白藜蘆醇對(duì)過氧化氫誘導(dǎo)巨噬細(xì)胞炎性衰老的抑制作用及機(jī)制
2017-11-15 07:48:49蘇雙阮云軍馬懿馮騫石靈林捷琪吳賽珠
山東醫(yī)藥 2017年37期
蘇雙,阮云軍,馬懿,馮騫,石靈,林捷琪,吳賽珠
(1南方醫(yī)科大學(xué)南方醫(yī)院,廣州5140000;2遵義醫(yī)學(xué)院附屬醫(yī)院)
白藜蘆醇對(duì)過氧化氫誘導(dǎo)巨噬細(xì)胞炎性衰老的抑制作用及機(jī)制
蘇雙1,阮云軍1,馬懿2,馮騫1,石靈1,林捷琪1,吳賽珠1
(1南方醫(yī)科大學(xué)南方醫(yī)院,廣州5140000;2遵義醫(yī)學(xué)院附屬醫(yī)院)
目的觀察白藜蘆醇(Res)對(duì)過氧化氫(H2O2)誘導(dǎo)的THP-1巨噬細(xì)胞炎性衰老的抑制作用,并探討其作用機(jī)制。方法用160 nmol/L佛波酯誘導(dǎo)人單核細(xì)胞THP-1巨噬細(xì)胞,將其隨機(jī)分為四組,空白對(duì)照組給予培養(yǎng)基培養(yǎng);H2O2組給予培養(yǎng)基培養(yǎng)1 h后再加300 μmol/L H2O2培養(yǎng)24 h;Res+H2O2組給予40 μmol/L Res培養(yǎng)1 h,再加300 μmol/L H2O2培養(yǎng)24 h;Res組給予 40 μmol/L Res培養(yǎng)24 h。用衰老相關(guān)β-半乳糖苷酶染色法檢測(cè)衰老THP-1巨噬細(xì)胞比例,用Western blotting法檢測(cè)THP-1巨噬細(xì)胞內(nèi)視網(wǎng)膜細(xì)胞瘤蛋白(Rb)、磷酸化視網(wǎng)膜細(xì)胞瘤蛋白(pRb)、自噬微管相關(guān)蛋白1清鏈3(LC3)、Bcl-2同源結(jié)構(gòu)域蛋白(Beclin1)、泛素結(jié)合蛋白(p62)表達(dá);ELISA法檢測(cè)THP-1巨噬細(xì)胞培養(yǎng)液中TNF-α、IL-1β、IL-6水平。結(jié)果與空白對(duì)照組比較,H2O2組衰老細(xì)胞比例高(P<0.05);與H2O2組比較,Res+H2O2組衰老細(xì)胞比例低(P<0.05)。與空白對(duì)照組比較,H2O2組Rb蛋白表達(dá)高、p-Rb蛋白表達(dá)低(P均<0.05);與H2O2組比較,H2O2+Res組Rb蛋白表達(dá)低、p-Rb蛋白表達(dá)高(P均<0.05)。與空白對(duì)照組比較,H2O2組,LC3-Ⅱ、Beclin-1蛋白表達(dá)及LC3-Ⅱ/LC3-Ⅰ低,p62蛋白表達(dá)高(P均<0.05);與H2O2組比較,H2O2+Res組LC3-Ⅱ、Beclin-1蛋白表達(dá)及LC3-Ⅱ/LC3-Ⅰ高,p62蛋白表達(dá)低(P均<0.05)。與空白對(duì)照組比較,H2O2組TNF-α、IL-1β、IL-6水平高(P均<0.05);與H2O2組比較,H2O2+Res組TNF-α、IL-1β、IL-6水平低(P均<0.05)。空白對(duì)照組與Res組以上指標(biāo)比較,P均>0.05。結(jié)論白藜蘆醇可能通過促進(jìn)細(xì)胞自噬來抑制H2O2誘導(dǎo)的THP-1巨噬細(xì)胞炎性衰老。
炎性衰老;巨噬細(xì)胞;白藜蘆醇;細(xì)胞自噬
衰老是一個(gè)復(fù)雜的生理病理過程。在衰老過程中大量細(xì)胞和分子發(fā)生變化,加速炎性疾病的發(fā)生發(fā)展。這種現(xiàn)象在老年人中被稱作“炎性衰老”[1]。炎性衰老是指衰老進(jìn)程中機(jī)體呈現(xiàn)的低水平慢性促炎狀態(tài),其中巨噬細(xì)胞、細(xì)胞應(yīng)激和遺傳因素在炎性狀態(tài)中起了主要作用。越來越多研究表明,炎性衰老的促炎細(xì)胞對(duì)動(dòng)脈粥樣硬化(AS)的發(fā)生發(fā)展起一定作用[1,2]。白藜蘆醇(Res)是一種多酚類植物抗毒素,是作用最強(qiáng)的沉默信息調(diào)節(jié)因子2相關(guān)酶1激活劑[3],并具有廣泛的生物活性及藥理作用,能夠抵抗或延緩與衰老及氧化等相關(guān)的疾病。研究表明,Res能保護(hù)血管內(nèi)皮細(xì)胞、平滑肌細(xì)胞、心肌細(xì)胞,延緩細(xì)胞衰老,減少炎性因子的釋放,對(duì)抗AS病變[3,4]。巨噬在AS疾病發(fā)生發(fā)展中發(fā)揮重要作用[5]。2016年1月~2017年7月,本研究觀察Res對(duì)過氧化氫(H2O2)誘導(dǎo)的THP-1巨噬細(xì)胞的衰老及炎癥的干預(yù)作用,并探討其可能的作用機(jī)制。
1 材料與方法
1.1 材料來源 人單核細(xì)胞株THP-1購(gòu)自中國(guó)科學(xué)院;RPMI1640細(xì)胞培養(yǎng)基、胎牛血清培養(yǎng)基購(gòu)自Gibco;Res、佛波酯(PMA)、3-甲基腺嘌呤購(gòu)自Sigma;細(xì)胞衰老β-半乳糖苷酶染色試劑盒、BCA檢測(cè)蛋白試劑盒購(gòu)自北京碧云天;巨噬細(xì)胞內(nèi)視網(wǎng)膜細(xì)胞瘤蛋白(Rb)、磷酸化視網(wǎng)膜細(xì)胞瘤蛋白(pRb)、自噬微管相關(guān)蛋白1清鏈3(LC3)、p62/SQSTMI、Bcl-2同源結(jié)構(gòu)域蛋白(Beclin1)、泛素結(jié)合蛋白(p62)、β-actin抗體均購(gòu)自美國(guó)CST公司;羊抗兔二抗購(gòu)自杭州弗德生物科技有限公司;ECL發(fā)光液購(gòu)自Thermo公司;TNF-α、IL-1β、IL-6的ELISA試劑盒購(gòu)自深圳晶美生物有限公司。
1.2 細(xì)胞培養(yǎng)、誘導(dǎo)及分組 THP-1細(xì)胞復(fù)蘇后用含10%胎牛血清的RPMI1640培養(yǎng)基,于37 ℃、5% CO2培養(yǎng)箱中靜置培養(yǎng),待細(xì)胞長(zhǎng)至80%~90%時(shí)行下一步實(shí)驗(yàn)。用含160 nmol/L PMA及10%胎牛血清的RPMI1640細(xì)胞培養(yǎng)基刺激36 h,誘導(dǎo)為THP-1巨噬細(xì)胞。細(xì)胞接種于96孔培養(yǎng)板(1×104/孔),每孔加含160 nmol/L PMA的培養(yǎng)液100 μL,置于含5% CO2培養(yǎng)箱中誘導(dǎo)貼壁36 h。棄培養(yǎng)液,每孔加入培養(yǎng)液100 μL。根據(jù)預(yù)實(shí)驗(yàn)獲得H2O2的最佳刺激濃度為300 μmol/L、最佳時(shí)間為24 h,Res分別為40 μmol/L、24 h。隨機(jī)將細(xì)胞分為四組,空白對(duì)照組給予培養(yǎng)基培養(yǎng);H2O2組于培養(yǎng)基培養(yǎng)1 h后再加300 μmol/L H2O2100 μL培養(yǎng)24 h;Res+H2O2組給予40 μmol/L Res 100 μL培養(yǎng)1 h,再加300 μmol/L H2O2100 μL培養(yǎng)24 h;Res組給予40 μmol/L Res 100 μL培養(yǎng)24 h。
1.3 THP-1巨噬細(xì)胞衰老情況檢測(cè) 采用衰老相關(guān)β-半乳糖苷酶(SA-β-GA)染色。收集各組細(xì)胞按照1×104/孔接種于六孔板中培養(yǎng)。按SA-β-GA染色試劑盒說明書進(jìn)行操作。各組經(jīng)藥物干預(yù)24 h后PBS漂洗2次,半乳糖染色固定液固定15 min。吸凈固定液,用PBS洗滌細(xì)胞3次,每次3 min。吸凈PBS,每孔加入工作液1 mL,37 ℃、隔絕CO2孵育過夜。顯微鏡下觀察細(xì)胞。鏡下觀察體積增大,胞質(zhì)和胞核出現(xiàn)藍(lán)色沉淀物的細(xì)胞為陽性細(xì)胞,即為衰老細(xì)胞。每次隨機(jī)挑選100個(gè)細(xì)胞,計(jì)算衰老細(xì)胞比例。
1.4 THP-1巨噬細(xì)胞內(nèi)Rb、pRb、LC3、Beclin1、p62蛋白表達(dá)檢測(cè) 采用Western blotting法。收集細(xì)胞孵育24 h后去上清培養(yǎng)液,用冰冷的PBS漂洗3次,加入細(xì)胞裂解液(RIPA∶PMSF∶PI=100∶1∶1),冰上靜置30 min,收集細(xì)胞裂解產(chǎn)物,于4 ℃、12 000 r/min離心15 min,取上清蛋白于-80 ℃保存?zhèn)溆?1個(gè)月內(nèi)檢測(cè))。按試劑盒BCA法測(cè)定蛋白濃度。各取蛋白30 μg,加5×SDS上樣緩沖液,100 ℃變性10 min,凝膠電泳分離,然后電轉(zhuǎn)移至PVDF膜上,5%脫脂奶粉溶液封閉1 h,PVDF膜與一抗(1∶1 000)4 ℃孵育14~16 h,TBST漂洗3×10 min,加羊抗兔IgG二抗(1∶5 000)室溫下孵育1.5 h,TBST漂洗3×10 min,用化學(xué)發(fā)光液顯色。以β-actin為內(nèi)參照,采用Image-ProPlus6.0圖像分析軟件分析蛋白條帶積分吸光度(OD)值,以靶蛋白OD值與β-actin OD值的比值作為靶蛋白相對(duì)表達(dá)量。
1.5 THP-1巨噬細(xì)胞培養(yǎng)液中TNF-α、IL-1β、IL-6水平檢測(cè) 收集各組細(xì)胞,將其接種于96孔板中培養(yǎng)24 h,收集上清液。按ELISA試劑盒說明書進(jìn)行操作,制作標(biāo)準(zhǔn)樣本,標(biāo)準(zhǔn)品孔加標(biāo)準(zhǔn)品50 μL、鏈霉親和素-HRP 50μL;樣品孔加入樣品40 μL、生物素標(biāo)記的抗炎因子抗體10 μL、鏈霉親和素-HRP 50 μL。輕輕振蕩混勻,37 ℃孵育1 h;PBS洗滌3次,每次3 min,洗凈PBS,依次加入顯色劑A、B各50 μL,37 ℃避光孵育5 min,加入終止液50 μL,用酶標(biāo)儀測(cè)波長(zhǎng)450 nm處OD值,根據(jù)標(biāo)準(zhǔn)品濃度及對(duì)應(yīng)OD值計(jì)算各樣品濃度。以上實(shí)驗(yàn)均重復(fù)3~5次,取平均值。
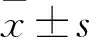
2 結(jié)果
2.1 各組衰老細(xì)胞比例比較 培養(yǎng)24 h,空白對(duì)照組、H2O2組、Res+H2O2組、Res組衰老細(xì)胞比例分別為13%、80%、50%、12%。與空白對(duì)照組比較,H2O2組衰老細(xì)胞比例高(P<0.05);與H2O2組比較,Res+H2O2組衰老細(xì)胞比例低(P<0.05)。
2.2 各組細(xì)胞Rb、p-Rb、LC3B、Beclin-1、p62蛋白表達(dá)比較 與空白對(duì)照組比較,H2O2組Rb蛋白表達(dá)高、p-Rb蛋白表達(dá)低(P均<0.05);與H2O2組比較,H2O2+Res組Rb蛋白表達(dá)低、p-Rb蛋白表達(dá)高(P均<0.05)。與空白對(duì)照組比較,H2O2組,LC3-Ⅱ、Beclin-1蛋白表達(dá)及LC3-Ⅱ/LC3-Ⅰ低,p62蛋白表達(dá)高(P均<0.05);與H2O2組比較,H2O2+Res組LC3-Ⅱ、Beclin-1蛋白表達(dá)及LC3-Ⅱ/LC3-Ⅰ高,p62蛋白表達(dá)低(P均<0.05)。空白對(duì)照組與Res組以上指標(biāo)比較,P均>0.05。見表1。
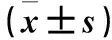
表1 各組細(xì)胞p-Rb、Rb、LC3B、Beclin-1、p62蛋白相對(duì)表達(dá)量及LC3-Ⅱ/LC3-Ⅰ比較
2.3 各組細(xì)胞培養(yǎng)液中TNF-α、IL-1β、IL-6水平比較 與空白對(duì)照組比較,H2O2組TNF-α、IL-1β、IL-6水平高(P均<0.05);與H2O2組比較,H2O2+Res組TNF-α、IL-1β、IL-6水平低(P均<0.05);空白對(duì)照組與Res組TNF-α、IL-1β、IL-6水平比較,P均>0.05。見表2。
3 討論
大量細(xì)胞和分子在衰老過程中發(fā)生了變化,加速了炎癥性疾病的發(fā)生發(fā)展,如AS。AS發(fā)生發(fā)展過程中的核心因素之一是炎癥。炎癥不僅參與AS的形成過程,而且引發(fā)急性血栓、斑塊破裂等并發(fā)癥[6]。單核源性巨噬細(xì)胞作為AS中重要的炎性細(xì)胞在AS的發(fā)生發(fā)展中起重要作用,不僅參與泡沫細(xì)胞的形成,且合成、分泌大量的炎性物質(zhì),如TNF-α、IL-1β、IL-6[5]。前期研究已成功用氧化應(yīng)激誘導(dǎo)心肌細(xì)胞、成纖維細(xì)胞、血管內(nèi)皮細(xì)胞衰老[7,8]。另外相關(guān)研究表明,氧化低密度脂蛋白可促進(jìn)THP-1巨噬細(xì)胞增殖[9]。AS中巨噬細(xì)胞隨著慢性氧化應(yīng)激的累積發(fā)生衰老,最終壞死,導(dǎo)致斑塊破裂誘發(fā)急性心腦血管事件,提示抗氧化劑可延緩衰老。細(xì)胞衰老過程受控于細(xì)胞信號(hào)傳遞系統(tǒng),Rb蛋白調(diào)控著細(xì)胞周期中最關(guān)鍵的調(diào)控點(diǎn)G1/S期,Rb蛋白磷酸化后失活,釋放核轉(zhuǎn)錄因子E2F,游離的E2F啟動(dòng)G1/S期轉(zhuǎn)換相關(guān)基因表達(dá),推動(dòng)細(xì)胞周期的進(jìn)程[10]。本研究發(fā)現(xiàn),與空白對(duì)照組相比,H2O2組Rb蛋白表達(dá)高,p-Rb蛋白表達(dá)低。說明本研究所設(shè)計(jì)的衰老細(xì)胞模型可靠。
表2 各組細(xì)胞培養(yǎng)液中TNF-α、IL-1β、IL-6水平比較
Res具有廣泛的生物活性及藥理作用,能夠抵抗或延緩與衰老以及氧化等相關(guān)的疾病[11]。但目前Res對(duì)巨噬細(xì)胞衰老的影響研究還較少。本研究發(fā)現(xiàn),與H2O2組比較,40 μmol/L的Res能上調(diào)pRb蛋白表達(dá)、下調(diào)Rb蛋白表達(dá),說明Res可減輕H2O2誘導(dǎo)的THP-1巨噬細(xì)胞衰老。
自噬是真核細(xì)胞中普遍存在的一種保護(hù)機(jī)制,通過自噬可以清除細(xì)胞中損傷或衰老的細(xì)胞器及生物大分子,來維持細(xì)胞的穩(wěn)態(tài)[12]。研究表明,抑制細(xì)胞自噬可導(dǎo)致早衰;而衰老的組織和細(xì)胞中自噬減少,例如正常老化的人腦組織中Beclin-1、LC3-Ⅱ等蛋白表達(dá)下調(diào)[13],提示衰老的機(jī)制之一可能是細(xì)胞自噬功能下調(diào)。本研究發(fā)現(xiàn),與空白對(duì)照組比較,H2O2組細(xì)胞Becin-1、LC3-Ⅱ、LC3-Ⅰ蛋白表達(dá)低,p62蛋白表達(dá)高;而給予40 μmol/L的Res干預(yù)后,Beclin-1、LC3-Ⅱ、LC3-Ⅰ蛋白表達(dá)上調(diào),p62蛋白表達(dá)下調(diào),表明Res可能通過調(diào)控細(xì)胞自噬來干預(yù)H2O2誘導(dǎo)的巨噬細(xì)胞衰老,維持細(xì)胞穩(wěn)態(tài)。研究發(fā)現(xiàn),巨噬細(xì)胞通過自噬在AS中起保護(hù)作用[14]。在AS中,炎癥的激活可導(dǎo)致蛋白酶分泌,組織破壞和斑塊破裂[6]。TNF-α、IL-1β、IL-6都是典型的炎性因子,Ida等[15]研究發(fā)現(xiàn),巨噬細(xì)胞通過自噬可明顯抑制炎性因子的分泌。本研究發(fā)現(xiàn),與H2O2組比較,H2O2+Res組TNF-α、IL-1β、IL-6水平低,說明通過自噬可以抑制巨噬細(xì)胞炎性因子的分泌。
綜上所述,本研究Res通過誘導(dǎo)自噬反應(yīng)抑制了H2O2誘導(dǎo)的THP-1巨噬細(xì)胞衰老,至于其相關(guān)信號(hào)通路需進(jìn)一步進(jìn)行探討。這可能是治療AS和其他炎癥疾病的一種新思路。
[1] Shayganni E, Bahmani M, Asgary S, et al. Inflammaging and cardiovascular disease: Management by medicinal plants[J]. Phytomedicine, 2016,23(11):1119-1126.
[2] Franceschi C, Capri M, Monti D, et al. Inflammaging and anti-inflammaging: a systemic perspective on aging and longevity emerged from studies in humans[J]. Mech Ageing Dev, 2007,128(1):92-105.
[3] Baur JA, Sinclair DA. Therapeutic potential of resveratrol: the in vivo evidence[J]. Nat Rev Drug Discov, 2006,5(6):493-506.
[4] Bradamante S, Barenghi L, Villa A. Cardiovascular protective effects of resveratrol[J]. Cardiovasc Drug Rev, 2004,22(3):169-188.
[5] MooreK J, Tabas I. Macrophages in the pathogenesis of atherosclerosis[J]. Cell, 2011,145(3):341-355.
[6] Deguchi J, Aikawa M, Tung C, et al. Inflammation in atherosclerosis: visualizing matrix metalloproteinase action in macrophages in vivo[J]. Circulation, 2006,114(1):55-62.
[7] Ruan YJ, Wu SZ, Zhang L, et al. Retarding the senescent of vascular endothelial cells induced by H2O2: effects of estradial mediated mitochondria protecting[J]. Biogerontology, 2014,15(4):167-375.
[8] Ma Y, Wu SZ, Zhong Y, et al. Polydatin inhibits the oxidative stress-induced proliferation of vascular smooth muscle cells by activating the eNOS/SIRT1 pathway[J]. J Mol Med, 2016,6:1652-1660.
[9] 黃官夢(mèng)茜,蔣米爾,黃英,等.氧化低密度脂蛋白對(duì)巨噬細(xì)胞的作用及機(jī)制[J].中華實(shí)驗(yàn)外科雜志,2015,32(1):63-66.
[10] Sama T, Ethel R, John WL. Role of pRb dephosphorylation in cell cycle regulation[J]. Front Biosci, 2000,1(5):121-137.
[11] 胡凌云,王海鈉,裴俊俊,等.白藜蘆醇防癌抗癌作用及其分子機(jī)制研究進(jìn)展[J].山東醫(yī)藥,2010,50(5):111-112.
[12] Xie Z, Klionsky DJ. Autophagosome formation:core machinery and adaptations[J]. Nat Cell Biol, 2007,9(10):1102-1109.
[13] Rubinsztein DC, Mari OG, Kroemer G. Autophagy and Aging[J]. Cell, 2011,146(5):682-695.
[14] Liao X, Sluimer JC, Wang Y, et al. Macrophage autophagy plays a protective role in advanced atherosclerosis[J]. Cell Metab, 2012,15(4):545-553.
[15] Ida P, Saveria A. ROS-dependent activation of autophagy through the PI3K/Akt/mTOR pathway is induced by hydroxysafflor yellow a-sonodynamic therapy in THP-1 macrophages[J]. Oxid Med Cell Longev, 2017,2017:1-16.
10.3969/j.issn.1002-266X.2017.37.012
R329.2
A
1002-266X(2017)37-0037-03
吳賽珠(E-mail:wusaizhuDr@126.com)
2017-07-03)
