極端低氣壓處理后低溫高溫交替對(duì)發(fā)狀念珠藻的光合生理活性的影響
2017-12-05 05:28:22劉穎慧褚亞東陸洪斌曹旭鵬
載人航天 2017年6期
劉穎慧,褚亞東,陸洪斌,曹旭鵬,薛 松
(中國(guó)科學(xué)院大連化學(xué)物理研究所,大連 116023)
極端低氣壓處理后低溫高溫交替對(duì)發(fā)狀念珠藻的光合生理活性的影響
劉穎慧,褚亞東,陸洪斌,曹旭鵬,薛 松?
(中國(guó)科學(xué)院大連化學(xué)物理研究所,大連 116023)
為考察發(fā)狀念珠藻在類地行星環(huán)境中生存的可行性,搭建了模擬類地行星環(huán)境的極端低氣壓實(shí)驗(yàn)裝置,并研究極端低氣壓與低溫-高溫交替作用對(duì)發(fā)狀念珠藻光合生理活性的影響。結(jié)果顯示,在氣體組成為CO2和N2(體積比為95∶5)的氛圍中,在-20℃、100 Pa、600 Pa和1200 Pa氣壓下,分別處理發(fā)菜4天、7天、15天、30天和60天,發(fā)菜的呼吸和光合放氧速率與未處理樣品沒有差別。此外,不同氣壓下,-20℃到25℃交替處理發(fā)菜10天、20天和30天,發(fā)菜的光合及呼吸速率與正常相比也沒有差異。以上結(jié)果表明,發(fā)菜能夠生存在極端低氣壓下的低溫-高溫交替環(huán)境。通過分析樣品的脂肪酸組成和游離脯氨酸含量發(fā)現(xiàn),處理后樣品的游離脯氨酸含量升高,脂肪酸組成也發(fā)生了相應(yīng)的變化,這些變化是維持發(fā)菜耐受極端低氣壓和低溫-高溫交替的物質(zhì)基礎(chǔ)。
發(fā)菜;低氣壓;光合;呼吸;脯氨酸
1 引言
微藻在空間站上被廣泛用于循環(huán)氮、水、CO2以獲得有機(jī)物[1-2],其中藍(lán)藻可以在極端惡劣環(huán)境生長(zhǎng)[3]。發(fā)狀念珠藻,俗稱發(fā)菜,是一種陸生藍(lán)藻,主要分布在干旱、半干旱地區(qū)[4]。我國(guó)是發(fā)菜分布最豐富的國(guó)家。發(fā)菜產(chǎn)地自然環(huán)境的特殊性及其變化節(jié)律已構(gòu)成發(fā)菜賴以生存的必需條件,使得其形成了獨(dú)特的生態(tài)學(xué)特性:極強(qiáng)的耐旱性、喜中溫而耐變溫、耐太陽輻射、耐鹽堿、耐貧瘠、耐大風(fēng)和熱干風(fēng)[5]。發(fā)菜的這些獨(dú)特的生物特性使得發(fā)菜很有可能在類地行星環(huán)境上生存。從公開報(bào)道中看到,以德國(guó)達(dá)姆施塔特工業(yè)大學(xué)為代表的一些研究團(tuán)隊(duì)計(jì)劃開展在火星溫室中篩選藻種的工作,也有報(bào)道表明美國(guó)國(guó)家航空航天局(NASA)嘗試將藍(lán)藻應(yīng)用于星際旅行和火星探索[6]。近來Fraunhofer-Gesellschaft報(bào)道兩種藻屬(一種綠藻和一種藍(lán)藻)能夠生活在國(guó)際空間站外面16個(gè)月[7]。然而,目前并沒有微藻在類地行星環(huán)境下的極端低氣壓下的研究報(bào)道。
本研究通過搭建模擬類地行星環(huán)境低氣壓裝置,利用LED光源模擬類地行星表面光照,將發(fā)菜培養(yǎng)在極端低氣壓及極端低氣壓處理后低溫高溫交替處理?xiàng)l件下,借助氧電極測(cè)定樣品的光合放氧和呼吸速率,并通過氣相色譜等分析方法測(cè)定樣品的脂肪酸、脯氨酸含量變化。
2 材料和方法
2.1 實(shí)驗(yàn)材料
發(fā)菜采自內(nèi)蒙古阿拉善盟,實(shí)驗(yàn)前已干燥存儲(chǔ)2年。
2.2 極端低氣壓處理器搭建
為了模擬類地行星環(huán)境低氣壓,我們?cè)O(shè)計(jì)了如圖1所示的實(shí)驗(yàn)裝置圖。通過選用高真空玻璃管,稱取1 g發(fā)菜放在玻璃管中,將氣體置換為95%體積的CO2和5%體積的N2后,通過真空泵抽取真空,通過壓力傳感器測(cè)定壓力,使測(cè)試壓力條件分別為100 Pa、600 Pa和1200 Pa,然后利用火焰噴燈將玻璃管密封,將發(fā)菜保存于極端低氣壓保存管內(nèi),使其在長(zhǎng)時(shí)間存放后仍保持所需氣壓及氣體組成(圖2)。
2.3 培養(yǎng)方法
為了考察發(fā)菜耐受極端溫度和極端低氣壓程度,將發(fā)菜低氣壓處理器放在-20℃冰柜中,通過溫度傳感器全程監(jiān)測(cè)溫度,如圖3所示,并設(shè)定不同處理時(shí)間(4天、7天、15天、30天和60天)。利用LED光源模擬類地行星表面光照,光譜波長(zhǎng)范圍為420 nm到680 nm,最大波長(zhǎng)為560 nm,其光譜圖見圖4。光照階段光強(qiáng)以地表光強(qiáng)40%(800 μmol photons/m2/s)為上限成正弦波變化,光暗比14 h∶10 h。
此外,為了考察不同低氣壓下-20℃到25℃交替循環(huán)對(duì)發(fā)菜存活的影響。以天為循環(huán)周期,將存有發(fā)菜的低壓處理器放在-20℃黑暗處理10 h,再將發(fā)菜轉(zhuǎn)移到25℃光照處理14 h,光照條件如圖4裝置所示,分別處理10天、20天和30天。通過溫度傳感器全程監(jiān)測(cè)溫度。
2.4 呼吸速率和光合放氧速率測(cè)定
稱取處理后發(fā)菜和未處理發(fā)菜各0.01 g,用去離子水清洗發(fā)菜3~4次,放在液體BG11培養(yǎng)基,25℃、40 μmol photons/m2/s下培養(yǎng)24 h。待樣品培養(yǎng)好后,將發(fā)菜剪成長(zhǎng)度小于0.5 cm的片段,置于反應(yīng)杯中。每個(gè)樣品設(shè)置了3~6個(gè)重復(fù),測(cè)定方法采用oxygenmeter model 782(Strathkelvin,Glasgow,UK)氧電極測(cè)定,測(cè)定條件為:25℃,樣品3 mL,光強(qiáng)為950 μmol photons/m2/s,通過加入少量Na2SO3粉末標(biāo)定零刻度,以空氣飽和的蒸餾水標(biāo)定氧電極的滿刻度[8]。
2.5 脂肪酸含量測(cè)定
稱取處理后發(fā)菜20 mg測(cè)定脂肪酸組成,其方法參考文獻(xiàn)[9]。具體步驟:在10 mL圓底燒瓶加入20 mg樣品和小轉(zhuǎn)子,之后用100 μL注射器取50 μL濃度為2 mg/mL的C17 TAG溶液作為內(nèi)標(biāo),之后加入5 mL 2%的硫酸-甲醇溶液后,70℃600 rpm油浴反應(yīng)1 h,冷凝回流。反應(yīng)結(jié)束后,冷卻20 min至室溫后,加入0.75 mL的去離子水和2 mL的正己烷,磁力攪拌器攪拌1 min,之后轉(zhuǎn)移到15 mL離心管中,2000 rpm離心2 min,取上層相1 mL加入到氣相色譜樣品瓶中進(jìn)行分析。氣相色譜(GC)檢測(cè)條件:色譜柱為DB-23色譜柱(30 m length×0.32 mm,I.D?0.25 μm film),F(xiàn)ID檢測(cè)器溫度270℃,進(jìn)樣口溫度270℃,升溫程序?yàn)槠鹗紲囟?30℃,之后10℃/min升到170℃,最后2.5℃/min升到200℃,進(jìn)樣量1 μL。每個(gè)樣品做了2個(gè)重復(fù)。
2.6 脯氨酸含量測(cè)定
將處理后發(fā)菜樣品研磨成粉末,由北京質(zhì)譜醫(yī)學(xué)研究有限公司采用HPLC-MS/MS進(jìn)行LC液相(戴安公司Ultimate3000)、MS質(zhì)譜(美國(guó)AB公司API 3200 Q TRAP)測(cè)定。每個(gè)樣品測(cè)定了1次。
2.7 統(tǒng)計(jì)學(xué)分析
通過STATISTICA?7.0(StatSoft Inc.,Tulsa,OK,USA)軟件,采用方差分析(ANOVA)和多重比較Tukey’s(HSD)分析實(shí)驗(yàn)數(shù)據(jù)。
3 結(jié)果和討論
3.1 極端溫度和極端低氣壓下發(fā)菜的呼吸和光合速率變化
表1顯示的是不同氣壓下分別處理4天、7天、15天、30天和60天的發(fā)菜的呼吸和光合作用速率。結(jié)果顯示:在-20℃,氣體組成為95%體積的CO2和5%體積的N2、氣壓為100 Pa、600 Pa和1200 Pa(分別相當(dāng)于大氣壓的0.1%,0.6%和1.2%),無論是處理4天、7天、15天、30天甚至是60天,發(fā)菜的呼吸和光合作用恢復(fù)與未處理發(fā)菜樣品之間沒有顯著性差異,它們的光合速率平均在400 μmol O2/h/g(DW),呼吸速率大約在100 μmol O2/h/g(DW)。這說明發(fā)菜能夠耐受低氣壓和極端溫度,這種特性不會(huì)隨著時(shí)間的延長(zhǎng)而發(fā)生改變,也表明發(fā)菜能夠長(zhǎng)期耐受極端溫度和極端低氣壓。然而,對(duì)于發(fā)菜是如何抵御這種環(huán)境條件的機(jī)制有待于后續(xù)進(jìn)一步研究。
3.2 低溫-高溫反復(fù)交替下發(fā)菜的呼吸和光合速率變化
表2是不同極端低氣壓下,-20℃到25℃交替處理10天、20天和30天的發(fā)菜的光合及呼吸速率。從表中可以看出,不論-20℃到25℃交替處理時(shí)間是10天、20天還是30天,經(jīng)過處理的發(fā)菜的光合放氧和呼吸速率樣品與正常相比沒有差異,仍然保持在大約光合放氧為400 μmol O2/h/g(DW)和呼吸作用速率為100 μmol O2/h/g(DW)。這說明發(fā)菜在100 Pa、600 Pa和1 200 Pa條件下,-20℃到25℃處理不同時(shí)間對(duì)發(fā)菜的光合部件沒有造成傷害,或者是發(fā)菜啟動(dòng)了相應(yīng)的防護(hù)機(jī)制以抵御這種環(huán)境條件所造成的傷害,從而保護(hù)發(fā)菜的光合部件,導(dǎo)致發(fā)菜的光合活性沒有受到損傷。以上實(shí)驗(yàn)結(jié)果說明發(fā)菜能夠在極端低氣壓下耐受低溫-高溫反復(fù)交替,這表明了發(fā)菜在低氣壓下的長(zhǎng)期冷熱頻繁交替不會(huì)影響其生命活動(dòng)。然而,對(duì)于發(fā)菜是如何抵御這種環(huán)境條件的機(jī)制有待于后續(xù)進(jìn)一步研究。

表1 -20℃不同氣壓下處理不同時(shí)間發(fā)菜光合和呼吸速率Table 1 The respiration and photosynthetic oxygen release rate of N.flagelliforme treated with different gas pressures at-20℃for different timeμmol O2/h/g(DW)
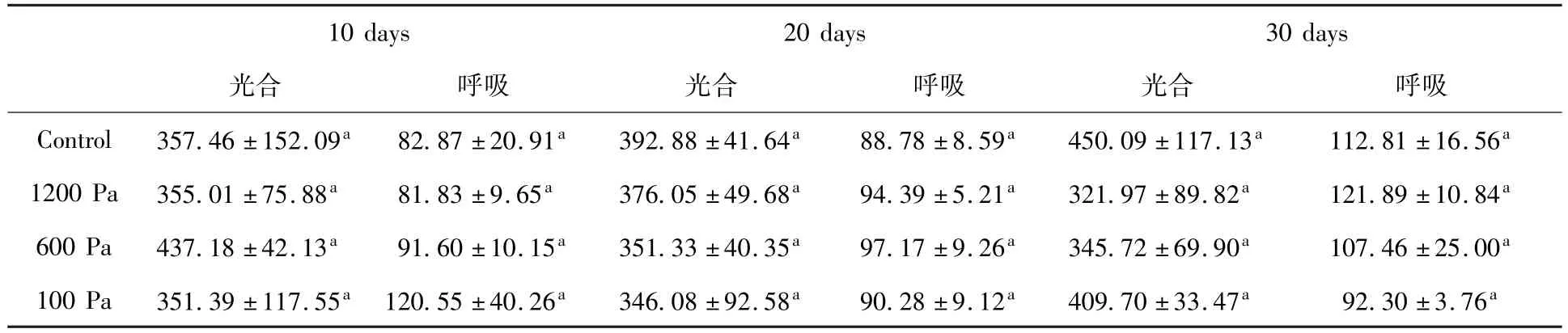
表2 -20℃到25℃不同氣壓下交替處理發(fā)菜的呼吸和光合作用速率Table 2 The respiration and photosynthetic oxygen release rate of N.flagelliforme treated with different gas pressures in-20℃to 25℃alternation for different timeμmol O2/h/g(DW)
3.3 發(fā)菜脂肪酸組成和脯氨酸含量變化
生物膜是生物將自身與環(huán)境分開的第一道屏障,這也導(dǎo)致生物膜是環(huán)境脅迫過程中發(fā)生損害的主要位點(diǎn)。目前很多研究表明不飽和脂肪酸在生物應(yīng)對(duì)各種環(huán)境脅迫起到了很重要的作用[10-13]。脯氨酸是分布最廣泛的一種滲透物質(zhì),大量研究表明植物細(xì)胞內(nèi)的脯氨酸增加有利于抵抗不良環(huán)境[14-16]。
為了揭示發(fā)菜能夠長(zhǎng)期耐受極端溫度和極端低氣壓以及低溫-高溫反復(fù)交替的機(jī)理,我們通過氣相色譜測(cè)定了處理樣品的脂肪酸組成并委托北京質(zhì)譜醫(yī)學(xué)研究有限公司采用液質(zhì)連用測(cè)定了脯氨酸含量變化。從表3數(shù)據(jù)顯示,發(fā)菜在1200 Pa、600 Pa和100 Pa氣壓下,-20℃到25℃交替處理30天,其脂肪酸組成主要是C16:0、C16:1、C18:0、C18:1、C18:2和C18:3,其中C16:1和C18:3所占脂肪酸比例最高,分別大約為26%和30%。C18:0分別為1.38%、1.44%和1.27%,C18:1n9分別為5.02%、3.77%和2.28%,相比正常樣品的C18:0為0.92%有顯著性上升,這可能會(huì)增強(qiáng)膜的流動(dòng)性。C18/C16脂肪酸比例在正常和1200 Pa、600 Pa和100 Pa氣壓下分別為1.16、1.21、0.89和1.15,可以看出在600 Pa低溫高溫交替下,C18/C16比例數(shù)值有所下降,但是沒有構(gòu)成顯著性差異。以上結(jié)果顯示發(fā)菜在1200 Pa、600 Pa和100 Pa氣壓下,-20℃到25℃交替處理30天,發(fā)菜脂肪酸組成發(fā)生了變化,C18:0有顯著性上升,這些可能與發(fā)菜耐受低溫高溫交替有關(guān)。此外不飽和脂肪酸占總脂肪酸比例與正常相比沒有顯著性差異,說明發(fā)菜不飽和脂肪酸占總脂肪酸比例在低氣壓下低溫高溫交替很穩(wěn)定,這對(duì)于維持發(fā)菜正常活性起到一定的保障作用。
此外,如表4所示,不同氣壓下處理60天發(fā)菜脂肪酸變化情況和不同氣壓下低溫高溫交替規(guī)律很相似,C18:1n9比對(duì)照上升,總的不飽和脂肪酸所占比例與正常沒有顯著差異,而C18:C16在600 Pa處理后數(shù)值有所下降,但是相比正常沒有顯著性變化。然而有研究指出發(fā)菜隨著培養(yǎng)溫度從30℃降到15℃,C18:C16是下降的趨勢(shì)[17]。這說明發(fā)菜在響應(yīng)低壓脅迫與溫度脅迫的脂肪酸反應(yīng)策略不一樣。
從表5中可以看到,在1200 Pa、600 Pa和100 Pa氣壓下,-20℃到25℃交替處理30天發(fā)菜的脯氨酸含量分別為51.04 μg/g(DW)、28.53 μg/g(DW)和47.29 μg/g(DW),而正常發(fā)菜的脯氨酸含量為28.32 μg/g(DW)。因此,1200 Pa和100 Pa氣壓下,-20℃到25℃交替處理30天,發(fā)菜的脯氨酸含量分別相比正常樣品增加了80%和67%。大量研究表明植物細(xì)胞內(nèi)的脯氨酸增加有利于抵抗不良環(huán)境。脯氨酸親水性強(qiáng),既有親水部分又有疏水部分,可以與蛋白質(zhì)的疏水部分結(jié)合,不影響蛋白質(zhì)內(nèi)部的疏水部分,使蛋白質(zhì)不致變性,對(duì)蛋白質(zhì)起到一定的保護(hù)作用。此外,脯氨酸可以與細(xì)胞內(nèi)的一些化合物形成聚合物,類似親水膠體,有一定的保護(hù)作用,可防止細(xì)胞脫水[14]。因此,發(fā)菜可能通過調(diào)節(jié)胞內(nèi)游離的脯氨酸含量抵抗極端低氣壓所造成的傷害。另外,在-20℃、1200 Pa氣壓下處理60天,發(fā)菜脯氨酸37.38 μg/g(DW),比未處理樣品上升了約32%,而600 Pa和100 Pa氣壓下,發(fā)菜脯氨酸含量沒有顯著性差異,這可能是因?yàn)檫@種處理對(duì)發(fā)菜沒有造成傷害,所以脯氨酸含量沒有增加。
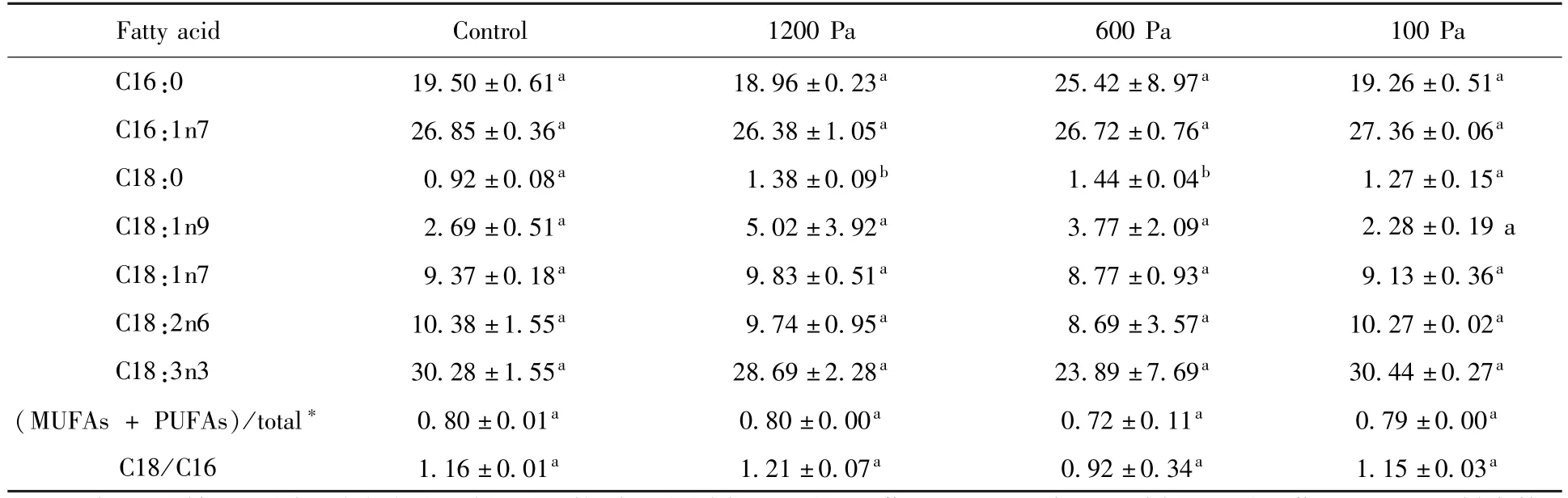
表3 -20℃到25℃不同氣壓下交替處理30天發(fā)菜的脂肪酸組成Table 3 Changes of fatty acid composition(%)in N.flagelliforme treated with different gas pressure in-20℃to 25℃alternation for 30 days
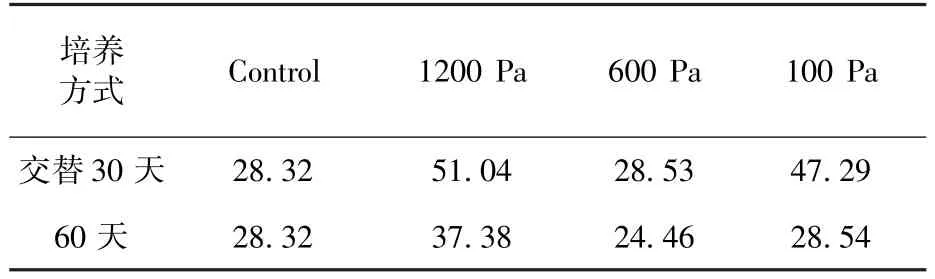
表5 -20℃不同氣壓下處理60天發(fā)菜和-20℃到25℃不同氣壓下交替處理30天的脯氨酸變化Table 5 The proline change of N.flagelliforme treated with different gas pressures in-20℃to 25℃alternation for 30 days and at-20℃for 60 days respectivelyμg/g(DW)
4 結(jié)論
1)當(dāng)發(fā)菜在-20℃、氣體組成為95%CO2、5%N2、氣壓分別為100 Pa、600 Pa和1200 Pa長(zhǎng)期處理達(dá)60天,發(fā)菜的光合速率與正常相比沒有差異,平均在400 μmol O2/h/g(DW),呼吸速率大約在100 μmol O2/h/g(DW),說明發(fā)菜能夠長(zhǎng)期耐受低溫和極端低氣壓;
2)當(dāng)發(fā)菜在氣體組成為95%CO2、5%N2,氣壓分別為100 Pa、600 Pa和1200 Pa,在-20℃到25℃交替處理30天,發(fā)菜的光合及呼吸速率與正常相比沒有差異,大約光合放氧為400 μmol O2/h/g(DW),呼吸作用速率為100 μmol O2/h/g(DW),說明發(fā)菜能夠在極端低氣壓下耐受低溫-高溫反復(fù)交替;
3)發(fā)菜能夠抵御極端低氣壓和低溫高溫反復(fù)交替的原因可能包含調(diào)節(jié)胞內(nèi)脂肪酸組成以及游離脯氨酸含量。
(References)
[1] 艾為黨,郭雙生,董文平,等.“平微藻-小白鼠”二元生態(tài)系統(tǒng)氣體交換規(guī)律研究[J].載人航天,2014,20(6):510-516.Ai W D,Guo S S,Dong W P,et al.Research on gas exchange in the Microalgae-Mice system[J].Manned Spaceflight,2014,20(6):510-516.(in Chinese)
[2] 艾為黨,郭雙生,董文平.受控生態(tài)生保系統(tǒng)螺旋藻藻種篩選研究[J].載人航天,2013,19(6):58-63.Ai W D,Guo S S,Dong W P.Study on selection of Spirulina for Controlled ecological life support system[J].Manned Spaceflight,2013,19(6):58-63.(in chinese)
[3] Whitton B.Introduction to the Cyanobacteria,Ecology of Cyanobacteria:Their Diversity in Time and Space[M].Netherlands Kluwer Academic Publishers,2003:1-11.
[4] Gao K.Chinese studies on the edible blue-green alga,Nostoc flagelliforme:a review[J].Journal of Applied Phycology,1998,10(1):37-49.
[5] 錢凱先,朱浩然,陳樹谷.發(fā)菜的生態(tài)條件及其規(guī)律分析[J].植物生態(tài)學(xué)與地植物學(xué)學(xué)報(bào),1989,13(2):97-105.Qian K X,Zhu H R,Chen S G.The ecological conditions for Nostoc flagelliforme and their analysis[J].Acta Phytoecologicaet Geobotanica Sinica,1989,13(2):97-105.(in Chinese)
[6] 羅輯.德國(guó)科學(xué)家研究如何讓藍(lán)藻改造火星大氣[J/OL].http://tech.qq.com/a/20141228/002512.htm.Luo J.German scientists investigate how to make cyanobacteria transform Martian atmosphere[J/OL].http://tech.qq.com/a/20141228/002512.htm.
[7] Fraunhofer-Gesellschaft.Algae survive heat,cold and cosmic radiation[J/OL].https://www.sciencedaily.com/releases/2017/02/170207104231.htm.
[8] Liu Y H,Yu L,Ke W T,et al.Photosynthetic recovery of Nostoc flagelliforme(Cyanophyceae)upon rehydration after 2 years and 8 years dry storage[J].Phycologia,2010,49(5):429-437.
[9] Liu J,Liu Y,Wang H.et al.Direct transesterification of fresh microalgal cells[J].Bioresource technology,2015,176:284-287.
[10] 年洪娟,陳麗梅.不飽和脂肪酸在逆境脅迫中的作用[J].中國(guó)微生態(tài)學(xué)雜志,2012,24(8):760-762.Nian H J,Chen L M.The role of unsaturated fatty acids in various environmental stresses[J].Chinese Journal of Microecology,2012,24(8):760-762.(in Chinese)
[11] Inaba M,Suzuki I,Szalontai B,et al.Gene-engineered rigidification of membrane lipids enhances the cold inducibility of gene expression in Synechocystis[J].Journal of Biological Chemistry,2003,278(14):12191-12198.
[12] Zhang J,Liu H,Sun J,et al.Arabidopsis fatty acid desaturase FAD2 is required for salt tolerance during seed germination and early seedling growth[J].PLoS One,2012,7(1):e30355.
[13] Yazawa H,Iwahashi H,Kamisaka Y,et al.Production of polyunsaturated fatty acids in yeast Saccharomyces cerevisiae and its relation to alkaline pH tolerance[J].Yeast,2009,26(3):167-184.
[14] 梅俊學(xué).逆溫下發(fā)菜脯氨酸含量及質(zhì)膜透性的變化與含水量的關(guān)系[J].山東師大學(xué)報(bào)(自然科學(xué)版),2000,15(2):178-181.Mei J X.The change of proline content and plasma membranepermeability of dry and wet Nostoc flagelliformein temperature inversion[J].Journal of Shandong Normal University(Natural Science),2000,15(2):178-181.(in Chinese)
[15] Battah M G.Alleviation of salinity stress on growth and some metabolites of Anabaena constricta and Nostoc linckia using L-proline or D L-B phenylalanine[J].Australian Journal of Basic and Applied Science,2009,3(3):1903-1909.
[16] Fatma T,Khan M,Choudhary M.Impact of environmental pollution on cyanobacterial proline content[J].Journal of Applied Phycology,2007,19(6):625-629.
[17] Liu X J,Jiang Y,Chen F.Fatty acid profile of the edible filamentous cyanobacterium Nostoc flagelliforme at different temperatures and developmental stages in liquid suspension culture[J].Process biochemistry,2005,40(1):371-377.
Effects of Low-high Temperature Alternations on Photosynthetic Physiology of Nostoc flagelliforme after Extreme Low Gas Pressure Exposure
LIU Yinghui,CHU Yadong,LU Hongbin,CAO Xupeng,XUE Song?
(Dalian Institute of Chemical Physics,Chinese Academy of Sciences,Dalian 116023,China)
In order to examine whether Nostoc flagelliforme can survive on terrestrial planets or not,an experimental apparatus providing extremely low air pressure for simulating terrestrial planet environment was constructed,and theeffects of low-high temperature alternation on the photosynthetic physiology of Nostoc flagelliforme under extremely low air pressures were investigated.It was found thatunder a gas atmosphereof CO2/N2(95/5,v/v),the respiration and photosynthetic oxygen release rate of N.flagelliforme were not affected when it was treated with various extremely low gas pressures(100 Pa,600 Pa,1200 Pa)for 4 days,7 days,15 days,30 days and 60 days at-20℃,
respectively.In addition,when N.flagelliforme was subjected to-20℃to-25℃alternation under different low gas pressuresfor 10 days,20 days and 30 days,the respiration and photosynthetic oxygen release rates showed also no difference as compared with the control.These results indicated that N.flagelliforme might survive under an extremely low gas pressure with low-high temperature alternations.Furthermore,the free proline contents increased and fatty acid composition changed when N.flagelliforme was treated with the extremely low gas pressure and low-high temperature alternations,which could be the material foundations for the tolerance capability of N.flagelliforme in response to these extreme environments.
Nostoc flagelliforme;low air pressure;photosynthesis;respiration;proline
Q935
A
1674-5825(2017)06-0829-06
2017-04-24;
2017-09-19
劉穎慧,女,博士,副研究員,研究方向?yàn)樗{(lán)藻光合和脅迫生理。Email:lyh8423@163.com
?通訊作者:薛松,女,博士,研究員,研究方向?yàn)槲⒃迳锬茉础mail:xuesong@dicp.ac.cn
(責(zé)任編輯:龐迎春)
