蝦夷扇貝生殖腺凝膠狀酶解物的 流變特性
2017-11-21 10:49:42,,,,,,,,*
食品工業科技 2017年21期
, , , ,,, ,,*
(1.大連工業大學食品學院,國家海洋食品工程技術研究中心,遼寧大連 116034; 2.海南出入境檢驗檢疫局,海南海口 570311)
蝦夷扇貝生殖腺凝膠狀酶解物的 流變特性
閻佳楠1,李毅2,唐越1,張萌1,商文慧1,杜椅楠1,姜卉1,吳海濤1,*
(1.大連工業大學食品學院,國家海洋食品工程技術研究中心,遼寧大連 116034; 2.海南出入境檢驗檢疫局,海南海口 570311)
本文對蝦夷扇貝生殖腺凝膠狀酶解物的流變特性進行了研究,為蝦夷扇貝的綜合利用奠定研究基礎。以蝦夷扇貝雄性生殖腺為原料,選用木瓜蛋白酶作為工具酶,通過SDS-PAGE分析酶解過程中蛋白質分子量的變化,利用流變儀考察酶解液的流變特性;利用脫氧核糖核酸酶(DNaseⅠ)在前期對樣品進行孵育,探討DNA分子對于其流變特性的作用影響。結果顯示,蝦夷扇貝雄性生殖腺經木瓜蛋白酶(30000 U/g蛋白)酶解15~180 min后,蛋白質發生顯著降解。在不同流變儀掃描模式下,酶解液的儲存模量G′、損耗模量G″、黏度η、復數黏度η*均明顯高于未酶解樣品。酶解3 h時,應變掃描中,應變為1%時,G′、G″分別達到了188.9 Pa和17.9 Pa;頻率掃描中,頻率為0.1 Hz時,η*達到230.9 Pa·s。經DNaseⅠ處理后,酶解液流變參數(G′、G″及η*)顯著降低,但略高于未酶解樣品。上述結果說明,蝦夷扇貝生殖腺凝膠狀酶解物以彈性占主導,具有明顯的剪切稀化現象,且其流變特性可能與肽分子和DNA分子有關。
蝦夷扇貝,雄性生殖腺酶解物,凝膠,流變特性
食品膠體是一類潤滑、黏稠或膠凍狀的物質,因其具有穩定、乳化、懸浮食品顆粒的作用,并可賦予食品適宜的口感,在食品工業中得到廣泛應用[1]。食品膠體通過氫鍵、范德華力或離子鍵橋的作用,形成空間網狀結構,進而形成凝膠狀物質[2]。近年來,隨著國民健康意識的增強,在保證食品功能特性的前提下,對食品營養價值的要求越來越高。因此,開發新型營養型食品膠體具有切實意義。
蝦夷扇貝(Patinopectenyessoensis)是重要的海產貝類,富含蛋白質等營養成分。2015年,全國扇貝總產量達到178.5萬噸[3]。在肥滿期,蝦夷扇貝生殖腺是扇貝的重要可食部位,極具開發價值。本文前期研究發現,經中性蛋白酶酶解后的蝦夷扇貝雄性生殖腺呈凝膠狀態,當達到適當濃度時,其硬度、凝聚性等指標可與食用膠相當,具有廣闊的應用前景[4-6]。
研究顯示,針對某些蛋白質原料,蛋白酶酶解可促進微粒間聚集作用,增加疏水結構,而形成空間三維網狀構象,使產品體現出一定的凝膠特性。例如大豆蛋白[7-8]、乳清蛋白[9-10]等在適當酶解條件下,均顯示出良好的凝膠狀態。另外,也有研究顯示,經胰蛋白酶水解后的卵黃顆粒,其流變性能也隨凝膠的形成顯著增強[11]。流變學主要用于研究物質的流動與形變的特性[12-14]。在食品工業中,流變特性可反映食品受到外力干擾后,其結構和性質的變化,近年來被廣泛應用于食品膠體及食品原料的特性研究中[15-17]。因此,借助流變特性這一相關有效研究手段,可對蝦夷扇貝生殖腺酶解物凝膠特性進行深入探究。
綜上所述,本文將研究對象鎖定為蝦夷扇貝雄性生殖腺,使用木瓜蛋白酶進行酶解,獲得凝膠狀酶解物,通過聚丙烯酰氨凝膠電泳(SDS-PAGE)了解其分子量隨時間變化情況;利用流變儀研究蝦夷扇貝生殖腺酶解物的流變特性。此外,鑒于雄性生殖腺中含有一定的脫氧核糖核酸,利用DNA酶(DNaseⅠ)研究DNA分子對酶解物流變特性的影響,為深入探究蝦夷扇貝生殖腺酶解物凝膠形成機制奠定研究基礎,以期挖掘蝦夷扇貝酶解物凝膠在食品膠體工業中的潛在應用價值。
1 材料與方法
1.1材料與試劑
肥滿期的蝦夷扇貝(3月初購買,貯存于-80 ℃冰箱) 大連長興水產品批發市場;木瓜蛋白酶(酶活為30000 U/g) 生工生物工程(上海)有限公司;DNA降解酶(DNaseⅠ酶,酶活為70 U/μL) 寶生物工程(大連)有限公司;其他試劑 均為國產分析純。
1.2儀器與設備
DF-101S型集熱式恒溫加熱磁力攪拌器 鞏義市予華儀器有限責任公司;Sartorius PB-10型pH計 塞多利斯科學儀器(北京)有限公司;AE-6450型垂直電泳儀 日本ATTO株式會社;MF-ChemiBIS 2.0型凝膠成像儀、DNR Bio-imaging;Discovery HR-1型旋轉流變儀 美國TA儀器;2KBTES-55型真空冷凍干燥機 美國Virtis公司;CF16RXⅡ型冷凍離心機 日本日立公司;BCD-203C型電冰箱 杭州華日電冰箱股份有限公司。
1.3實驗方法
1.3.1 原料處理 選取鮮活蝦夷扇貝,分離雄性生殖腺,乳白色,用去離子水將其洗凈后,煮沸10 min使內源酶失活,待冷卻至室溫,將其進行凍干,收集凍干粉貯藏于-80 ℃冰箱備用。
1.3.2 蝦夷扇貝雄性生殖腺酶解物的制備 以蝦夷扇貝雄性生殖腺凍干粉為原料,用去離子水配制成底物蛋白含量為4%(m/w)的懸濁液,采用0.1 mol/L NaOH溶液調整溶液pH至7.0,加入木瓜蛋白酶,加酶量為3000 U/g蛋白質、50 ℃、pH7.0的外界條件下酶解15~180 min,之后在100 ℃條件下滅酶10 min,之后將酶解液冷卻備用。
在考察DNaseⅠ對蝦夷扇貝雄性生殖腺酶解物流變特性影響時,在加入木瓜蛋白酶前,采用DNaseⅠ在室溫下處理原料,加酶量為500 U/mL,180 min后,經100 ℃滅酶10 min,冷卻后,按照1.3.2方法利用木瓜蛋白酶進行酶解(3 h)。
1.3.3 水解度的測定 根據李毅[6],采用pH-stat法加以適當修改,進行蝦夷扇貝生殖腺酶解物水解度的測定,計算公式如下:
式中:DH-水解度,%;B-保持溶液pH不變時所消耗的NaOH的體積,mL;Nb-堿的當量濃度,0.5 mmol/L;Mp-底物中蛋白質總量,g;htot-底物蛋白中肽鍵總數,mmol/g,本研究中htot以7.5計;α-水解過程中α-氨基的解離度,計算公式如下:

式中:pH-水解度測定過程中,酶解液體系恒定酸堿度;pK-本研究中pK以7.5計。
1.3.4 SDS-PAGE檢測 以不同酶解時間(15~180 min)的樣品為待測樣,以250 mmol/L pH7.5 Tris-HCl緩沖液+8 mol/L尿素+5% SDS+5%巰基乙醇為上樣緩沖液,將二者以1∶1 (v/v)的比例混合,振蕩搖勻,利用沸水浴加熱5 min,在振蕩條件下過夜靜置,次日,在4 ℃下,以12000×g轉速離心10 min,取離心后上清液進行SDS-PAGE垂直電泳檢測。實驗中,濃縮膠和分離膠的濃度分別為5%、15%,電流分別為8、15 mA,以SDS-Tris-甘氨酸體系作為電泳緩沖液。電泳完畢后,利用考馬斯亮藍R-250對實驗樣品進行染色,之后利用50%乙醇+9%冰乙酸脫色液進行脫色,利用凝膠成像儀對所得電泳膠片進行成像分析。
1.3.5 流變特性的測定 以利用木瓜蛋白酶(3000 U/g蛋白)酶解30、180 min以及經DNase I孵育后的蝦夷扇貝生殖腺酶解液為研究對象,將各酶解液100 ℃滅活后,在4 ℃內靜置過夜,采用流變儀進行酶解液的流變特性測定。
流變測定的基本測定參數為:夾具采用40 mm的平行板,Gap值為1 mm,溫度控制為15 ℃,掃描模式包括應變掃描(Oscillation Amplitude)、頻率掃描(Oscillation Frequency)和剪切速率掃描(Flow Sweep)。應變掃描范圍為0.1%~1000%,固定頻率為1 Hz;頻率掃描范圍為0.1~10 Hz,固定應變為2%;剪切速率掃描范圍為0.1~100 s-1。
1.4數據處理
采用SPSS 13.0軟件進行顯著性分析,p<0.05時具有顯著性差異;采用Microsoft Excel工作表軟件進行作圖。
2 結果與討論
2.1蝦夷扇貝雄性生殖腺酶解過程中蛋白質酶解情況
利用木瓜蛋白酶對蝦夷扇貝雄性生殖腺進行酶解,當酶解15、30、60、120、180 min時,其水解度可分別可達到8.58%、10.24%、12.32%、12.99%以及13.64%(數據未顯示),說明蝦夷扇貝雄性生殖腺在不斷被木瓜蛋白酶降解,120 min時,其降解速率減緩,之后趨于穩定。采用SDS-PAGE分析蝦夷扇貝雄性生殖腺在酶解過程中蛋白質的降解情況,如圖1所示,蝦夷扇貝雄性生殖腺中含有多種不同分子量的蛋白質,在14.3~200 kDa 之間均有不同程度的分布,其主要組分集中14.3~20.1 kDa以及44.3 kDa 左右。隨酶解時間延長,不同分子量的蛋白質均顯示出不同程度的降解,在酶解時間為15 min時,電泳條帶與未酶解樣品條帶相比,顯著變淺,且20.1kDa以上組分酶解較明顯。當達到180 min時,蝦夷扇貝雄性生殖腺中蛋白質組分全部降解,產物主要集中在14.3 kDa以下。上述結果說明隨著酶解時間的延長,蝦夷扇貝生殖腺中大分子蛋白質逐漸降解,由大分子蛋白質轉變為小分子的肽組分。
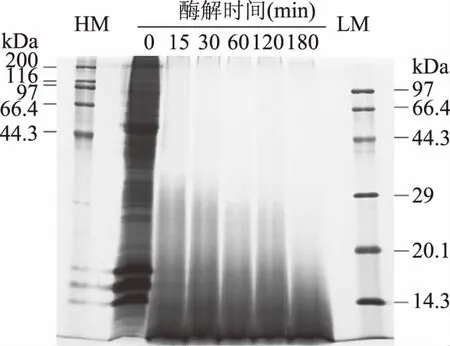
圖1 蝦夷扇貝雄性生殖腺酶解過程中蛋白質的酶解情況Fig.1 SDS-PAGE pattern of SMGHs注:HM代表高分子量Marker,LM代表低分子量Marker。
2.2蝦夷扇貝雄性生殖腺酶解物流變特性的應變掃描
應變掃描是在恒定的頻率和溫度下,給材料施加一定范圍的交變應變,測量聚合物黏彈響應隨應變變化的關系,其中G′為彈性模量,代表流體彈性行為;G″為黏性模量,代表流體黏性行為,二者相平行的區間為流體線性黏彈性區(LVR),其余區間為非線性黏彈性區(n-LVR),區間中間點所對應的應變值可作為頻率掃描的固定參數[18-19]。如圖2所示,蝦夷扇貝雄性生殖腺未酶解的懸濁液(0 min),隨著應變值增加,G′和G″均保持不變,響應值很小,反應出此時的樣品凝膠性極差或者不存在凝膠特性。當酶解時間達到30 min和180 min時,隨著應變量的增加,G′和G″起初均保持不變且相互平行,當應變達到2%后,G′開始急劇減小,與此同時,G″急劇增加之后呈下降趨勢,直到應變達到60%左右,G′均大于G″,表明此階段的酶解物表現為半固態行為,流動性相對較弱,呈現出一定的凝膠特性,當應變進一步提高,G′和G″二者出現交點,黏性特性開始高于彈性特性,G″占主導,此時的凝膠被逐漸破壞,流動性逐漸增強,酶解物顯示出流體性質。此外,隨著酶解時間增加G′、G″也相應增加,在180 min,應變為1%時,G′、G″分別達到了188.9 Pa和17.9 Pa,說明酶解物的彈性和黏性隨酶解程度的增加而增強。
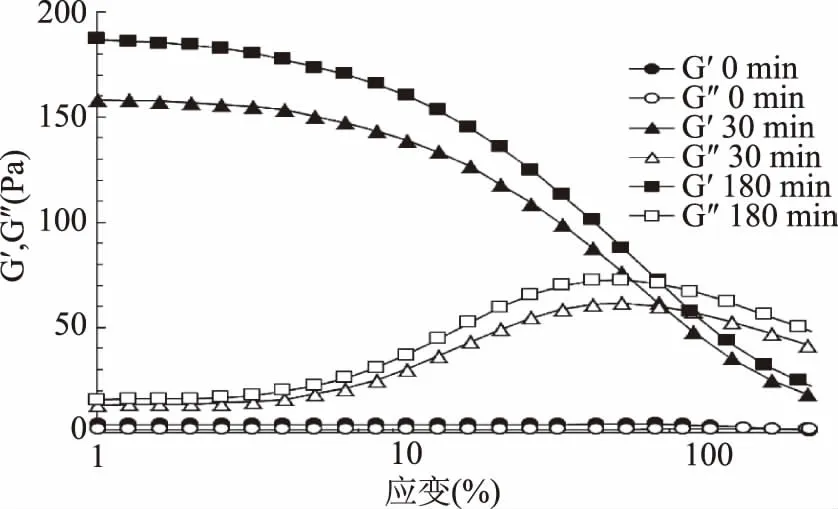
圖2 蝦夷扇貝雄性生殖腺酶解物的應變掃描Fig.2 Oscillation strain scanning of SMGHs
在未進行酶解時(0 min),蝦夷扇貝雄性生殖腺凍干粉溶液呈懸濁液狀態,無凝膠性。在木瓜蛋白酶(3000 U/g 蛋白)酶解后,經滅酶處理,冷卻放置過夜后,樣品呈現凝膠狀,具有流變性。因此,以30 min和180 min酶解所得樣品為代表,進一步利用流變儀考察蝦夷扇貝雄性生殖腺酶解物的流變特性。
2.3蝦夷扇貝雄性生殖腺酶解物流變特性的頻率掃描
頻率掃描是在固定應變下,對材料施加正弦頻率的交變運動,測試材料黏彈性與頻率變化的關系[18]。如圖3A所示,在頻率為0.1~10 Hz的范圍內,蝦夷扇貝雄性生殖腺酶解液均體現出典型的黏彈性固體流變曲線,即G′始終大于G″,進一步表明,蝦夷扇貝雄性生殖腺酶解物的彈性占主導。另外,隨著酶解時間延長,G′和G″相應增加,說明酶解促進了蝦夷扇貝雄性生殖腺酶解物的黏彈特性。
復數黏度η*體現了黏度和彈性共同的貢獻,可作為表征聚合物流體的黏彈性質的一個重要指標。如圖3B所示,一方面,隨著酶解時間增加,復數黏度η*值增加,這與圖2和圖3A的結果趨勢相一致,在180 min,頻率為0.1 Hz時,η*可達到230.9 Pa·s;另一方面,復數黏度η*隨頻率增加急劇下降,說明蝦夷扇貝雄性生殖腺酶解物在不斷增加的頻率剪切下,呈現出明顯的剪切變稀現象,體現出弱凝膠性質。
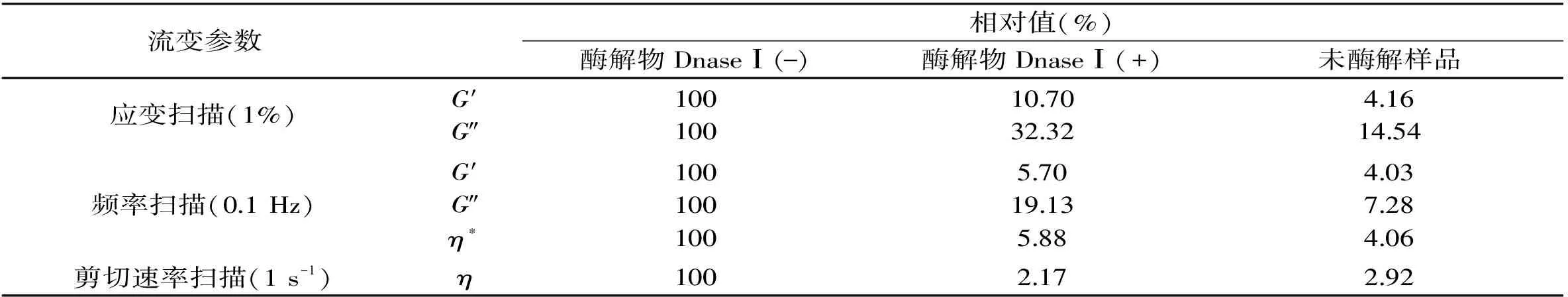
表1 DNaseⅠ對蝦夷扇貝雄性生殖腺酶解物流變特性的影響相對值Table 1 Relative values for the effect of DNaseⅠon hydrolysates from SMGs
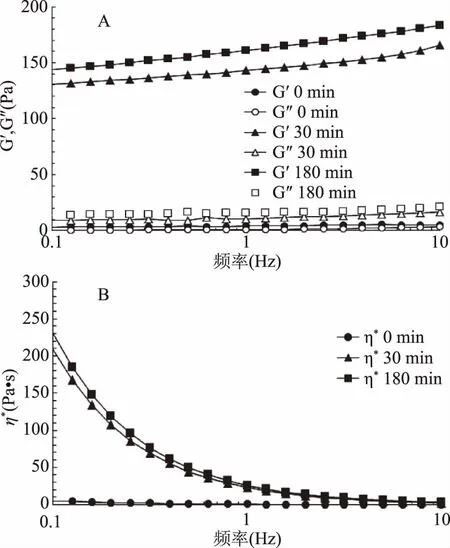
圖3 蝦夷扇貝雄性生殖腺酶解物的頻率掃描Fig.3 Frequency scanning of SMGHs
注:酶解物DnaseⅠ(-)代表未經DNase I處理的樣品,酶解物DnaseⅠ(+)表示經過DNase I處理的樣品。
2.4蝦夷扇貝雄性生殖腺酶解物流變特性的剪切速率掃描
剪切速率掃描即在規定的剪切速率范圍內,對流體進行剪切掃描,可獲得黏度、剪切應力等指標的變化規律,之后以剪切應力作為剪切速率的函數作圖,將得到的曲線進行一定方程擬合可得出屈服應力,此方法可以真實反映樣品性質,從而得到最廣泛應用[20]。如圖4所示,在規定的剪切速率范圍內(0.1~5 s-1),隨著剪切速率的增加,蝦夷扇貝雄性生殖腺酶解物的黏度急劇減少;直到5 s-1時,酶解物黏度的減小趨勢變緩,趨于穩定,呈現出明顯的剪切稀化現象。另外,在剪切速率掃描模式下,酶解30、180 min的蝦夷扇貝酶解物樣品的黏度相差不大,考慮原因可能為樣品的特殊性,呈現弱凝膠特性,故在剪切速率掃描時,30 min和180 min的樣品的黏度變化相差不大。但二者的黏度均明顯高于未酶解樣品。
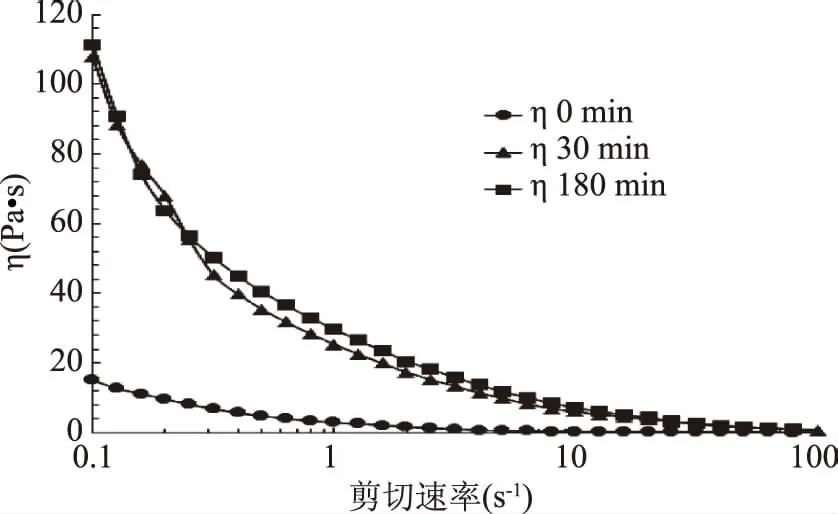
圖4 蝦夷扇貝雄性生殖腺酶解物剪切速率掃描Fig.4 Shear rate scanning of SMGHs
2.5DNA對蝦夷扇貝雄性生殖腺酶解物流變特性的影響
DNA降解酶(DNase I)作為一種典型的DNA降解酶,可有效降解樣品中DNA組分。為了進一步探索在蝦夷扇貝雄性生殖腺酶解物凝膠形成時,DNA分子的作用貢獻,通過前期利用DNase I酶對原料進行孵育,之后測定孵育前后酶解物在不同應變、頻率和剪切速率下流變特性的變化,結果見表1所示。與未添加DNase I的酶解物相比,經DNase I處理后的酶解液的流變特性發生顯著變化。在應變和頻率掃描中,DNase I可明顯降低蝦夷扇貝雄性生殖腺酶解物的G′、G″以及η*,但略高于未酶解樣品。在剪切速率掃描中,DNase I可明顯降低蝦夷扇貝雄性生殖腺酶解物的η,且略低于未酶解樣品。以上結果表明,經過DnaseⅠ處理后,蝦夷扇貝雄性生殖腺凝膠狀酶解物的凝膠特性被不同程度的破壞,呈現出液體狀態。近年來,有關于DNA分子與肽分子的相互作用得到廣泛研究[21-26]。Bitton等[21]研究發現,GCN4(KDPAALKRAR NTEAARRSRA RKLQRMKQLE)多肽與DNA分子間通過靜電引力相互作用。Gour等[22]利用寡核苷酸與二苯基二肽形成共軛體系,制備DNA-肽自組裝體,經實驗發現,新體系呈囊泡狀,可作為一種潛在的包埋材料。Kye等[23]建立了超分子結構的肽-DNA共軛體系,螺旋狀結構的DNA與β-折疊狀結構的肽通過靜電引力相互結合,具有穩定的結構。在本次研究中,由于加入了DNA降解酶DNase I的緣故,使樣品內部DNA分子遭到一定程度的破壞,進而導致DNA分子無法與樣品中肽組分進行結合,使樣品的凝膠特性被破壞。綜上所述,在蝦夷扇貝雄性生殖腺酶解成膠過程中,DNA分子可能起到了關鍵性作用,而關于其具體作用機制以及成膠機理,還需進一步探究。
3 結論
經過木瓜蛋白酶處理后的蝦夷扇貝雄性生殖腺酶解物可體現出較好的流變特性,其儲存模量G′、損耗模量G″、黏度η和復數黏度η*較未酶解樣品相比均顯著提高。蝦夷扇貝雄性生殖腺凝膠狀酶解物的流變特性以彈性占主導,具有明顯的剪切稀化現象。蝦夷扇貝雄性生殖腺凝膠狀酶解物的形成可能與酶解后的小分子肽以及DNA分子有關。
[1]侯團偉,張虹,畢艷蘭,等. 食品膠體的凝膠機理及協同作用研究進展[J]. 食品科學,2014,35(23):347-353.
[2]黃來發. 食品增稠劑[M]. 北京:中國輕工業出版社,2000:1-12.
[3]農業部漁業局. 2016中國漁業統計年鑒[M].北京:中國農業出版社,2016:28-29.
[4]Jin WG,Wu HT,Zhu BW,et al. Functional properties of gelation-like protein hydrolysates from scallop(Patinopectenyessoensis)male gonad[J]. European Food Research and Technology,2012,234(5):863-872.
[5]Jin WG,Wu HT,Li XS,et al. Microstructure and intermolecular forces involved in gelation-like protein hydrolysate from neutrase-treated male gonad of scallop(Patinopectenyessoensis)[J]. Food Hydrocolloids,2014,40(1):245-253.
[6]李毅,張萌,陳庭家,等. 蝦夷扇貝生殖腺酶解液的凝膠特性[J]. 中國食品學報,2015,15(4):22-29.
[7]Zhong F,Wang Z,Xu SY,Shoemaker CF. The evaluation of proteases as coagulants for soy protein dispersions[J]. Food Chemistry,2007,100(1):1371-1376.
[8]Lv Y,Guo ST,Yang BC. Aggregation of hydrophobic soybean protein hydrolysates:Changes in molecular weight distribution during storage[J]. LWT-Food Science and Technology,2009,42(1):914-917.
[9]Pouliot Y,Guy MM,Tremblay M,et al. Isolation and characterization of an aggregating peptide from a tryptichydrolysate of whey proteins[J].Agricultural and Food Chemistry,2009,57(1):3760-3764.
[10]Spellman D,Kenny P,O’Cuinn G,et al. Aggregation properties of whey protein hydrolysates generated withBacilluslicheniformisproteinase activitie[J]. Agricultural and Food Chemistry,2005,53(4):1258-1265.
[11]Orcajo J,Marcet I,Paredes B,et al. Egg yolk hydrolysed granules:characteristics,rheological properties and applications[J]. Food and Bioproducts Processing,2015,91(91):457-463.
[12]Rao M A,Andy. Rheology of fluid and semisolid foods:principles and applications[J]. Food Engineering,2013,12(1):124-128.
[13]Zhang H,Yin L,Zheng Y,et al. Rheological,textural,and enzymatic hydrolysis properties of chickpea starch from a Chinese cultivar[J]. Food Hydrocolloids,2015,54(1):23-29.
[14]Derkach S R,Ilyin S O,Maklakova A A,et al. The rheology of gelatin hydrogels modified by κ-carrageenan[J]. LWT-Food Science and Technology,2015,63(1):612-619.
[15]蔡路昀,馮建慧,聶小華,等. 草魚皮明膠-可得然膠共混體系流變特性及作用機理的研究[J]. 食品工業科技,2017(1):49-53.
[16]Szwajgier D,Gustaw W. The addition of malt to milk-based desserts:Influence on rheological properties and phenolic acid content[J]. LWT-Food Science and Technology,2015,62(1):400-407.
[17]Derkach S R,Ilyin S O,Maklakova A A,et al. The rheology of gelatin hydrogels modified by κ-carrageenan[J]. LWT-Food Science and Technology,2015,63(1):612-619.
[18]辜婷,朱大勇,劉典新等. 旋轉流變儀及其在塑料中的研究應用[J]. 塑料工業,2017,45(2):97-100.
[19]Ahmed J,Ramaswamy H S,Sashidhar K C. Rheological characteristics of tamarind(TamarindusindicaL.)juice concentrates[J]. LWT-Food Science and Technology,2007,40(2):225-231.
[20]張智明,張虹,畢艷蘭,等. 旋轉流變儀在油脂研究中的應用[J]. 中國油脂,2013,38(9):1-6.
[21]Bitton R,Schmidt J,Biesalski M,et al. Self-assembly of model DNA-binding peptide amphiphiles[J]. Langmuir,2005,21(25):11888-11895.
[22]Gour N,Kedracki D,Safir I,et al. Self-assembling DNA-peptide hybrids:morphological consequences of oligonucleotide grafting to a pathogenic amyloid fibrils forming dipeptide[J]. Chemical Communications,2012,48(44):5440-5442.
[23]Kye M,Lim Y B. Reciprocal Self-Assembly of Peptide-DNA Conjugates into a Programmable Sub-10-nm Supramolecular Deoxyribonucleoprotein[J]. Angewandte Chemie,2016,55(39):12003.
[24]Morán M C,Miguel M G,Lindman B. Surfactant-DNA gel particles:Formation and release characteristics[J]. Biomacromolecules,2007,8(1):3886-3892.
[25]Thompson M. DNA sequence-directed assembly of two peptide bioconjugates[J]. Bioorganic & Medicinal Chemistry,2006,34(5):235-247.
[26]Morán M C,Baptista F R,Ramalho A,et al. DNA gel nanoparticles:preparation and controlling the size[J]. Soft Matter,2009,5:2538-2542.
Rheologicalpropertiesofproteinhydrolysateswithgelpropertiesfromscallop(Patinopectenyessoensis)malegonads
YANJia-nan1,LIYi2,TANGYue1,ZHANGMeng1,SHANGWen-hui1,DUYi-nan1,JIANGHui1,WUHai-tao1,*
(1.School of Food Science and Technology,Dalian Polytechnic University,National Engineering Research Center of Seafood,Dalian 116034,China; 2.Inspection and Quarantine Technology Center,Hainan Entry-Exit Inspection and Quarantine Bureau,Haikou 570311,China)
In order to provide a foundation for comprehensive utilization of scallop(Patinopectenyessoensis),the rheological properties of hydrolysates from scallop(Patinopectenyessoensis)gonad(SMGHs)were studied. The hydrolysates were obtained from scallop male gonads(SMGs)by using papain as a tool enzyme. The changes in molecular mass of proteins were investigated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis(SDS-PAGE). The rheological properties of SMGHs were determined by rheometer. The effect of DNA on rheological properties of SMGHs was also investigated by incubated with DNase Ⅰ. The results showed that the proteins of SMGs were significantly degraded after hydrolysis with papain at a dosage of 30000 U/g protein for 15~180 min. Under various sweep models,the values of storage modulusG′,loss modulusG″,viscosityη,complex viscosityη*of SMGHs were obvious higher than those of SMGs. After hydrolysis for 3 h,the initial values ofG′,G″ during oscillation strain scanning were up to 188.9,17.9 Pa at oscillation strain of 1%,respectively. The value ofη*was 230.9 Pa·s at frequency of 0.1 Hz. The rheological properties(G′,G″η*)of SMGHs decreased significantly,but slightly higher than those of SMGs,by treatment with DNaseⅠ. These results suggest that the elastic property is dominant in SMGHs exhibiting obvious shear thinning effect. Moreover,the rheological properties of SMGHs might be related to peptides and DNA molecule.
scallop(Patinopectenyessoensis);male gonads hydrolysates;gel;rheological properties
2017-05-02
閻佳楠(1993-),女,碩士研究生,研究方向:食品生物技術,E-mail:1070378773@qq.com。
*
吳海濤(1980-),女,博士,副教授,研究方向:食品生物技術,E-mail:wht205@163.com
國家自然科學基金項目(31671808)。
TS254
A
1002-0306(2017)21-0047-05
10.13386/j.issn1002-0306.2017.21.010
