新疆外來入侵植物意大利蒼耳和刺蒼耳種子的越冬性能
2017-11-23 02:39:03李杰馬淼
生態學報 2017年21期
李 杰 馬 淼
石河子大學生命科學學院, 石河子 832003
新疆外來入侵植物意大利蒼耳和刺蒼耳種子的越冬性能
李 杰 馬 淼*
石河子大學生命科學學院, 石河子 832003
入侵新疆的外來植物意大利蒼耳和刺蒼耳的果實于深秋季節成熟后,便開始陸續脫落。脫離母體的果實可能會落置在干燥的地面上,還有可能會掉落在水體中,還有部分果實會保留在枝條上并不脫落直到翌年春天。由于越冬生境的差異會直接影響翌年春天種子的萌發活性,為了評價外來入侵植物意大利蒼耳和刺蒼耳的入侵能力,預測其潛在分布區,為對其實施有效防控提供科學指導。故模擬了其種子在新疆自然條件下可能遭遇的冬季生境類型,設計了4種實驗處理,即低溫換水浸泡(模擬了落入流水或大型水體中結冰前的生境)、低溫不換水浸泡(模擬了落入小型靜水中結冰前的生境)、冰凍(模擬了落入水體中結冰后的生境)和干燥冷凍處理(模擬了掉落在干燥地面或一直停留在植株上的生境類型),并以室溫干燥儲存為對照。處理90 d后,進行種子萌發實驗,結果顯示干凍、冰凍及低溫換水浸泡處理后意大利蒼耳的萌發率與對照組無顯著差異,不換水浸泡處理降低了意大利蒼耳種子的活力,顯著降低了其萌發能力。而干凍處理對刺蒼耳種子的萌發沒有影響,冷水浸泡(換水浸泡和不換水浸泡)顯著降低刺蒼耳的萌發率,與對照組相比分別下降了37%、65%。冰凍處理后刺蒼耳沒有萌發。說明意大利蒼耳種子對新疆的各種越冬生境均有較好的耐受性,而刺蒼耳種子對低溫和水淹環境的耐受性較差。鑒于二者種子萌芽習性的特點,建議在全疆范圍內的中生、濕生和季節性水澇生境中對意大利蒼耳實施全方位的監控,農田、林下、路邊、渠旁、湖泊及水庫周圍都有可能成為其潛在的入侵地;對刺蒼耳的重點監測區域則應當放在低緯度、低海拔的中生和中旱生生境,尤其要切實加強南疆各國際口岸進口貨物的檢疫工作。
入侵植物;意大利蒼耳;刺蒼耳;種子;越冬性能
隨著我國向西開放戰略部署的深化,以及“一帶一路”政策下新絲綢之路各國國際間貨運貿易往來的日趨頻繁,一大批外來物種悄然涌入了新疆,尤其是菊科蒼耳屬(XanthiumL.)的植物意大利蒼耳(XanthiumitalicumMoretti)與刺蒼耳(X.spinosumLinnaeus)正以極快的速度向新疆各地蔓延擴張,對當地的農業和畜牧業造成了嚴重危害[1],對新疆本就脆弱的綠洲生態系統而言無疑是雪上加霜。對其入侵機理的闡釋,入侵能力的評價和潛在分布區的預測是對該物種實施有效防控的必要前提。然而,目前對意大利蒼耳和刺蒼耳的研究僅限于形態描述[2- 3],分布區范圍[2,4],和化學組分及其化感作用[5- 7]等方面,對其入侵機理和種群擴張趨勢的研究尚未見報道。
種子萌發是植物生活史中的一個關鍵環節[8],其對植物繁衍種族、擴展分布區域、提高環境適應能力具有重要意義[9]。作為繁殖體的種子,其對環境適應能力的強弱會直接影響個體在新生境中的存活。年降水量、年均溫度、年極端低溫以及年均積溫等是影響外來植物地理分布的重要因素[10]。入侵地的冬季低溫環境是許多原產于溫熱地區的外來植物所面臨的嚴峻挑戰[11],在被引入地區是否能夠安全越冬便成了許多外來植物入侵成功的限制瓶頸[12]。因此,對外來入侵植物種子越冬性能的研究不僅是闡釋其入侵機理的關鍵環節,也是評價其入侵能力和潛在分布區的重要依據[13]。新疆高山與盆地相間,荒漠與濕地并存[14],外來植物的種子不僅要經受冬季陸地的嚴寒,而且還有可能遭遇到浸泡于冰水環境的考驗。故本研究運用實驗生態學的研究方法,模擬各種可能遭遇到的自然越冬生境,研究了意大利蒼耳和刺蒼耳種子經歷了不同越冬條件后的萌發狀況,旨在通過探討不同生境類型與其種子越冬性能之間的聯系,為其入侵機理的闡釋,入侵能力的評價和潛在分布區的預測提供科學依據。
1 材料和方法
1.1 種子采集
兩種植物的果實于2014年10月采自新疆伊寧市周邊(81°0′—83°59′E,42°50′—44°03′N),將果實快速運回實驗室風干備用。
1.2 處理方法
1.2.1 處理設置
挑選450粒健康成熟、大小一致的果實,依據其在新疆可能遭遇到的4種越冬生境類型,設置了相應的4種實驗處理,并以室溫條件下的干燥儲存處理為對照,具體如下(本文中的果實實際上是指具總苞的果序):
(1)室溫干燥儲存:取90枚果實,存放在黑暗、干燥的室溫環境中,溫度為20 ℃。
(2)低溫換水浸泡處理:為了避免長期水浸條件下滋生的大量微生物對實驗結果的影響,我們對實驗器皿和蒸餾水進行了高壓滅菌,對果實進行了表面消毒。具體如下:取90枚果實,置于燒杯中用蒸餾水沖洗3遍,以清除總苞表面的灰塵與雜物,然后轉入70%的酒精溶液中浸泡30 s,再轉入0.1%的升汞中浸泡10 min進行果實表面消毒,用無菌水沖洗后,最后轉入滅過菌的500 mL三角瓶中,浸泡在400 mL的無菌水里,用無菌過濾透氣封口膜封口,置于5 ℃黑暗的冷藏室中,每天換水1次(以上步驟均在超凈工作臺上進行),以此來模擬落入流水但未曾結冰的果實的生境類型。
(3)低溫不換水浸泡處理:種子消毒及封裝同上,置于5 ℃黑暗的冷藏室中儲存,以此來模擬落入小型靜水中的果實的生境類型。
(4)干燥冷凍處理:取90枚果實,存放在黑暗、干燥冷凍環境中,溫度為-20 ℃[15],以此來模擬落置在干燥的地面上或一直著生在植株上不曾脫落的果實的生境類型。
(5)冰凍處理:取90枚果實,置于塑料燒杯中用蒸餾水沖洗3遍,以清除總苞表面的灰塵與雜物,在5 ℃條件下浸泡24 h后,轉移至-20 ℃的冰箱中冷凍儲藏,以此來模擬落入水體中且水體結冰后的果實的生境類型。
以上各組處理的時間均為90 d。
1.2.2 萌發實驗
處理90 d后將果實取出進行種子萌發實驗。將室溫干燥和干燥冷凍條件下儲存的果實在室溫條件下用蒸餾水浸泡24 h,使總苞軟化;將冰凍處理的果實從冰箱中取出后在室溫下解凍。由于總苞中兩粒種子具有不同的發芽特性[16],為了避免其對實驗結果的影響,在不對內部種子造成傷害的條件下小心地剝去果序外的總苞,取其中較大的種子均勻地擺放在盛有河沙的塑料萌發盒(20 cm×12 cm×5 cm)中進行萌發實驗,埋藏深度為1 cm,河沙的相對濕度為70%。每盒中擺放30枚種子,每種處理設置3組重復。將萌發盒置于光照培養箱,在每天12 h光照,光照強度為450 μmol m-2s-1,晝夜溫度為25 ℃/20 ℃的條件下培養15 d,萌發過程中始終保持70%土壤相對濕度,以子葉露出土面為發芽依據,每24 h統計一次發芽情況。
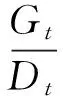
1.2.3 種子活力檢測
萌發實驗處理15 d后,從各組處理的萌發盒中隨機挖出意大利蒼耳和刺蒼耳未發芽的種子各60枚,清水洗凈表面的沙土,用刀片將胚橫切成薄片后置于潔凈的在載玻片上,用TTC染色法染色,檢測種子活力。
1.3 數據處理
用SPSS 19.0軟件進行實驗數據的統計分析,不同處理下的種子萌發率和發芽指數采用單因素方差分析(One-way ANOVA),用Duncan多重比較(Duncan′s multiple range test)檢測不同處理間的差異顯著性水平。
2 結果與分析
2.1 種子發芽勢
室溫干燥處理下的意大利蒼耳種子從第3天開始萌發,其他4種處理下的種子從第4天開始萌發,但不同處理對種子發芽勢的影響顯著(圖1)。低溫不換水浸泡處理極顯著地降低了意大利蒼耳種子的發芽勢(P<0.01);室溫干燥處理的種子的發芽勢最高(95.56%)但與換水浸泡、干凍及冰凍處理無顯著差異(P>0.05)。
不同條件處理對刺蒼耳的種子萌發造成了強烈影響,室溫干燥處理下的種子在第4天就開始萌發了,不換水浸泡的種子在第7天才開始萌發,而冰凍條件下的種子至試驗結束都沒有種子萌發(圖2)。干凍處理下與室溫干燥處理的種子的發芽勢均為42.22%;低溫水浸泡極顯著地降低了刺蒼耳種子的發芽勢。
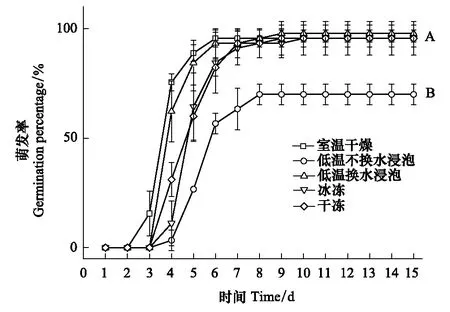
圖1 5種處理下意大利蒼耳種子的累積萌發率 Fig.1 The germination percentage of seeds of X. italicum under 5 treaments各曲線所標字母不同,表示曲線對應的處理間在第15天時的差異達到極顯著水平(P<0.01)
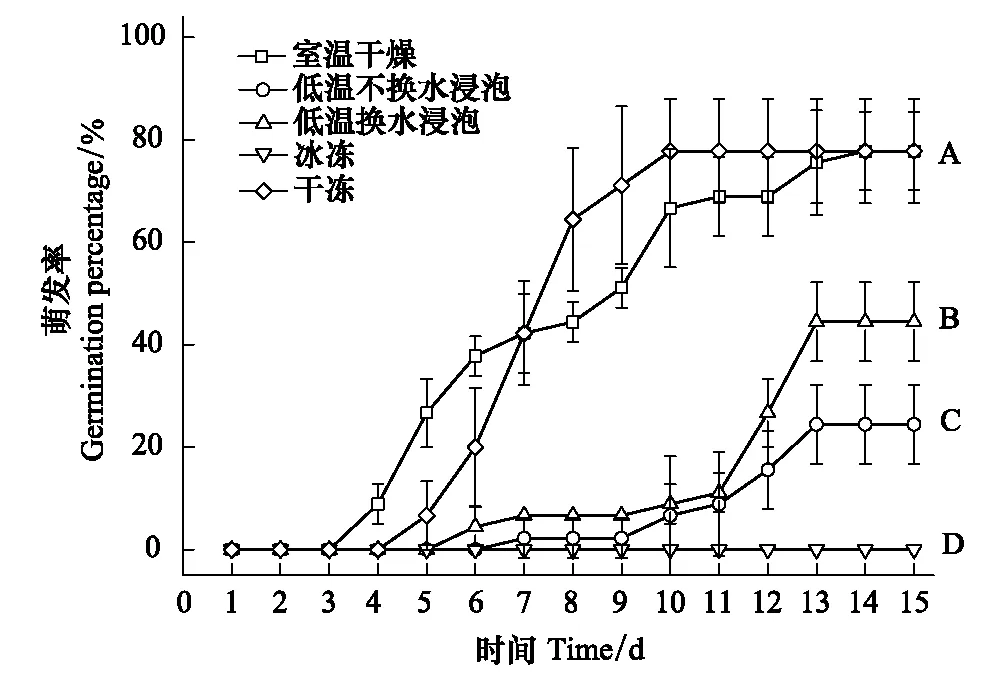
圖2 5種處理下刺蒼耳種子的累積萌發率 Fig.2 The germination percentage of seeds of X. spinosum under 5 treaments
2.2 種子萌發率
種子萌發率與7 d的發芽勢結果一致,低溫不換水浸泡處理下意大利蒼耳的種子發芽率僅為70%,極顯著低于其他4種處理(P<0.01);室溫干燥、換水浸泡、干凍及冰凍處理的種子萌發率無顯著差異,均在90%以上。
冰凍處理后的刺蒼耳種子無萌發,室溫干燥處理及干凍處理下刺蒼耳種子的萌發率均為70%,顯著高于水浸泡處理(不換水浸泡和換水浸泡)。并且換水浸泡處理下的種子萌發率極顯著高于不換水浸泡處理的種子萌發率(P<0.01),分別比對照組下降了37%和65%。
2.3 發芽指數
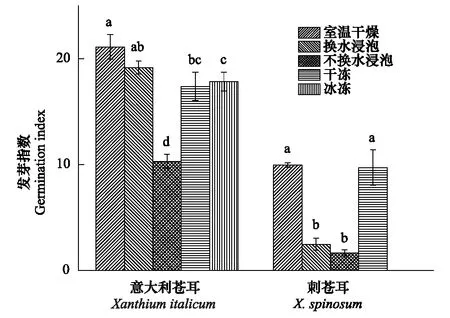
圖3 5種處理下意大利蒼耳和刺蒼耳種子的發芽指數 Fig.3 The seed germination index of X. italicum and X. spinosum under 5 treaments同一植物下,不同小寫字母表示差異達到顯著水平(P<0.05)
意大利蒼耳在室溫干燥儲存處理下種子的發芽指數最高,但和模擬流水的冷水浸泡處理(勤換水)間無顯著差異(P>0.05);冰凍和干凍處理的種子的發芽指數無顯著差異(P>0.05);模擬靜水的冷水浸泡處理(不換水)后種子的發芽指數及顯著低于其他4種處理(P<0.01)。經室溫干燥處理的刺蒼耳種子的發芽指數最高,但與干凍干燥處理無顯著差異(P>0.05);其次為冷水浸泡處理(換水及不換水處理)。在相同處理下,意大利蒼耳的發芽指數均大于刺蒼耳的發芽指數(圖3)。
2.4 種子活力檢測
意大利蒼耳種子活力檢測結果顯示60枚未發芽的種子均已不具有活力,部分種子已經腐爛;而刺蒼耳的60枚未萌發的種子中只有4枚尚具活性,但活力較低,其余種子的活力也均已完全喪失。因此,在本研究中將以萌發率的高低作為種子活力強弱的評價指標。
3 討論
干燥冷凍是對落置在干燥的地面或保留在植株上不曾脫落的果實越冬生境的模擬,研究表明干燥冷凍處理會顯著降低意大利蒼耳種子的發芽指數,但其對意大利蒼耳種子的發芽勢和萌發率卻無影響。而干燥冷凍處理對刺蒼耳的萌發無顯著影響。發芽勢、萌發率和發芽指數分別反應種子的發芽整齊度、種子質量和種子發芽速率的重要指標。結果表明,新疆的冬季低溫環境會使落置在干燥的地面或保留在植株上不曾脫落的意大利蒼耳種子的萌發進程滯后,但對最終的萌發率并無顯著影響,而對刺蒼耳種子的萌發無顯著影響。意大利蒼耳和刺蒼耳的種子可在此生境中安全越冬。
在濕生環境中,成熟種子脫落后往往被浸泡入水中,使水淹成為濕生環境中的主要環境脅迫,對種子萌發有很大影響。有研究表明,長期的水浸泡會降低蒼耳種子的萌發率和活力,并且其萌發率隨著水淹時間的延長而逐漸降低[18]。水淹對種子萌發的影響主要有以下兩方面的原因:①水中的溶解氧分壓遠低于空氣,種子的正常呼吸代謝被抑制;②胚中的無機鹽、蛋白質、淀粉等物質的滲出會損壞胚,從而影響了種子的活力[19]。
刺蒼耳在水浸泡處理(換水浸泡和不換水浸泡)90 d后,其發芽勢和萌發率與室溫干燥儲藏的對照組相比并顯著降低,說明刺蒼耳種子經過長時間的水浸泡后,種胚損壞使得種子活力降低。同時,與換水浸泡相比較,不換水浸泡處理下浸泡種子的水中的氧分壓的下降抑制了種子正常的呼吸代謝,加重了種子活力的下降。表現為換水浸泡的種子的萌發率要高于不換水浸泡處理。
對意大利蒼耳而言,低溫換水浸泡90 d后,意大利蒼耳種子的發芽勢和萌發率可高達93.33%和97.78%,與對照組無顯著差異。說明長時間的水浸泡并不會對意大利蒼耳種子的胚造成破壞。而不換水浸泡處理下,意大利蒼耳種子的發芽勢和發芽率分別為63.33%和70%,發芽指數也顯著低于換水浸泡處理。而導致這一差異的原因可能是由于缺氧導致的種子活力的喪失,因此,以高密度掉落在不流動淺水水域中的意大利蒼耳種子越冬安全性較低。由于在自然條件下淺水水體中的種子一般很難達到90粒/400mL的密度水平,所以掉落在冷水中的意大利蒼耳種子具有較高的越冬安全性。
結冰是新疆冬季常見的自然事件,冰凍處理僅僅降低了意大利蒼耳種子的發芽指數,對發芽勢和萌發率無顯著影響。然而刺蒼耳果實遇水結冰后對種子活力的影響極為強烈,在本試驗中沒有種子萌發。因此,刺蒼耳的種子在濕地中很難越冬。
意大利蒼耳和刺蒼耳于2012年在新疆被首次記錄[2],意大利蒼耳現已廣泛分布于博州、伊犁、塔城、額敏、阿勒泰、石河子、昌吉和烏魯木齊地區,刺蒼耳則主要分布于烏魯木齊、昌吉、伊犁等地[3,20]。意大利蒼耳和刺蒼耳分布格局的差異恰好可以由二者種子萌發特性的差別得到解釋。意大利蒼耳種子對新疆冬季的水熱條件均有較好的耐受性,因此其分布區的緯度跨度較大,在緯度高氣溫偏低的塔城、額敏和阿勒泰地區均有大面積的分布,在中生、濕生和季節性水澇的環境中也均可生長,生境選擇多樣;然而由于刺蒼耳種子對低溫和水淹環境的耐受性差,故其分布區集中于低緯度的天山北坡沿線,傾向于中生環境的農田或較干燥的荒漠草原等生境類型。雖然二者目前的入侵區域僅限于北疆,然而與北疆相比,地處暖溫帶的南疆冬季氣溫相對和暖,對意大利蒼耳與刺蒼耳種子的安全越冬會更加有利。之所以目前在南疆地區還尚未見分布,主要原因可能是天山屏障的阻隔。但是隨著新疆作為國家向西開放橋頭堡地位的確定和南北疆之間以及南疆與諸多鄰國之間貿易往來的日趨密切,天山的地理屏障效應很有可能會被打破,因此在全疆視野下綜合考慮防治措施可能會更加有效。
外來入侵植物是否能夠成功入侵,主要取決于物種自身的入侵力(invasiveness)與生境的可入侵性(invisibility)[21- 23]。入侵力主要取決于該物種自身的生物學特性,而生境的可入侵性則取決于生境的非生物環境及生物環境對入侵的抵抗力[24- 25]。外來植物自身的特性對入侵、生存和擴展極為重要[26],除了在新引種區內缺乏天敵[27]和具有空余生態位[28- 29]以外,其物種本身對環境適應能力的廣譜性[30]也是外來生物得以成功入侵的重要原因。
鑒于二者種子萌芽習性的特點,建議在全疆范圍內的中生、濕生和季節性水澇生境中對意大利蒼耳實施全方位的監控,農田、林下、路邊、渠旁、湖泊及水庫周圍都有可能成為其潛在的入侵地;對刺蒼耳的重點監測區域則應當放在低緯度、低海拔的中生和中旱生生境,尤其要切實加強南疆各國際口岸進口貨物的檢疫工作。
[1] 張祥林, 張偉, 吳衛. 新疆植物檢疫性有害生物. 北京: 中國質檢出版社, 2012: 112- 113.
[2] 杜珍珠, 徐文斌, 閻平, 王少山, 郭一敏. 新疆蒼耳屬3種外來入侵新植物. 新疆農業科學, 2012, 49(5): 879- 886.
[3] 裴會明. 入侵植物刺蒼耳的形態學特征與防治. 甘肅林業科技, 2015, 40(1): 24- 25, 46- 46.
[4] 宋珍珍, 譚敦炎, 周桂玲. 入侵植物刺蒼耳在新疆的分布及其群落特征. 西北植物學報, 2012, 32(7): 1448- 1453.
[5] 董芳慧, 劉影, 蔣夢嬌, 李海冰, 胡云霞. 入侵植物刺蒼耳對小麥和苜蓿種子的化感作用. 干旱區研究, 2014, 31(3): 530- 535.
[6] Andreani S, Barboni T, Desjobert J M, Paolini J, Costa J, Muselli A. Essential oil composition and chemical variability ofXanthiumitalicumMoretti from Corsica. Flavour and Fragrance Journal, 2012, 27(3): 227- 236.
[7] 邵華, 邰鳳嬌, 韓彩霞. 外來植物意大利蒼耳入侵對土壤微生物的影響及其生態效應 // 中國第七屆植物化感作用學術研討會論文摘要集. 昆明: 中國植物保護學會植物化感作用專業委員會, 2015: 1-2
[8] 李曉梅, 趙曉英, 趙法, 郭名軍. 新疆3種錦雞兒屬植物種子萌發對溫度和土壤水分的響應. 草業科學, 2009, 26(10): 140- 145.
[9] 高蕊, 魏巖, 嚴成. 角果藜的地上地下結果性與種子萌發行為. 生態學雜志, 2008, 27(1): 23- 27.
[10] 張帥, 郭水良, 管銘, 印麗萍, 張若軒. 我國入侵植物多樣性的區域分異及其影響因素——以74個地區數據為基礎. 生態學報, 2010, 30(16): 4241- 4256.
[11] 魯萍, 桑衛國, 馬克平. 外來入侵種飛機草在不同環境脅迫下抗氧化酶系統的變化. 生態學報, 2006, 26(11): 3578- 3585.
[12] 邱寵華, 王奇志, 余巖. 外來入侵假臭草在中國分布區的預測. 應用與環境生物學報, 2011, 17(6): 774- 781.
[13] Xu Z L. Potential distribution of invasive alien species in the upper Ili river basin: determination and mechanism of bioclimatic variables under climate change. Environmental Earth Sciences, 2015, 73(2): 779- 786.
[14] 邱娟, 地里努爾·沙里木, 譚敦炎. 入侵植物黃花刺茄在新疆不同生境中的繁殖特性. 生物多樣性, 2013, 21(5): 590- 600.
[15] 李景林, 張山清, 普宗朝, 王命全, 王勝蘭, 趙書琴. 近50a新疆氣溫精細化時空變化分析. 干旱區地理, 2013, 36(2): 228- 237.
[16] 吳冬, 黃姝博, 李宏慶. 意大利蒼耳二形性種子萌發、植株生長差異及生態適應性. 生態學報, 2009, 29(10): 5258- 5264.
[17] 宋松泉, 程紅焱, 龍春林, 姜孝成. 種子生物學研究指南. 北京: 科學出版社, 2005: 57- 61.
[18] 王強, 袁興中, 劉紅, 林芳淼, 陳忠禮, 張躍偉. 水淹對三峽水庫消落帶蒼耳種子萌發的影響. 濕地科學, 2011, 9(4): 328- 333.
[19] 宋松泉, 程紅焱, 姜孝成. 種子生物學. 北京: 科學出版社, 2008: 105- 240.
[20] 馬金雙. 中國外來入侵植物調研報告. 北京: 高等教育出版社, 2014: 233- 233.
[21] 李博, 徐炳聲, 陳家寬. 從上海外來雜草區系剖析植物入侵的一般特征. 生物多樣性, 2001, 9(4): 446- 457.
[22] Chaudhary N, Narayan R, Sharma D. Differential biomass allocation to plant organs and their allelopathic impact on the growth of crop plants: A case study on the invasibility of Ageratum conyzoides in Indian dry tropics. Indian Journal of Agricultural Science, 2015, 85(11): 1404- 1411.
[23] Gorgone-Barbosa E, Pivello V R, Baeza M J, Fidelis A. Disturbance as a factor in breaking dormancy and enhancing invasiveness of African grasses in a Neotropical Savanna. Acta Botanica Brasilica, 2016, 30(1): 131- 137.
[24] Alpert P, Bone E, Holzapfel C. Invasiveness, invasibility and the role of environmental stress in the spread of non-native plants. Perspectives in Plant Ecology, Evolution and Systematics, 2000, 3(1): 52- 66.
[25] 李博, 陳家寬. 生物入侵生態學: 成就與挑戰. 世界科技研究與發展, 2002, 24(2): 26- 36.
[26] 彭少麟, 向言詞. 植物外來種入侵及其對生態系統的影響. 生態學報, 1999, 19(4): 560- 568.
[27] Allen A C, Keith E O. Using the West Indian manatee (Trichechus manatus) as a mechanism for invasive aquatic plant management in Florida. Journal of Aquatic Plant Management, 2015, 53: 95- 104.
[28] Johnson S D, Raguso R A. The long-tongued hawkmoth pollinator niche for native and invasive plants in Africa. Annals of Botany, 2016, 117(1): 25- 36.
[29] Verhelst P, Boets P, Van Thuyne G, Verreycken H, Goethals P L M, Mouton A M. The distribution of an invasive fish species is highly affected by the presence of native fish species: evidence based on species distribution modelling. Biological Invasions, 2016, 18(2): 427- 444.
[30] Kelley A L, De Rivera C E, Buckley B A. Cold tolerance of the invasiveCarcinusmaenasin the east Pacific: molecular mechanisms and implications for range expansion in a changing climate. Biological Invasions, 2013, 15(10): 2299- 2309.
Seedoverwinterperformanceoftwoinvasiveplants:XanthiumitalicumMorettiandX.spinosumLinnaeusinXinjiang
LI Jie, MA Miao*
CollegeofLifeScience,ShiheziUniversity,Shihezi832003,China
The fruits of the Italy cocklebur (XanthiumitalicumMoretti) and stab cocklebur (X.spinosumLinnaeus), two kinds of invasive plants in Xinjiang, begin to fall after it matures in late autumn. Some of the fruits detach from mother plant and drop onto dry ground, some fall in the water, and others remain on the branches and do not detach until the next spring. The habitat to which seeds are exposed to in winter directly affects seed germination in the following year. To assess their invasiveness, predict their potential distribution area, and provide scientific guidance for effective prevention and control of these invaders, four kinds of natural winter habitats in which seeds might be exposed in Xinjiang were simulated (i.e., cold flowing water, cold dead water, cold dead water followed by freezing, and cold dry soil or cold air). Treatments in which seeds were soaked in frequently changed cold water (5 ℃), soaked in still cold water (5 ℃), frozen in the air less than -20 ℃, and soaked in still water for 24 h then frozen with the water less than -20 ℃were designed. Seeds kept at dry room temperature served as controls. After 90-day pretreatment, seed germination experiments were conducted. The results showed that: (1) For the Italy cocklebur, the germination percentage of seeds soaked in unchanged cold water was significantly lower than other treatments. Others were not significantly different. (2) For stab cocklebur, seeds that were stored at dry room temperature and frozen in the air had the highest germination percentage (70%), and germination percentage of the seeds soaked in changed and unchanged water was 37% and 65% lower than that of the control. The soaked and frozen seeds did not germinate. It was suggested that multiple habitats, including those with moderate moisture, and wet and seasonally flooded habitats throughout Xinjiang should be strictly monitored for Italy cocklebur, including farmland, understory, roadsides, riversides, and around lakes and reservoirs, which might be potential invasion areas. The key monitoring area for stab cocklebur should have moderate moisture and dry habitats at low latitude and low elevation. The quarantine of imported goods in international ports around southern Xinjiang should be strengthened.
invasive plants;XanthiumitalicumMoretti;X.spinosumLinnaeus; seed; overwinter performance
國家自然科學基金項目(31360047)
2016- 08- 28; < class="emphasis_bold">網絡出版日期
日期:2017- 07- 11
*通訊作者Corresponding author.E-mail: mamiaogg@126.com
10.5846/stxb201608281750
李杰,馬淼.新疆外來入侵植物意大利蒼耳和刺蒼耳種子的越冬性能.生態學報,2017,37(21):7181- 7186.
Li J, Ma M.Seed overwinter performance of two invasive plants:XanthiumitalicumMoretti andX.spinosumLinnaeus in Xinjiang.Acta Ecologica Sinica,2017,37(21):7181- 7186.
猜你喜歡
國畫家(2022年2期)2022-04-13 09:07:46
四川文學(2021年4期)2021-07-22 07:11:54
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
絲綢之路(2014年9期)2015-01-22 04:24:46
兒童與健康(2011年4期)2011-04-12 00:00:00
