小麥品種東農(nóng)冬麥2號根中 TaEXPA7部分同源基因的克隆及表達特性分析
2017-12-11 08:50:56董佳敏徐永清彭麗娜姚樹寬趙巧芩李鳳蘭胡寶忠
麥類作物學報 2017年11期
董佳敏,徐永清,彭麗娜,馮 旭,姚樹寬,趙巧芩,李鳳蘭,胡寶忠,2
(1.東北農(nóng)業(yè)大學生命科學學院,黑龍江哈爾濱 150030; 2.哈爾濱學院,黑龍江哈爾濱 150086)
小麥品種東農(nóng)冬麥2號根中 TaEXPA7部分同源基因的克隆及表達特性分析
董佳敏1,徐永清1,彭麗娜1,馮 旭1,姚樹寬1,趙巧芩1,李鳳蘭1,胡寶忠1,2
(1.東北農(nóng)業(yè)大學生命科學學院,黑龍江哈爾濱 150030; 2.哈爾濱學院,黑龍江哈爾濱 150086)
膨脹素(expansin)是植物生長發(fā)育過程中誘導(dǎo)細胞壁松弛和伸展的蛋白,包括EXPA、EXPB、EXLA、EXLB四個基因家族,對植物根系的形成及快速發(fā)育具有重要作用。本試驗以寒地冬小麥品種東農(nóng)冬麥2號兩葉一心期的根為材料,克隆了 TaEXPA7基因的3個CDS全長,其氨基酸序列同源性較高,僅信號肽處有兩個氨基酸不同,其核酸序列長度均為777 bp,編碼258個氨基酸,分別命名為 TaEXPA7-A、 TaEXPA7-B、 TaEXPA7-D,且分別定位于2AL、2BL、2DL染色體。蛋白均含有DPBB-1和Pollen-allerg-1兩個保守結(jié)構(gòu)域,分子量為27 670.74 Da,等電點為8.09,均為疏水性蛋白;與節(jié)節(jié)麥、二穗短柄草的相似性分別為99%和92%。采用兩葉一心期的根進行qRT-PCR分析發(fā)現(xiàn), TaEXPA7-A/B/D三個基因在根尖中的表達量均較高,伸長區(qū)和成熟區(qū)次之;對冬小麥根進行低溫、干旱和激素處理后,這三個基因均下調(diào)表達,并且 TaEXPA7-B基因的相對表達量較高, TaEXPA7-D的相對表達量較低。由此可見,多倍體小麥不同染色體上的基因在應(yīng)對不同環(huán)境脅迫時,表達以及應(yīng)答模式存在差異,并且在不同的環(huán)境脅迫下,發(fā)揮主導(dǎo)作用的染色體也存在差異;此外,該基因在根中的表達可能與低溫、干旱以及外源激素的脅迫有一定的關(guān)系,可能是促進根系生長的重要基因。
冬小麥; TaEXPA7;部分同源;表達特性
膨脹素(expansin)是植物生長發(fā)育過程中誘導(dǎo)細胞壁松馳和伸展的蛋白,于1989年首次被發(fā)現(xiàn)。1992年,McQueen-Mason等[1]從黃瓜中分離純化出這種蛋白,并發(fā)現(xiàn)其能調(diào)節(jié)幼苗下胚軸的伸長,第一次將其命名為expansin。在植物中,expansin可以分為四個基因家族α-expansin(EXPA)、β-expansin(EXPB)、expansin-likeA(EXLA)以及expansin-likeB(EXLB),由信號肽、催化區(qū)和結(jié)合區(qū)三部分構(gòu)成[2]。
Hu等[3]分別克隆了小麥 TaEXPA1-A/B/D三個基因,并將其轉(zhuǎn)入擬南芥,發(fā)現(xiàn)其具有相似的功能。然而,小麥染色體組的復(fù)雜多倍體特性使得對其遺傳、表達特性及其功能的分析極具挑戰(zhàn)性。研究發(fā)現(xiàn), OsEXPA1、 OsEXPA2、 OsEXPA3、 OsEXPA4、OsEXPB2和 OsEXPA17等基因均在水稻根尖特異性表達,參與根尖形態(tài)建成,促進根系生長發(fā)育[4-7]。近年來,國內(nèi)外關(guān)于膨脹素基因在各種非生物脅迫下的表達特性被廣泛研究,如干旱、低溫和激素會影響膨脹素基因的表達,從而影響植物生長。Che等[8]研究發(fā)現(xiàn),Al離子誘導(dǎo)后, OsEXPA10基因參與水稻的根細胞伸長。韓陽陽等[9]研究發(fā)現(xiàn),GA3、ET和IAA顯著抑制了 TaEXPB23的表達水平,而MeJA則上調(diào)了該基因的表達,IAA處理先抑制隨后誘導(dǎo)了 TaEXPB23的表達,而ABA則在后期稍微抑制了 TaEXPB23的表達;此外,鹽脅迫能夠明顯提高 TaEXPB23的mRNA表達水平,而高溫則抑制該基因的表達。
目前,在小麥中關(guān)于膨脹素和根之間關(guān)系的研究鮮有報道,復(fù)雜的小麥各個基因組之間的關(guān)系以及基因的表達差異關(guān)系也尚未明確。因此,本研究以寒地高產(chǎn)冬小麥品種東農(nóng)冬麥2號為材料,同源克隆了 TaEXPA7-A/B/D三個基因;通過生物信息學分析了解該基因基本的生物學功能;利用qRT-PCR技術(shù),研究了這三個基因在小麥幼苗期根中不同部位及在低溫、干旱和激素脅迫下的表達特性及表達差異,以期為進一步研究膨脹素在根系形態(tài)建成中的作用提供理論依據(jù)。
1 材料與方法
1.1 試驗材料及處理
試驗材料為高抗寒的冬小麥品種東農(nóng)冬麥2號,由東北農(nóng)業(yè)大學農(nóng)學院小麥育種研究室提供。
挑選顆粒飽滿的東農(nóng)冬麥2號種子,用0.1% HgCl2溶液消毒處理30 s,蒸餾水沖洗6 h,室溫浸種24 h,然后將種子平鋪于含有雙層濾紙的培養(yǎng)皿中培養(yǎng),露白后移栽至苗缽,置于培養(yǎng)室中培養(yǎng)[9]。待根長至大約30 cm時對根尖、伸長區(qū)和成熟區(qū)進行取樣,每個部位均單株取材,隨機取三株。以水培為對照組,對4 ℃、聚乙二醇(PEG)、脫落酸(ABA)、茉莉酸甲脂(MeJA)及水楊酸(SA)分別處理2 h、6 h、12 h、24 h和48 h的根進行取材,均單株取材,隨機取三株,取樣后均迅速放入液氮中速凍,然后置于-80 ℃超低溫冰箱保存?zhèn)溆谩?/p>
RNA提取試劑盒TransZol、RT-PCR所用熒光染料SYBR Green、Top Taq酶、 Peasy-T3克隆載體、質(zhì)粒提取試劑盒、DNA回收試劑盒、反轉(zhuǎn)錄試劑盒均購自全式金生物科技有限公司;試驗所有引物合成和測序在哈爾濱博仕生物進行;試驗所需大腸桿菌 DH5α、ABA、茉莉酸甲酯、水楊酸、聚乙二醇、LB肉湯、LB營養(yǎng)瓊脂、氯仿、甘油等生化試劑均購自凱譽生物科技有限公司。使用Primer 5.0軟件進行引物設(shè)計;使用Excel和GraphPad Prism軟件完成作圖和數(shù)據(jù)處理。
1.2 RNA的提取及cDNA的合成
使用TransZol試劑盒(ER501)進行總RNA的提取,-80 ℃保存?zhèn)溆谩J褂肬V-240紫外分光光度計檢測RNA濃度,OD260/OD280=1.8~2.1,且OD260/OD230>2.0,濃度不小于500 ng·μL-1,使用瓊脂糖凝膠電泳檢測總RNA的完整性。以O(shè)ligo(DT)為引物,將所得RNA反轉(zhuǎn)錄為cDNA,-20 ℃保存,并用β-actin檢測反轉(zhuǎn)錄效果。
1.3 TaEXPA7-A/B/D基因的克隆及序列分析
組織特異性表達分析表明, TaEXPA7基因在根中的相對表達量較高,且東農(nóng)冬麥2號的RNA-seq結(jié)果顯示,該基因的整體表達量較高,可能與根系生長有重要關(guān)系。在NCBI的GenBank中查詢 TaEXPA7三個同源基因的CDS序列,對這三個基因的相似部分利用Primer 5.0設(shè)計特異性引物(表1)。以冬小麥幼苗期根的cDNA為模板進行PCR擴增,電泳回收后,將回收產(chǎn)物連接到Peasy-T3載體上,轉(zhuǎn)入DH5α感受態(tài)細胞,用含Amp(100 μg·mL-1)的LB平板篩選目的菌落,隨機挑取30個單菌落,采取菌液PCR的方法進行目的基因擴增,將出現(xiàn)目標基因的克隆送公司測序。利用NCBI ORF Find分析CDS序列的ORF,并用Primer 5.0軟件預(yù)測編碼的氨基酸序列,對獲得的序列用BLAST程序搜索GenBank中與 TaEXPA7-A/B/D基因同源的氨基酸序列,采用DNAMAN 6.0和MEGA 6.0軟件進行氨基酸序列比對和系統(tǒng)進化樹的構(gòu)建。
1.4 TaEXPA7-A/B/D基因的表達特性分析
在NCBI的GenBank中查詢 TaEXPA7-A/B/D的CDS差異序列并參照此差異序列分別設(shè)計特異性qRT-PCR引物(表1)。對1.2中得到的各處理cDNA進行梯度稀釋,根據(jù)Ct值確定最佳反應(yīng)濃度。以確定好最佳濃度的cDNA為模板,以小麥肌動蛋白基因β-actin為內(nèi)參,采用qRT-PCR進行擴增、分析,生物學重復(fù)和技術(shù)重復(fù)各3次。反應(yīng)體系(20 μL)如下:2×Trans Star Top Green Qpcr Super Mix 10 μL,上下游引物各0.4 μL,cDNA 1.0 μL,ddH2O 8.2 μL。反應(yīng)在實時熒光定量PCR儀(FQD-9620)上進行。采用2-ΔΔCt法計算相對表達量,采用DPS 7.05 數(shù)據(jù)處理系統(tǒng)軟件進行方差及顯著性分析。

表1 引物信息Table 1 Information of primers used in this study
2 結(jié)果與分析
2.1 RNA的檢測結(jié)果
以電壓150 V,電流120 mA,1%的瓊脂糖凝膠電泳檢測所提取的RNA(圖1),可以看出,28S和18S兩條譜帶清晰且前者的亮度大于后者,說明提取的總RNA質(zhì)量較好。
2.2 TaEXPA7-A/B/D基因CDS的克隆及序列分析
為了獲得 TaEXPA7的部分同源基因,根據(jù) TaEXPA7-A/B/D三個基因的ORF(Open Reading Frame,開放閱讀框)序列(GenBank登錄號:AA0307290.1、AA0383180.1、AA0559500.1)的相似部分,設(shè)計了兩條特異引物( TaEXPA7-F,TaEXPA7-R),進行了PCR,經(jīng)瓊脂糖凝膠電泳后獲得了一條800 bp左右的條帶(圖2)。將攜帶該目的條帶的克隆挑取30個進行測序,獲得了3條分別定位于2AL、2BL和2DL染色體上的CDS序列,其長度均為777 bp,依次命名為: TaEXPA7-A、TaEXPA7-B、TaEXPA7-D。生物信息學分析發(fā)現(xiàn),3個基因均編碼258個氨基酸,分子量為27 670.74 Da,等電點為8.09,均為疏水性蛋白,包括DPBB-1和Pollen-allerg-1兩個保守結(jié)構(gòu)域,其三級結(jié)構(gòu)沒有明顯差異(圖3)。利用DNAMAN進行氨基酸同源比對結(jié)果(圖4)表明,3條序列僅在信號肽處有兩個氨基酸的編碼存在差異。而冬小麥與春小麥中 TaEXPA7-A/B/D僅在核苷酸序列中存在堿基差異,蛋白序列不存在差異。眾所周知,一般的蛋白質(zhì)都通過保守的結(jié)構(gòu)域行使其功能,而本試驗中的這3條氨基酸序列僅在信號肽處存在差異,其功能是否相似還有待研究。
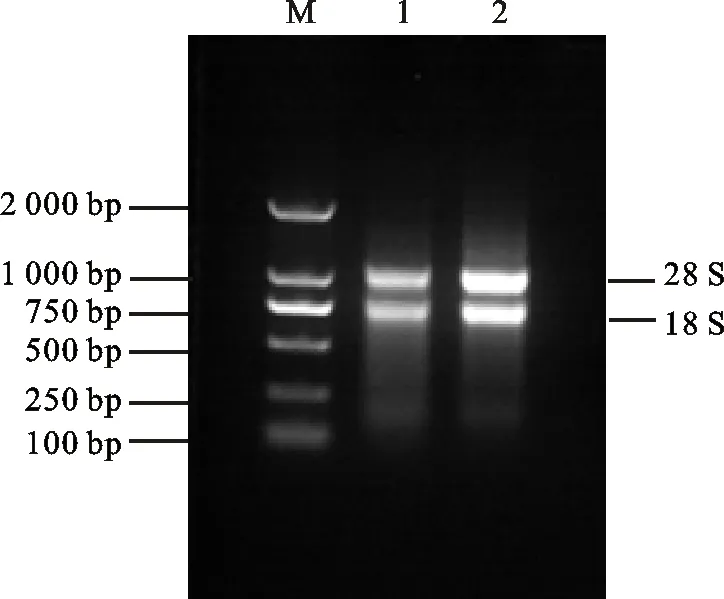
M為DL2000;1和2為樣品
M:DL2000;1 and 2:Sample
圖1東農(nóng)冬麥2號根中提取的總RNA
Fig.1TotalRNAextratedfromrootofDongnongdongmai2
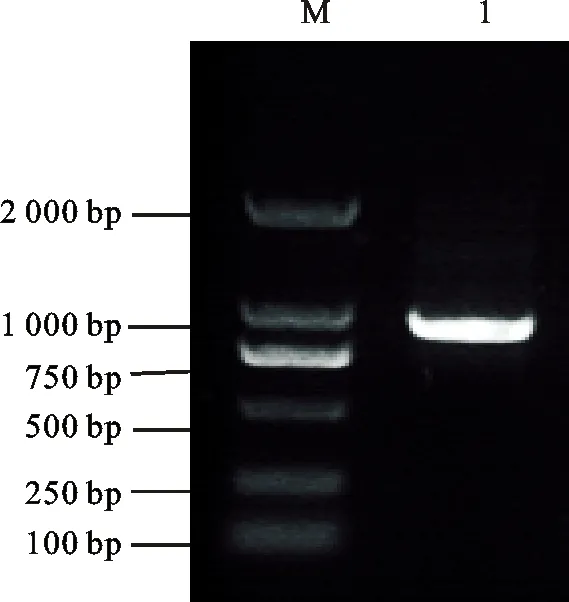
M為DL2000,1為樣品
M:DL2000, 1:Sample
圖2TaEXPA7-A/B/DCDS的PCR擴增結(jié)果
Fig.2PCRamplificationofTaEXPA7-A/B/DCDS
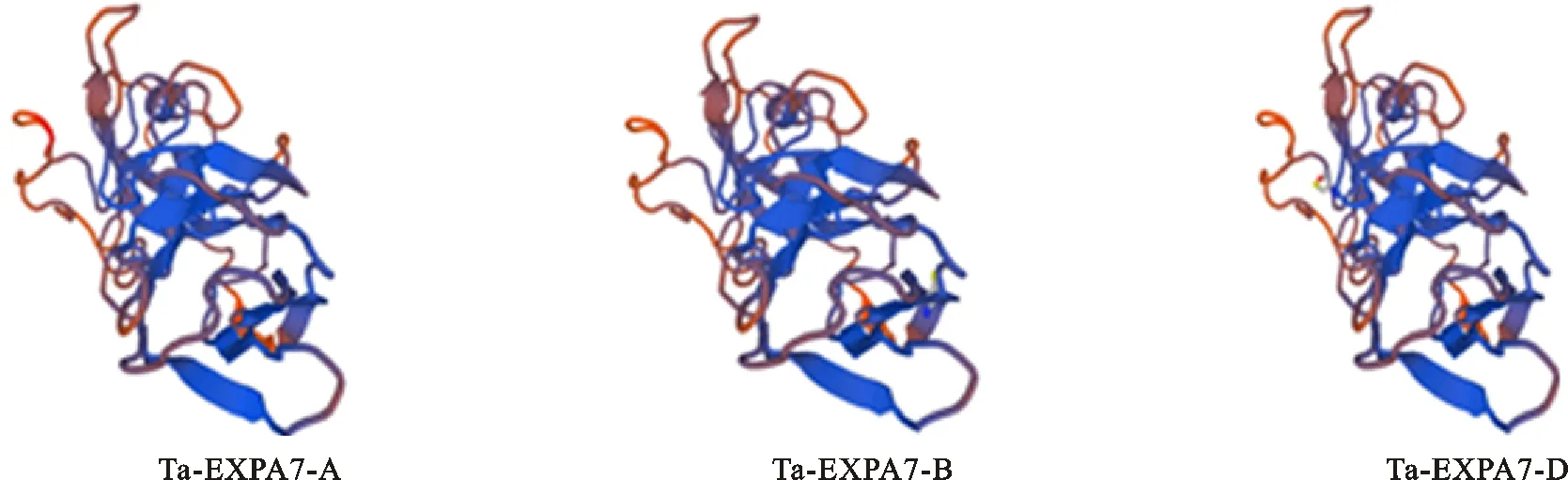
圖3 TaEXPA7-A/B/D編碼蛋白質(zhì)三級結(jié)構(gòu)的預(yù)測
2.3 TaEXPA7-A/B/D基因的序列比對和系統(tǒng)進化樹的構(gòu)建
在NCBI數(shù)據(jù)庫中進行BLAST檢索,發(fā)現(xiàn)TaEXPA7-A/B/D基因序列與節(jié)節(jié)麥(XP_020162651.1)、二穗短柄草(XP_003580384.1)、水稻(XP_015636818.1)等大多數(shù)單子葉植物的同源性較高,表明 TaEXPA7-A/B/D基因普遍存在于單子葉植物中。用ClustalX軟件對這些氨基酸序列進行多序列比對,用MEGA 6.0進行系統(tǒng)進化樹的構(gòu)建(圖5),并用BOXSHADE對這些氨基酸序列進行著色,分析其結(jié)構(gòu)(圖6)。結(jié)果表明,這些基因序列均具有信號肽和兩個保守結(jié)構(gòu)域(DPBB-1和Pollen-allerg-1),小麥與節(jié)節(jié)麥的氨基酸序列相似性最高,親緣關(guān)系最近。
2.4 TaEXPA7-A/B/D基因的表達特性分析
以獲得的RNA為模板進行反轉(zhuǎn)錄,采用qRT-PCR引物進行熒光定量PCR,測定東農(nóng)冬麥2號TaEXPA7-A/B/D在兩葉一心期根中不同組織部位的表達特性以及三個基因之間的表達差異。結(jié)果(圖7)顯示,TaEXPA7-A/B/D基因在根的根尖、伸長區(qū)和成熟區(qū)均有表達,且在根尖中的相對表達量都顯著高于伸長區(qū)和成熟區(qū)。其中,TaEXPA7-B在根尖中的相對表達量最高。
以獲得的RNA為模板進行反轉(zhuǎn)錄,采用qRT-PCR引物進行熒光定量PCR,測定冬小麥兩葉一心期根中TaEXPA7-A/B/D基因在不同環(huán)境脅迫下的表達特性以及表達差異。結(jié)果(圖8至圖10)顯示,無論在低溫、干旱還是激素處理下,該基因均下調(diào)表達。
低溫4 ℃處理后,TaEXPA7-A/B/D基因的表達量均呈現(xiàn)下降的趨勢,處理12 h時的表達量最低,在12 h之前,TaEXPA7-D的相對表達量最高,12 h以后TaEXPA7-B的相對表達量最高。
干旱處理后,TaEXPA7-A/B/D基因的表達量均呈現(xiàn)下降的趨勢,相對表達量表現(xiàn)為 TaEXPA7-B>TaEXPA7-A>TaEXPA7-D。30%PEG處理后,TaEXPA7-A/B/D基因的表達量呈現(xiàn)急劇下降的趨勢,而20%PEG處理后基因的表達量下降得較緩慢。20%PEG處理后,在12 h時,其相對表達量最低,此后又呈現(xiàn)上升的趨勢。

圖4 TaEXPA7-A/B/D的氨基酸同源比對
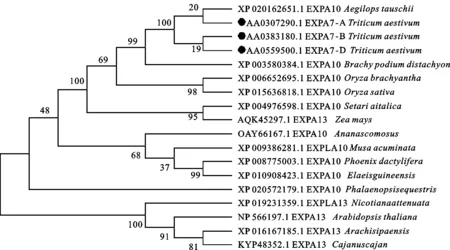
圖5 TaEXPA7-A/B/D與其他不同物種同源基因編碼蛋白的系統(tǒng)發(fā)生分析

圖6 氨基酸序列相似性分析
激素處理后,TaEXPA7-A/B/D基因的表達量均呈下降的趨勢。在MeJA處理后,在0~2 h之間,基因的表達量顯著下降,在2~48 h之間其表達量變化較小;在ABA處理后,基因的表達量呈現(xiàn)先下降后上升再下降的趨勢,TaEXPA7-D基因的表達量最低;在SA處理后,在0~2 h之間,基因的表達量顯著下降,在2 h時其表達量最低,此后,各基因的表達量隨時間的增加而緩慢上升。
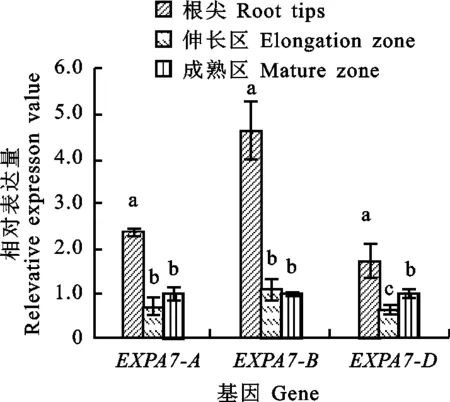
同一組圖柱上不同字母表示根的不同部位之間差異顯著(P<0.05)。
The different letter above columns indicate significant difference between different parts of the root(P<0.05).
圖7TaEXPA7-A/B/D基因在根中不同組織部位的表達差異
Fig.7DifferentialexpressionofTaEXPA7-A/B/Dindifferentregionsofroots
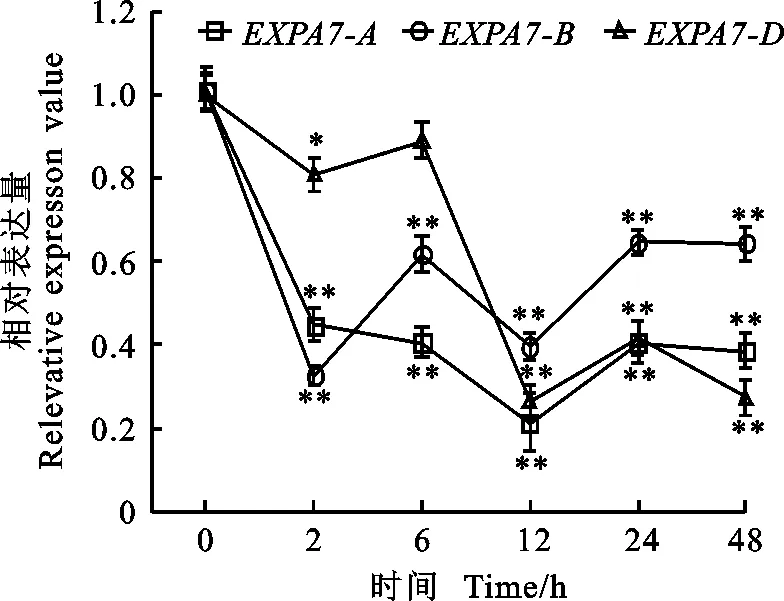
*和**分別表示與處理前差異顯著(P<0.05)和極顯著(P<0.01)。下同。
* and ** indicate difference between before and after treatment at 0.05 and 0.01 level , respectively. The same at following figures.
圖84℃脅迫下TaEXPA7-A/B/D基因的表達差異
Fig.8DifferentialexpressionofTaEXPA7-A/B/Dunder4℃stress
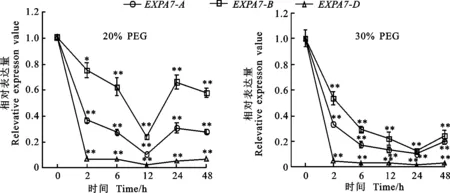
圖9 干旱脅迫下TaEXPA7-A/B/D基因的表達差異
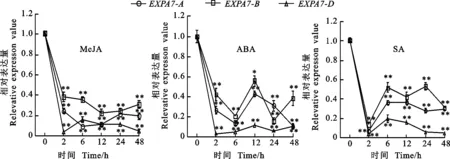
圖10 不同激素脅迫下TaEXPA7-A/B/D基因的表達差異
3 討 論
多倍化在生物學中是一種普遍存在的現(xiàn)象,是物種形成和進化的重要途徑之一。多倍體基因組中的重復(fù)基因可能有以下三種不同的命運:保持原有的功能、基因沉默和分化并執(zhí)行新的功能[10-11]。近年來,對于多倍化的研究越來越多。2007年,Shitsukawa等[12]分析了普通小麥中一個MADS 家族基因WLHS1 的3 個同源基因的結(jié)構(gòu)和功能,發(fā)現(xiàn)WLHS1-A 基因存在一個新序列的插入,WLHS1-B的DNA高度甲基化,只有WLHS1-D 是有功能的。2010年,陳 琰等[13]采用RACE方法克隆了TaEXPB8的四個同源基因,在染色體定位時發(fā)現(xiàn)了多拷貝現(xiàn)象,其表達分析結(jié)果表明,該基因在小麥初生根的伸長區(qū)表達量最高,并且受外源高濃度生長素的抑制,由此推測出該基因可能在小麥根中起著重要作用。2012年孫其信等[14]對TaEXPA1-A/B/D基因研究發(fā)現(xiàn),三個基因在幼根中的表達是沉默的,TaEXPA1-A/D在幼葉中表達,而TaEXPA1-B在幼葉中沉默,推測該基因的沉默可能與啟動子中某一區(qū)域的甲基化和乙酰化有關(guān),并對TaEXA1-A/B/D基因進行擬南芥的遺傳轉(zhuǎn)化,結(jié)果發(fā)現(xiàn)他們具有相似的功能。本試驗從東農(nóng)冬麥2號中克隆了TaEXPA7-A/B/D三個基因,發(fā)現(xiàn)其分別定位于2AL、2BL和2DL染色體上,對其核酸序列以及氨基酸序列進行分析發(fā)現(xiàn),雖然核酸序列有幾處的差異,但是氨基酸序列僅在信號肽位置有兩處差異,三個基因的表達量表現(xiàn)為 TaEXPA7-B>TaEXPA7-A>TaEXPA7-D。這三個基因是否行使相似的功能以及在多倍體基因組上的作用機制還有待研究。
當受到非生物脅迫時,植物可通過調(diào)控脅迫相關(guān)基因的表達,激活植物對脅迫的響應(yīng)機制,這些基因可以編碼控制抗逆基因表達的調(diào)節(jié)因子,也可以直接編碼與抗脅迫有關(guān)的酶和蛋白質(zhì)。因此,研究基因的表達特性對分析植物抵御非生物脅迫的作用機制是十分重要的。2016年,李 飛等[15]對低溫處理下東農(nóng)冬麥2號小麥三葉期根組織TaEXPA基因的表達分析表明,TaEXPA5、TaEXPA6和 TaEXPA7三個基因的表達量可能與其抗御低溫脅迫的能力正相關(guān)。本研究中,幼苗期根中的 TaEXAP7基因隨著4 ℃處理時間的延長呈現(xiàn)先下降后上升的趨勢,且其表達量均小于未處理時基因的表達量,由此可見,該基因在冬小麥生長的不同時期的表達量以及表達模式是不相同的,其作用機制也可能存在差異。趙美容等[16]于2016年在抗旱性不同的小麥擴展蛋白活性及基因表達分析中發(fā)現(xiàn),TaEXPA3基因的表達在干旱脅迫下上調(diào),且與品種抗旱性相關(guān),表明干旱脅迫誘導(dǎo)的擴展蛋白活性的提高與擴展蛋白表達上調(diào)有關(guān)。而本研究中, TaEXPA7-A/B/D基因的表達在干旱脅迫下下調(diào),且PEG濃度越高,其表達量隨著時間的延長下降越明顯。在膨脹素龐大的基因家族中,不同的基因行使著不同的功能,所以該基因也有可能參與和抗旱有關(guān)的作用機制。ABA、SA和JA是植物生長必要的激素,可通過復(fù)雜而精細的網(wǎng)絡(luò)來調(diào)控植物不同階段的生長發(fā)育,還可以響應(yīng)各種生物與非生物脅迫,從而提高植物對環(huán)境的適應(yīng)性。近年來,有人在外源激素影響基因表達特性的研究中發(fā)現(xiàn),茉莉酸主要通過誘導(dǎo)擴展蛋白基因的上調(diào)表達來提高擴展蛋白的活性[17-19]。 TaEXPAB23在受到MeJA的處理后其mRNA水平表達量升高。本研究中, TaEXPA7-A/B/D三個基因均屬于下調(diào)基因。該基因能否降低膨脹素活性,是否參與了植物生長的主要機制還有待研究。
最近,關(guān)于膨脹素基因與根系發(fā)育的研究已經(jīng)有許多報道。AtEXPA7、OsEXPA10、OsEXPA7、 HvEXPB7等均是與植物根系生長相關(guān)的重要基因[20-22]。
TaEXPA7-A/B/D與 OsEXPA10基因有較高的相似性,結(jié)合本試驗研究結(jié)果,推測 TaEXPA7基因也是一組和植物根系生長相關(guān)的重要基因,而根系能否正常生長發(fā)育又是冬小麥成功越冬的關(guān)鍵,因此推測該基因可能是影響冬小麥生長發(fā)育的重要基因。
[1]COSGROVE D J.Loosening of plant cell walls by expansins[J].Nature,2000,407(6802):321.
[2]SAMPEDRO J,COSGROVE D J.The expansinsuperfamily[J].GenomeBiology,2005,6(12):242.
[3] HU Z,HAN Z,SONG N,etal.Epigenetic modification contributes to the expression divergence of three TaEXPA1 homoeologs in hexaploid wheat(Triticumaestivum)[J].NewPhytologist,2013,197(4):1344.
[4]CHO H T,KENDE H.Expression of expansin genes is correlated with growth in deepwaterrice[J].PlantCell,1997,9(9):1661.
[5] 俞志明.EXPA7亞家族調(diào)控水稻根毛伸長機制研究[D].杭州:浙江大學,2010:9
YU Z M.Regulated mechanism of EXPA7 subclade members on rice root hair elongation [D].Hangzhou:Zhejiang University,2010:9.
[6] 文乙豪.水稻 OsEXPB2基因的功能研究[D].重慶:重慶大學,2013:77.
WENWEN Y H.Study on the function of rice OsEXPB2 gene [D].Chongqing:Chongqing University,2013:77.
[7] 何曉薇.水稻根毛發(fā)育相關(guān)基因 OsEXP17 的克隆和功能研究[D].杭州:浙江大學,2008:58
HE X W.Expression and function of OsEXP17 gene in rice root hair development [D].Hangzhou:Zhejiang University,2008:58.
[8]CHE J,YAMAJI N,SHEN R F,etal.An Al-inducible expansin gene, OsEXPA10 is involved in root cell elongation of rice.[J].PlantJournal,2016,88(1):132.
[9]HAN Y,LI A,LI F,etal.Characterization of a wheat(TriticumaestivumL.) expansin gene, TaEXPB23,involved in the abiotic stress response and phytohormone regulation[C]//7thCongress of Shandong Plant Physiology Society and Symposium on Plant Biology and Modern Agriculture,2012:49-58.
[10] 楊 繼.植物多倍體基因組的形成與進化[J].植物分類學報,2001,39(4):357.
YANG J.Preparation and evolution of plant polyploid genomes [J].ChineseJournalofPlantTaxonomy,2001,39(4):357.
[11] 張連全.小麥異源六倍化過程及其在遺傳育種中的應(yīng)用[D].雅安:四川農(nóng)業(yè)大學,2007:1.
ZHANG LIAN-QUAN.Application of heterotopic six-fold process of wheat and its application in genetic breeding [D].Ya’an:Sichuan Agricultural University,2007:1.
[12]SHITSUKAWA N,TAHIRA C,KASSAI K,etal.Genetic and epigenetic alteration among three homoeologous genes of a class E MADS box gene in hexaploid wheat.[J].PlantCell,2007,19(6):1723.
[13] 陳 琰,鐘伶桃,姚穎垠,等.小麥膨脹素基因 TaEXPB8部分同源cDNA序列的克隆、定位及表達分析[J].農(nóng)業(yè)生物技術(shù)學報,2010,18(4):645.
CHEN Y,ZHONG L T,YAO Y Y,etal.Cloning,mapping and expression analysis of partial homologous cDNA sequences of TaEXPB8 in wheat swelling protein [J].JournalofAgriculturalBiotechnology,2010,18(4):645.
[14]HU Z,SONG N,XING J,etal.Overexpression of three TaEXPA1 homoeologous genes with distinct expression divergence in hexaploid wheat exhibit functional retention inArabidopsis[J].PlosOne,2013,8(5):e63667.
[15] 李 飛,王曉磊,徐永清,等.低溫處理下東農(nóng)冬麥1號小麥根組織EXPA基因的表達分析[J].麥類作物學報,2016,36(9):1159.
LI F,WANG X L,XU Y Q,etal.Expression ofEXPAgene in wheat root tissue of Dongnongdongmai No.1 wheat under low temperature treatment [J].ActaMetallurgicaSinica,2016,36(9):1159.
[16] 趙美榮,李永春,劉 輝,等.抗旱性不同的小麥擴展蛋白活性及基因表達分析[J].江蘇農(nóng)業(yè)科學,2016,44(1):108.
ZHAO M R,LI Y CH,LIU H,etal.Expression of wheat protein and gene expression in wheat with different drought resistance [J] .JiangsuAgriculturalSciences,2016,44(1):108.
[17] 韓翠英.干旱脅迫下四個功能基因的表達模式與不同品種小麥的抗旱相關(guān)性分析[D].楊凌:西北農(nóng)林科技大學,2015:31.
HAN C Y.Study on the relationship between the expression patterns of four functional genes and the drought resistance of different varieties of wheat under drought stress [D].Yangling:Northwest A&F University,2015:31.
[18] 段龍飛.茉莉酸信號途徑上關(guān)鍵基因家族COI/JAZ/MYC分子進化分析[D].楊凌:西北農(nóng)林科技大學,2013:7.
DUAN L F.Study on evolution of COI/JAZ/MYC molecules in key gene family of jasmonic acid signal pathway [D].Yangling:Northwest A&F University,2013:7.
[19] 王文艷.葡萄水楊酸和茉莉酸信號轉(zhuǎn)導(dǎo)途徑中4個重要基因的克隆及表達分析[D].南京農(nóng)業(yè)大學,2011:6.
WANG W Y.Cloning and expression analysis of four important genes in salicylic acid and jasmonic acid signal transduction pathway [D].Nanjing:Nanjing Agricultural University,2011:6.
[20]HE X,ZENG J,CAO F,etal. HvEXPB7,a novel β-expansin gene revealed by the root hair transcriptome of Tibetan wild barley,improves root hair growth under drought stress [J].JournalofExperimentalBotany,2015,66(22):7405.
[21]XU Q,SANALKUMAR K,EMILY M,etal.Gibberellin regulation and genetic variations in leaf elongation for tall fescue in association with differential gene expression controlling cell expansion[J].ScientificReports,2016,6:30258.
[22]LIN C,CHOI H S,CHO H T.Root hair-specific EXPANSIN A7 is required for root hair elongation inArabidopsis[J].MoleculesandCells,2011,31(4):393.
CloningandCharacterizationofTaEXPA7PartialHomologousGenesinRootsofWinterWheatDongnongdongmai2
DONGJiamin1,XUYongqing1,PengLina1,FENGXu1,YAOShukuan1,ZHAOQiaoqin1,LIFenglan1,HUBaozhong1,2
(1.College of Life Science,Northeast Agriculture University,Harbin,Heilongjiang 150030,China;2.Harbin University,Harbin,Heilongjiang 150086,China)
Expansin is a protein that induces cell wall relaxation and elongation during plant growth and development, which containsEXPA,EXPB,EXLAandEXLBfour gene families,playing an important role in the formation and rapid development of plant roots. In this experiment, the root of cold winter wheat Dongnongdongmai 2 at two leaf stage was used as the material to clone the three CDS full-length of the TaEXPA7 gene.Their amino acid sequences were highly homologous with only two different amino acids at signal peptide.The nucleic acid sequence length was 777 bp, encoding 258 amino acids.These three genes were named as TaEXPA7-A, TaEXPA7-B, TaEXPA7-D, located at 2AL, 2BL and 2DL chromosomes, respectively. The protein contained two conserved domains, DPBB-1 and Pollen-allerg-1, with molecular weight of 27 670.74 Da and isoelectric point of 8.09 and they were hydrophobic proteins. The similarity between them andAegilopstauschiiandBrachypodiumdistachyonwas 99% and 92%, respectively. qRT-PCR analysis of roots of winter wheat at two leaves stage showed that TaEXPA7-A/B/D expressed higher in root tips, followed by elongation zone and mature zone. The winter wheat roots were treated with low temperature, drought and hormone and the three genes were all down regulated,the relative expression of TaEXPA7-B gene was higher and that of TaEXPA7-D was lower,which indicated that genes in different chromosomes of polyploid wheat responded differently to environmental stresses,their gene expression and response patterns were different, and the dominant chromosomes were also different under different environmental stresses. Moreover, the expression of these genes in roots may be related to stress such as low temperature, drought and exogenous hormones.They may be important gene for root growth.
Winter wheat; TaEXPA7; Partial homology; Expression characteristics
時間:2017-11-14
網(wǎng)絡(luò)出版地址:http://kns.cnki.net/kcms/detail/61.1359.S.20171114.1027.018.html
2017-05-25
2017-07-08
國家基礎(chǔ)科學人才培養(yǎng)基金項目(J1210069);中俄國際合作項目(2013DFR30270)
E-mail:1970113717@qq.com
李鳳蘭(E-mail:lifenglan@neau.edu.cn);胡寶忠(E-mail:bzhu@neau.edu.cn)
S512.1;S330
A
1009-1041(2017)11-1419-09
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
民用飛機設(shè)計與研究(2020年4期)2021-01-21 09:15:02
小天使·一年級語數(shù)英綜合(2019年8期)2019-08-27 02:23:00
中國特種設(shè)備安全(2018年11期)2019-01-08 02:08:32
電子制作(2018年18期)2018-11-14 01:48:24
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
山東工業(yè)技術(shù)(2016年15期)2016-12-01 05:31:22
鄭州大學學報(醫(yī)學版)(2015年2期)2015-02-27 14:50:46
中國中醫(yī)藥現(xiàn)代遠程教育(2014年11期)2014-08-08 13:23:44
山東女子學院學報(2014年6期)2014-03-01 02:24:55
