谷氨酸和肽聚糖在促進細菌間納米管形成中作用的研究
2017-11-21 03:19:24鐘橋石杭亞平劉衍伶陳艷慧胡龍華
中國人獸共患病學報 2017年10期
鐘橋石,杭亞平,劉衍伶,丁 慧,俞 鳳,陳艷慧,胡龍華
谷氨酸和肽聚糖在促進細菌間納米管形成中作用的研究
鐘橋石1,杭亞平1,劉衍伶2,丁 慧1,俞 鳳1,陳艷慧1,胡龍華1
目的對細菌間納米管形成機制及組成成分進行探索,為預防細菌耐藥提供潛在靶標。方法利用掃描電子顯微鏡觀察細菌間納米管的形成,用表面解吸常壓化學電離質譜(SDAPCI-MS)技術和酶聯免疫吸附實驗(ELISA)分別檢測細菌間納米管形成前后谷氨酸(glutamic,Glu)和肽聚糖(peptidoglycan,PGN)的濃度。結果相同菌量的金黃色葡萄球菌和大腸埃希菌在納米管形成后PGN含量明顯增多,且它們在納米管形成后消耗了更多的Glu。結論PGN是納米管組成成分,Glu是促進細菌間納米管形成的主要能量來源。
納米管;谷氨酸;肽聚糖
由于抗菌藥物在臨床的廣泛使用,臨床常見細菌在抗菌藥物的選擇壓力下產生耐藥性,過往對細菌耐藥機制關注最多的是水解酶[1-2]的產生、藥物作用靶位點及細菌藥物外排泵的改變[3]等;尤其是細菌還可以通過獲得耐藥基因的方式,形成多重耐藥甚至泛耐藥菌,極大的增加了臨床用藥選擇壓力。然而,最新研究發現細菌間可以形成納米管(Nanotubes)來傳遞耐藥基因[4]。而納米管的物質組成成分及形成機制尚不完全清楚。本文將對細菌間納米管形成機制及組成成分進行探索,旨在為預防細菌耐藥提供潛在靶標。
1 材料與方法
1.1菌株 金黃色葡萄球菌ATCC25923和大腸埃希菌ATTCC25922。
1.2主要試劑與儀器 VITEK 2-Compact全自動細菌分析儀(法國)及配套鑒定卡,Bio-merieux公司產品;掃描電子顯微鏡(SEM Quanta200),美國FEI公司生產;LTQ-XL線性離子阱質譜儀,美國菲尼根公司產品;表面解吸常壓化學電離源(SDAPCI),為陳煥文教授[5]自主研發;哥倫比亞血瓊脂培養基,法國生物梅里埃公司產品。
1.3細菌培養及鑒定 將菌株轉種至血瓊脂平板分離細菌,依據血瓊脂平板上的生長情況,獲得純培養后,經革蘭染色、凝固酶、觸酶、氧化酶試驗等初篩,再用GPI或GNI卡在VITEK 2-Compact分析儀上進行菌株鑒定,嚴格按儀器操作規程進行操作。
1.4電鏡觀察掃描電子顯微鏡樣品制備 將含有OD600=0.05的菌液的金黃色葡萄球菌和大腸埃希菌分別在25 mL LB固體、LB液體培養基上37 ℃孵育3 h后,把碳支持膜貼到菌膜或菌液中,在37 ℃繼續孵育3 h;取出碳支持膜用0.1M PBS清洗后,用2%戊二醛固定2 h。用25%、50%、75%和95%濃度的酒精各脫水10 min,最后用無水乙醇脫水20 min;待測標本鍍金處理后在掃描電子顯微鏡觀察。
1.5離子濃度測定 SDAPCI-MS檢測細菌間納米管形成前后谷氨酸的相對離子濃度
1.5.1檢測條件 用正、負離子2種檢測模式,將SDAPCI離子源耦合到LTQ線性離子阱質譜儀中,檢測掃描m/z 20-200,LTQ-Tune功能自動優化離子透鏡、檢測系統及其他參數。以空白平板為對照。
1.5.2檢測方法 將經培養后培養基置于密閉管中,空白對照血瓊脂平板隨機取樣。充入氮氣的空氣樣品作為反應氣體用SDAPCI-MS檢測,手動方式進樣,待氣流平穩、峰值穩定,每個樣品持續進樣30~60 s。按照培養時間由短到長的順序,空白對照與樣品交替分析,每個獨立樣品分別測定3次。
1.6酶聯免疫吸附實驗(ELISA)檢測細菌間納米管形成前后肽聚糖的濃度
1.6.1菌株篩選 金黃色葡萄球菌ATCC25923和大腸埃希菌ATTCC25922作為研究菌株,選擇的標準分別為體外納米管形成陽性和陰性菌;分別將含有OD600=0.05的菌液金黃色葡萄球菌在25 mL LB固體、液體培養基上37 ℃孵育3 h后,把碳支持膜貼到菌膜中,在37 ℃繼續孵育3 h,電鏡確認金黃色葡萄球菌間和大腸埃希菌間納米管形成情況。
1.6.2檢測方法 分別將含有OD600=0.05的菌液的納米管形成前后的金黃色葡萄球菌和大腸埃希菌用PBS(PH7.2~7.4)稀釋細胞懸液,通過裂解液試劑,以使細菌破壞并放出細菌內成份。離心20 min(2000~3000 r/min)。收集上清。保存過程中如有沉淀形成,應再次離心;依據肽聚糖ELISA法分析使用說明書檢測納米管形成前后金黃色葡萄球菌和大腸埃希菌肽聚糖的含量。
2 結 果
2.1掃描電子顯微鏡結果 經掃描電子顯微鏡分別觀察到金黃色葡萄球菌間和大腸埃希菌間形成的納米管(見圖1)。
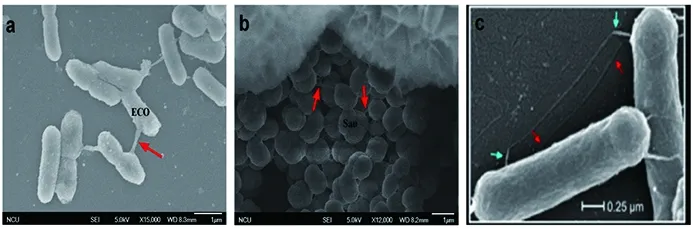
注:a:大腸埃希菌菌株間形成的納米管 b:金黃色葡萄球菌菌株間形成的納米管 c:攜帶有pLS20質粒的枯草芽孢桿菌(GD127)間的菌毛接合(×70 000 Gyanendra P et al.2011)。Note:a, formation of nanotubes in E.coli;b, formation of nanotubes in staphylococcus aureus;c, pilus conjunction in carry pLS20 plasmid of bacillus subtilis (GD127)圖1 掃描電鏡下各菌株間形成的納米管Fig.1 Nanotubes formed in various spaces by scanning electron microscopy
2.2質譜檢測 經SDAPCI-MS檢測得到其20-200范圍內離子流譜圖,通過進樣時間篩選出需要的進樣峰(64,空氣中幾乎檢測不到)。發現148(谷氨酸)高強度地出現在無納米管形成的菌株中,納米管形成后強度明顯下降,與金黃色葡萄球菌相比較,大腸埃希菌在納米管形成過程中消耗了更多的谷氨酸(見圖2)。金黃色葡萄球菌和大腸埃希菌在納米管形成后,谷氨酸的相對離子強度明顯低于無納米管形成時的相對離子強度(見圖3)。
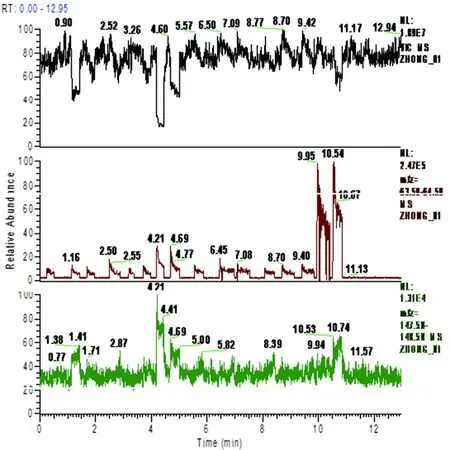
注:縱軸為相對離子強度,橫軸為進樣時間;黑線為離子流譜圖;紅線為進樣峰,綠色為148信號峰(為Glu)Y-axis is relative abundance, X-axis for sample injection time.Black line is spectra of ion flow;The red line is sample injection peak, green for 148 signal peak (Glu)圖2 sau和eco菌納米管形成前后Glu質譜圖Fig.2 The amass spectrum of glutamate in E. coli and Staphylococcus aureus in the formation of nanotubes before and after
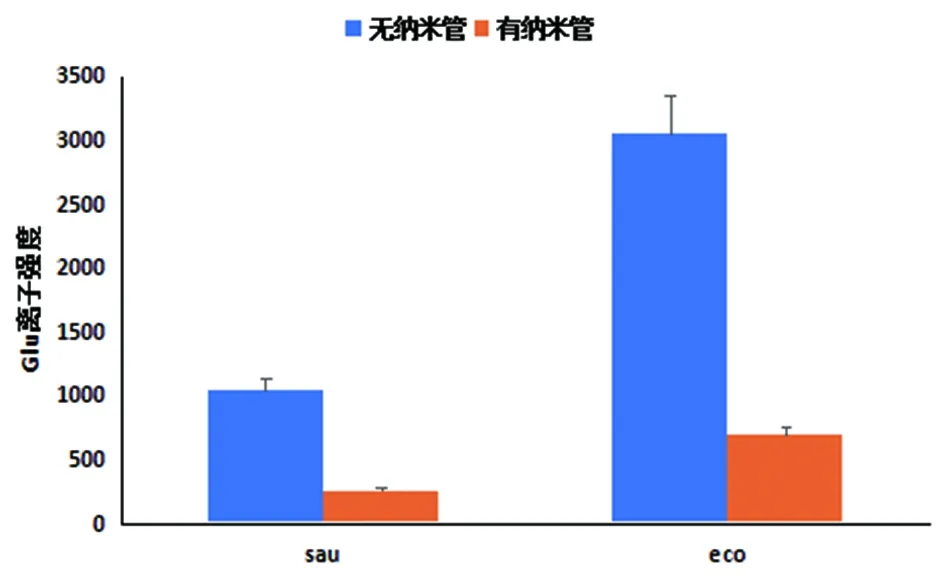
注:sau為金黃色葡萄球菌納米管形成前后,上層氣體中Glu平均離子強度;eco為大腸埃希菌納米管形成前后,上層氣體中Glu平均離子強度sau is average ion strength of glutamate of upper gas in staphylococcus aureus in the formation of nanotubes before and after; eco is average ion strength of glutamate of upper gas in E. coli in the formation of nanotubes before and after圖3 sau和eco菌納米管形成前后Glu平均離子強度Fig.3 The average ion strength of glutamate in E. coli and Staphylococcus aureus in the formation of nanotubes before and after
2.3ELISA檢測結果 經ELISA檢測待測菌株在納米管形成前后肽聚糖含量,我們發現相同菌量的金黃色葡萄球菌和大腸埃希菌在納米管形成后PGN含量明顯增多(見圖4)。
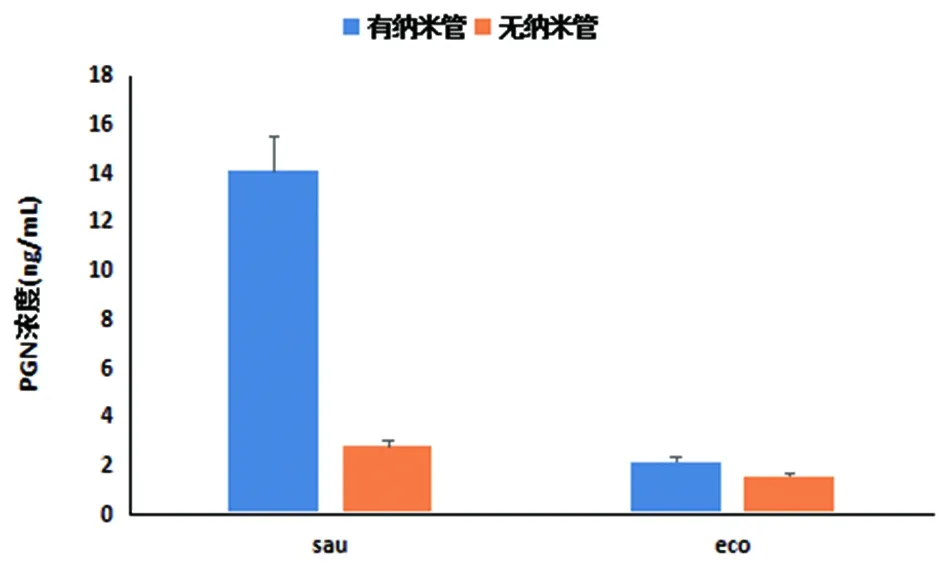
注:sau為金黃色葡萄球菌納米管形成前后PGN的平均濃度;eco為大腸埃希菌納米管形成前后PGN的平均濃度sau is average concentration of polysaccharides in staphylococcus aureus in the formation of nanotubes before and after;eco is average concentration of polysaccharides in E.coli in the formation of nanotubes before and after the formation圖4 sau與eco菌納米管形成前后PGN平均濃度Fig.4 The average concentration of polysaccharides in E.coli and Staphylococcus aureus in the formation of nanotubes before and after the formation
3 討 論
隨著廣譜抗菌藥物在臨床的廣泛使用,細菌的耐藥性呈逐年上升趨勢,臨床分離的多重耐藥或泛耐藥的細菌也越來越多,對此類細菌的治療給臨床帶來極大的困難。近年來,通過抑制耐藥基因的傳遞來控制耐藥菌的產生受到越來越多的關注。
2010年Gyanendra P等研究發現[4],枯草芽孢桿菌和大腸埃希菌在固體培養基上能形成類似管道的結構(納米管),且進一步證實不同菌屬之間也能形成納米管,而在液體培養基中不能形成,細菌間這種全新的通信方式——相鄰細菌間的納米管形成受到關注。在電鏡中,納米管的結構與細胞膜類似,長可達1 μm,徑寬30~130 nm。該研究證實,納米管能有效、快速地在細菌間進行胞質小分子、耐藥基因及質粒的傳遞,尤其是在進行質粒傳遞時,其效率遠高于質粒接合,且其轉移的速度與納米管內徑寬度大小成正相關。由于納米管能夠快速的傳遞耐藥基因,因此針對開發抑制納米管形成來預防耐藥菌的快速增加的研究具有重要意義。
研究顯示納米管內部有多層結構,而具體組成成分及其形成機制尚不完全清楚,但有研究表明,Glu能通過調節生物膜內部與外圍細菌能量供給的方式,使生物膜的形成產生類似集體振蕩(collective oscillations,生物膜增長間歇性停止)的方式,從而提高生物膜抵御來自外部化學侵蝕的能力[6]。細菌生物膜除了含有水和細菌外,還含有蛋白質、多糖、PGN等物質[7]。PGN是細菌等原核生物細胞壁的特有成分,其對提高細胞壁的機械強度和維持細胞形狀起著重要作用[8]。Glu和PGN在生物膜形成中具有重要作用,而納米管形成的條件與細菌生物膜形成的條件又有眾多相似之處,PGN和Glu在納米管的形成中是否也具有重要作用,國內外還尚無研究報道。我們研究發現相同菌量的金黃色葡萄球菌和大腸埃希菌在納米管形成后PGN含量明顯增多(見圖4),提示PGN是納米管組成成分;通過SDAPCI-MS檢測還發現它們在納米管形成后將消耗更多的Glu(見圖2-3);研究證實,通過納米管進行物質交換并不消耗能量,由此表明Glu是促進細菌間納米管形成的主要能量來源,Glu和PGN在納米管形成中是否相互關聯有待進一步研究。
隨著細菌耐藥性的不斷上升,越來越多的研究關注細菌間耐藥基因的傳遞機制,從而控制耐藥菌的產生。我們研究首次發現Glu和PGN在促進細菌間納米管的形成中具有重要作用,為有效干預細菌獲得耐藥基因提供新的思路,為通過抑制納米管形成防止耐藥基因在菌株間傳遞提供潛在靶標。
[1] Koch G, Yepes A, Forstner KU, et al. Evolution of resistance to a last-resort antibiotic in Staphylococcus aureus via bacterial competition[J]. Cell, 2014,158(5): 1060-1071. DOI: 10.1016/j.cell.2014.06.046.
[2] Nijhuis RH, Oueslati S, Zhou K, et al. OXY-2-15, a novel variant showing increased ceftazidime hydrolytic activity[J]. J Antimicrob Chemother , 2015,70(5): 1429-1433. DOI: 10.1093/jac/dkv002.
[3] Kim CH, Kang HY, Kim BR, et al. ,Mutational inactivation of OprD in carbapenem-resistantPseudomonasaeruginosaisolates from Korean hospitals[J]. J Microbiol ,2016,54(1): 44-49. DOI: 110.1007/s12275-016-5562-5. Epub 2016 Jan 5.
[4] Dubey GP, Ben-Yehuda S. Intercellular nanotubes mediate bacterial communication[J]. Cell, 2011, 144(4): 590-600. DOI: 10.1016/j.cell.2011.01.015
[5] Chen H, Luo M, Xiao S, et al. Extractive electrospray ionization mass spectrometry for uranium chemistry studies [J]. Mass Spectrom (Tokyo),2013, 2(9): S0021.DOI:10.5702/massspectrometry.S0021.
[6] Liu J, Prindle A, Humphries J, et al. Metabolic co-dependence gives rise to collective oscillations within biofilms [J]. Nature, 2015, 523(7562): 550-554. DOI: 10.1038/nature14660.
[7] Denoel T, Zervosen A, Lemaire C, et al. Enantioselective synthesis of alpha-benzylated lanthionines and related tripeptides for biological incorporation intoE.colipeptidoglycan [J]. Org Biomol Chem, 2014, 12(48): 9853-9863. DOI: 10.1039/c4ob01476f.
[8] McGlynn SE, Chadwick GL, Kempes CP, et al. Single cell activity reveals direct electron transfer in methanotrophic consortia [J]. Nature, 2015, 526(7574): 531-535. DOI: 10.1038/nature15512.
Theroleofglutamateandpeptidoglycaninnanotubesformationbetweenadjacentbacteria
ZHONG Qiao-shi1,HANG Ya-ping1,LIU Yan-lin2,DING Hui1,YU Feng1,CHEN Yan-hui1,HU Long-hua1
(1.DepartmentofLaboratoryMedicine,theSecondAffiliatedHospitalofNanchangUniversity,Nanchang330006,China;2.DepartmentofLaboratoryMedicine,theFirstAffiliatedHospitalofNanchangUniversity,Nanchang330006,China)
We investigated the formation mechanism and immediate constituents of nanotubes between adjacent bacteria,preventing rapid increase of drug-resistant bacteria provide potential targets.We used scanning electron microscopy(SEM) to observe formation of nanotubes;glutamic was determined by surface desorption atmospheric pressure chemical ionization mass spectrometry(SDAPCI-MS)and peptidoglycan was determined by enzyme linked immunosorbent assay(ELISA) in the formation of nanotubes before and after.The results showed thatStaphylococcusaureusandEscherichiacoliconsumed more glutamic after formation of nanotubes;at the same time, the concentration of peptidoglycan in nanotubes formation ofS.aureusandE.coliincreased significantly.This study illuminated the glutamic and peptidoglycanin role mechanism of nanotubes form.We found that peptidoglycan is constituents of nanotubes and glutamic is the main energy source for the formation of nanotubes.
nanotubes;glutamic; peptidoglycan
Hu Long-hua,Email:longhuahu@163.com
10.3969/j.issn.1002-2694.2017.10.008
江西省青年自然基金資助項目(No.20171BAB2150798)和江西省自然基金資助項目(No.20133BAB205062)聯合資助
胡龍華,Email:longhuahu@163.com
1.南昌大學第二附屬醫院檢驗科,南昌 330006;
2.南昌大學第一附屬醫院檢驗科,南昌 330006
378
A
1002-2694(2017)10-0893-04
Funded by the Youth Natural Science Foundation of Jiangxi and the Natural Science Foundation of Jiangxi(No.20133BAB205062)
2017-06-11編輯李友松
猜你喜歡
保健醫苑(2022年5期)2022-06-10 07:46:38
現代臨床醫學(2022年3期)2022-06-06 07:59:40
昆明醫科大學學報(2022年1期)2022-02-28 07:43:40
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
科學大眾(2020年12期)2020-08-13 03:22:22
海峽科技與產業(2016年3期)2016-05-17 04:32:12
