是NaHS對不同品系的作用效應或對這三種酶活性的調節存在差異,這有待于進一步研究。
MDA含量是衡量膜質過氧化的一個重要指標,其含量的多少與植物遭受的氧化脅迫成正比[23]。本實驗中鹽脅迫顯著增加了兩種加工番茄萌發過程MDA的含量,這個結果與劉文瑜等[24]的研究結果一致。用不同濃度的NaHS處理后MDA含量發生不同程度的降低。說明NaHS能降低番茄種子在鹽脅迫下萌發過程中MDA的積累,能夠通過緩解質膜受損程度來促進幼苗的生長,但是在兩品系中MDA含量最低時的NaHS濃度不同,可能是不同濃度NaHS處理對MDA含量的調節在品系間存在差異。
綜上,本文中外源H2S對鹽脅迫下兩個耐鹽性不同加工番茄品系種子耐鹽性及抗氧化酶影響的研究表明,外源NaHS能不同程度增加兩種加工番茄品系的發芽相關指標,當 NaHS濃度為0.4 mmol·L-1時種子發芽率、發芽勢、發芽指數、活力指數、胚軸長、胚根長、側根數、胚軸鮮重、根鮮重在鹽脅迫下都達到了最大。同樣的,低濃度的NaHS增強了種子萌發過程中的抗氧化酶活性,其中NaHS濃度為0.4mmol·L-1時 SOD、POD、CAT活性在鹽脅迫下最強。種子在鹽脅迫萌發過程中MDA含量在KT-63和 KT-18中分別在 NaHS濃度為 0.6 mmol·L-1和0.4mmol·L-1時降到了最低。整體表現為低濃度的NaHS緩解了鹽脅迫下加工番茄種子的萌發,增強了抗氧化酶的活性,降低了丙二醛含量的積累,并且NaHS對耐鹽性弱的KT-63緩解效應表現得更強。
[1] Deinlein U,Stephan A B,Horie T,etal.Plant salt-tolerancemechanisms[J].Trends in plant Science,2014,19(6):371-379.
[2] 王 聰,朱月林,楊立飛,等.NaCl脅迫對菜用大豆種子膨大過程中抗氧化系統及滲透調節物質的影響[J].西北植物學報,2012,32(2):297-305.
[3] Mao R,Fitzpatrick R,Liu X,et al.Chemical properties of selected soils from the North China Plain[C]//Mcvicar TR,Li R,Walker J,etal.Regionalwater and soil assessment formanaging sustainable agriculture in China and Australia.Canberra:Australian Centre for International Agricultural Research,2002.
[4] 曾幼玲,蔡忠貞,馬 紀,等.鹽分和水分脅迫對兩種鹽生植物鹽爪爪和鹽穗木種子萌發的影響[J].生態學雜志,2006,25(9):1014-1018.
[5] WANG R.Physiological implications of hydrogen sulfide:a whiff exploration that blossomed[J].Physiol Rev,2012,92(2):791-896.
[6] Zhang H,Hu L Y,Hu K D,et al.Hydrogen sulfide promoteswheat seed germination and allevi-ates oxidative damage against copper stress[J].Integr Plant Biol,2008,50(12):1518-1529.
[7] Zhang H,Hu LY,Li P,et al.Hydrogen sulfide alleviated chromium toxicity in wheat[J].Biol Plant,2010,54(4):743-747.
[8] Zhang H,Jiao H,Jiang CX,etal.Hydrogen sulfide protects soybean seedlings against drought-induced oxidative stress[J].Acta Physiol Plant,2010,32(5):849-857.
[9] Zhang H,Tan ZQ,Hu L Y,et al.Hydrogen sulfide alleviates aluminum toxicity in germinating wheat seedlings[J].Integr Plant Biol,2010,52(6):556-567.
[10] Zhang H,Wang M F,Hua L Y,et al.Hydrogen sulfide promotes wheat seed germination under osmotic stress[J].Russ JPlant Physiol,2010,57(4):532-539.
[11] Wang YQ,Li L,CuiW T,etal.Hydrogen sulfide enhancesalfalfa(Medicago sativa)tolerance against salinity during seed germination by nitric oxide pathway[J].Plant Soil,2012,351(1-2):107-119.
[12] Zhang H,Tang J,Liu X P,et al.Hydrogen sulfide promotes root organogenesis in Ipromoea batatas,Salixmatsudana and Glycinemax[J].Integr Plant Biol,2009,51(12):1084-1092.
[13] 王蘭香,侯智慧,侯麗霞,等.H2O2介導的H2S產生參與干旱誘導的擬南芥氣孔關閉[J].植物學報,2012,47(3):217-225.
[14] 孟紅梅,韓多紅,李彩霞,等.NaCl脅迫對板藍根種子萌發的影響[J].干旱地區農業研究,2008,26(1):213-216.
[15] 高俊鳳.植物生理學實驗指導[M].北京:高等教育出版社,2006.
[16] Chance B,Maehly A C.Assay of catalase and peroxidase[J].Methods in Enzymology,1995,764(2):775.
[17] Lisjak M,Teklic T,Wilson ID,et al.Hydrogen sulfide:Environmental factor or signalingmolecule[J].Plant cell Environ,2013,36(9):1607-1616.
[18] Li ZG,Gong M,Liu P.Hydrogen sulfide isamediator in H2O2-induced seed germination in Jatropha curcas[J].Acta Physiol Plant,2012,34(6):2207-2213.
[19] Li Y J,Chen J,Xian M,etal.In Site Bioimaging ofHydrogen Sulfide Uncovers Its Pivotal Role in Regulating Nitric Oxide-Induced Lateral Root Formation[J].PLoSONE,2014,9(2):e90340.doi:10.1371/journal.pone.0090340.
[20] Bowler C,Van M,Inzc D.Superoxide dismutaseand stress tolerance[J].Annual Review plant Physiology and Plant Molecular Biology,1992,(43):83-116.
[21] 何慶元,向仕華,吳 萍,等.硫化氫對鹽脅迫條件下大豆抗氧化酶活性的影響[J].大豆科學,2015,34(3):427-431.
[22] 趙可夫,盧元芳,張寶澤,等.Ca2+離子對小麥幼苗降低鹽害效應的研究[J].植物學報,1993,35(1):51-56.
[23] 李合生.植物生理生化實驗原理和技術[M].北京:高等教育出版社,2000:267-268.
[24] 劉文瑜,楊宏偉,魏小紅,等.外源NO調控鹽脅迫下蒺藜苜蓿種子萌發生理特性及抗氧化酶的研究[J].草業學報,2015,24(2):85-95.
Effects of exogenous hydrogen sulfide on salt tolerance and antioxidant enzymes of processing tomato seeds
ZHENG Zhou-yuan1,2,LIN Hai-rong1,CUIHui-mei1,2
(1.College of Agriculture,ShiheziUniversity,Shihezi,Xinjiɑng 832003,Chinɑ;2.Xinjiɑng Productionɑnd Construction Corps Key Lɑborɑtory of Speciɑl Fruitsɑnd Vegetɑbles Cultivɑtion Physiologyɑnd Germplɑsm Resources Utilizɑtion,Shihezi,Xinjiɑng 832003,Chinɑ)
The effects of exogenous hydrogen sulfide on the seed germination of processing tomato under salt stress were investigated.Seeds of two processing tomato cultivars KT-63(salt resistance isweak)and KT-18(stronger salt tolerance)were treatmentwith semi-lethal concentration of NaCl(85mmol·L-1,120mmol·L-1)and NaHS(0.2,0.4,0.6,1.0mmol·L-1)mixture,respectively.The results showed that:With increasing NaHS level,the germination rate,germination energy,germination index,vigor index,SOD,POD and CAT activities first increased and then decreased.Compared with salt stress,treatmentswith 0.4mmol·L-1NaHS significantly increased seed germination rate,germination index,vigor index and the activities of SOD,POD and CAT in KT-63(70%,250%,120%,130%,6.7%,150%and 180%,respectively)and KT-18(50%,250%,60%,80%,4.3%,160%and 200%,respectively),and the content of MDA was decreased.Themitigation effect of hydrogen sulfide on KT-63 is stronger than KT-18.Results provide evidence that0.4mmol·L-1of H2Smay alleviate damage to seeds caused by salt stress,expedition of seed germination.
hydrogen sulfide;salt stress;processing tomato;seed germination;antioxidase
S641.2;S603.4
A
1000-7601(2017)05-0236-06
10.7606/j.issn.1000-7601.2017.05.35
2016-08-05
2016-12-13
留學回國人員科技活動項目(2010LX005)
鄭州元(1989—),男,甘肅武威人,碩士研究生,研究方向為蔬菜遺傳育種。E-mail:1294450645@qq.com。
崔輝梅(1971—),女,新疆石河子人,教授,主要從事蔬菜種質資源與遺傳育種研究。E-mail:chm agr@shzu.edu.cn。
