細胞周期調控基因及Bcl-2、c-Jun基因在人肝細胞脂肪變性過程中的表達變化
2017-11-15 07:48:44包成畢玉晶楊瑞馥智發朝
山東醫藥 2017年37期
包成,畢玉晶,楊瑞馥,智發朝
(1南方醫科大學南方醫院·廣東省南方消化病研究所,廣州510515;2北京軍事醫學科學院微生物流行病研究所)
細胞周期調控基因及Bcl-2、c-Jun基因在人肝細胞脂肪變性過程中的表達變化
包成1,2,畢玉晶2,楊瑞馥2,智發朝1
(1南方醫科大學南方醫院·廣東省南方消化病研究所,廣州510515;2北京軍事醫學科學院微生物流行病研究所)
目的觀察細胞周期調控基因及Bcl-2、c-Jun基因在人肝細胞L02脂肪變性過程中的表達變化。方法以油酸和軟脂酸(2∶1)混合液處理L02細胞0、5、10、20 h,建立人肝細胞脂肪變性模型。用油紅O染色檢測L02細胞內脂質含量;用熒光定量PCR技術檢測L02細胞脂肪變性過程中CDK2、CDK4、CyclinD1、Bcl-2、c-Jun mRNA表達。結果隨造模時間延長,L02細胞內橘紅色脂滴聚集越來越多,至造模20 h,肝細胞染成橘紅色。L02細胞內CDK2 mRNA相對表達量逐漸升高,造模5 h達最高(P均<0.05),隨后逐漸下降;CDK4 mRNA相對表達量逐漸升高,造模10 h達最高(P均>0.05),隨后逐漸下降,造模20 h最低(P均>0.05);不同時間點Cyclin D1 mRNA相對表達量比較差異無統計學意義(P均>0.05);Bcl-2 mRNA相對表達量隨造模時間延長而升高,造模10 h達最高(P<0.05),隨后降低;造模5、10、20 h的c-Jun mRNA相對表達量均較造模0 h高(P均<0.05),但無變化規律。結論人肝細胞L02脂肪變性過程中癌癥相關基因CDK2、CDK4、Bcl-2、c-Jun表達先升高后降低,Cyclin D1表達變化不明顯。
肝細胞脂肪變性;細胞周期;細胞周期蛋白;細胞周期蛋白依賴性激酶;細胞周期蛋白依賴性激酶抑制因子;B細胞白血病-淋巴瘤基因2;c-Jun基因
脂肪肝是最常見的與代謝紊亂相關的慢性肝病,在全世界發病率高,約為11%~46%,并有潛在的嚴重后遺癥[1]。脂肪肝疾病譜包括單純性脂肪肝、脂肪性肝炎、肝纖維化、肝硬化和肝細胞癌。細胞周期分為G1、S、G2、M期。細胞周期轉換主要受細胞周期蛋白(Cyclin)、細胞周期蛋白依賴性激酶(CDK)和CDK抑制因子(CKI)調控。細胞周期調控異常與癌癥發生發展有關。細胞周期紊亂可導致癌癥的發生。B細胞白血病-淋巴瘤基因2(Bcl-2)是抗凋亡基因,可通過抑制細胞凋亡從而介導腫瘤的產生[2]。c-Jun家族癌基因在多種惡性腫瘤發展過程中起重要作用[3]。脂肪肝可以逐步發展為肝癌。研究在肝細胞脂肪變性過程中癌癥相關調控基因的表達變化,對探討脂肪肝發展為肝癌的過程具有重要意義。2016年8月~2017年1月,本研究旨在探究癌癥相關基因(CDK2、CDK4、CyclinD1、Bcl-2、c-Jun基因)在人肝細胞脂肪變性的體外模型中不同造模時相的表達變化。
1 材料與方法
1.1 細胞、試劑及儀器來源 人肝細胞株L02由北京軍事醫學科學院贈送;DMEM培養液購自Neuronbc公司,胰蛋白酶購自GE Healthcare Life Sciences公司,軟脂酸、油酸、油紅O試劑、蘇木精試劑購自Sigma公司;倒置顯微鏡購自Zeiss公司,熒光定量PCR儀器購自Roche公司,SYBR premix ExTaq購自Takara公司。
1.2 細胞培養及模型建立 將L02細胞接種于培養瓶,用含10%胎牛血清的DMEM培養液于37 ℃、5% CO2細胞培養箱內培養,2 d換液1次。待細胞生長至80%~90%時用胰蛋白酶消化,按1∶5比例傳代。取對數生長期細胞接種于六孔培養板進行實驗。加入0.5 mmol/L油酸和軟脂酸(2∶1)混合液處理人L02肝細胞株,建立肝細胞脂肪變性模型。
1.3 L02細胞內脂質含量檢測 采用油紅染色法。分別于油酸和軟脂酸混合液處理(造模)0、5、10、20 h收集相應的脂肪變性L02細胞,加4%多聚甲醛孵育1 h,棄甲醛再加60%異丙醇并孵育5 min,棄60%異丙醇,均勻加油紅O工作液,搖勻并室溫避光孵育20 min,棄油紅O工作液,水洗2~5次,加蘇木精孵育1 min。棄蘇木精,水洗2~5次。用蒸餾水覆蓋細胞,在顯微鏡下觀察并對細胞進行拍照。脂質形成的標準為經油紅染色后L02細胞胞質內形成橘紅色脂滴。
1.4 L02細胞內CDK2、CDK4、CyclinD1、Bcl-2、c-Jun mRNA表達檢測 采用熒光定量PCR技術。分別于造模0、5、10、20 h收集L02細胞提取RNA。配消化DNA體系50 μL(10×DNase Ⅰ buffer 5 μL,rDNase Ⅰ 2 μL,RNA 4~7 μg,剩余以水填充)。于PCR儀中消化,37 ℃、30 min;加入DNA酶失活劑10 μL混勻滅活DNase,離心后取上清液用于逆轉錄。變性:配體系(N6隨機引物1 μL,消化離心后的RNA 23 μL),于PCR儀中進行變性,70 ℃ 10 min,變性后立即放置于-20 ℃冰箱8 min防止復性。逆轉錄:配體系(5×First-Strand buffer 8 μL,0.1 mol/L DTT 4 μL,10 mmol/L dNTP 2 μL,RNase抑制劑1 μL),每管加入15 μL,再加入SuperscriptⅢ逆轉錄酶1 μL。逆轉錄程序:25 ℃、10 min,50 ℃、60 min,70 ℃、15 min,4 ℃恒溫。配體系:SYBR premix 10 μL,上下引物各0.6 μL,H2O 6.8 μL,模板2 μL。引物序列:CDK2正向引物:5′-CATGAGGTGGTGACCCTGTGG-3′,反向引物:5′-GTCGTAGTGCAGCATTTGCG-3′;CDK4正向引物:5′- CATTCTGGTGACAAGTGGTGG-3′,反向引物:5′-CGGCTTCAGAGTTTCCACAGA-3′;Cyclin D1正向引物:5′-TTGTTGAAGTTGCAAAGTCCTGG-3′,反向引物:5′- CGGATGGTTTCCACTTCGC-3′;Bcl-2正向引物:5′- CATGTGTGTGGAGAGCGTCAA-3′,反向引物:5′-GCCGGTTCAGGTACTCAGTCA-3′;c-Jun正向引物:5′- GCCAGGTCGGCAGTATAGTC-3′, 反向引物:5′- GGACTCTGCCACTTGTCTCC-3′。采用熒光定量PCR技術進行檢測。用LCS480 1.50.SP4軟件(Roche)計算CDK2、CDK4、CyclinD1、Bcl-2、c-Jun mRNA相對表達量。
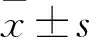
2 結果
油紅O染色結果:隨造模時間延長,L02細胞里的橘紅色脂滴越來越多;造模20 h,L02細胞被染成橘紅色,說明L02細胞發生脂肪變性。隨造模時間延長,CDK2 mRNA相對表達量逐漸升高,造模5 h達最高(P均<0.05),隨后逐漸下降;CDK4 mRNA相對表達量逐漸升高,造模10 h達最高(P均>0.05),隨后逐漸下降,造模20 h最低(P均>0.05);不同時間點Cyclin D1 mRNA相對表達量比較差異無統計學意義(P均>0.05);Bcl-2 mRNA相對表達量隨造模時間的延長而升高,造模10 h達最高(P<0.05),隨后降低;造模5、10、20 h的c-Jun mRNA相對表達量均較造模0 h高(P均<0.05)。見表1。
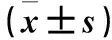
表1 L02細胞不同時間點癌變相關基因表達比較
3 討論
細胞周期轉換主要受Cyclin、CDK和CKI的調控。Cyclins與CDK結合可促進細胞周期的轉換[4]。CDK2的活性對細胞周期的運行至關重要[5]。本研究發現,CDK2、CDK4 mRNA在肝細胞脂肪變性過程中呈一過性表達升高,提示肝脂肪變可能會激活肝細胞CDK2和CDK4的活性,潛在影響細胞周期調控;而CyclinD1 mRNA在肝脂肪變過程中其表達并未出現明顯變化。Rb蛋白磷酸化不足時會出現CyclinD1過表達[6],推測油酸和軟脂酸混合液可能會使Rb蛋白磷酸化。
細胞周期通過正負調節因子作用來維持正常運行,當調節因子失衡時,異常細胞逃離正常細胞周期的監測,從而為細胞癌變的發生提供基礎[7]。癌癥中細胞周期調控基因會發生異常,如CDK2在癌癥組織中可長期呈高表達[8]。因此CDK調節功能和細胞周期檢查位點功能的缺失可與癌癥的分子病理機制有關。研究發現,高脂飲食可促進注射了CT26結腸癌細胞的BALB/c小鼠主動脈微血管生成、實體瘤生長以及腫瘤肺轉移,免疫組化法檢測顯示,小鼠腫瘤組織中cyclin A、Cyclin D1和CDK2的表達明顯增加[9],表明高脂飲食可以破壞正常細胞的周期調控機制,導致細胞進行無限增殖分化。細胞周期調控無論是在肝細胞脂肪變性還是肝細胞癌變的過程中都起到重要作用。本研究表明細胞周期調控對脂肪肝以后發展為肝細胞癌具有重要意義。
Bcl-2是一種抗凋亡基因,在生長因子受體介導的信號傳導途徑中發揮作用,它通過抑制細胞凋亡從而介導腫瘤的發生發展[2]。Bcl-2家族蛋白調節線粒體膜的通透性,使細胞色素C從線粒體泄漏,從而調控細胞凋亡。本研究結果顯示,肝細胞脂肪變性模型可以誘導活化Bcl-2,表明Bcl-2對脂肪肝將來發展為肝細胞癌具有一定的影響。
c-Jun家族癌基因在惡性腫瘤的進展中也起到重要作用。在癌旁異型增生肝組織以及肝癌細胞內出現c-Jun異常表達[3]。JNK信號轉導通路是胰島素抵抗和脂肪酸誘導的肝毒性的關鍵介質。肝細胞在脂毒性的狀態通過上調JNK和Bcl-2相互作用介質而最終發生凋亡。有關腸癌動物模型的研究表明,c-Jun的致瘤功能依賴于JNK對其的磷酸化作用,因此JNK/c-Jun通路有致瘤作用[10,11]。本研究結果顯示,肝細胞脂肪變性可使c-Jun mRNA的表達升高,表明癌癥相關基因c-Jun在脂肪肝時期就有了轉錄水平的升高。
綜上所述,肝細胞脂肪變性模型實驗顯示,細胞周期調控基因、Bcl-2、c-Jun等癌癥相關基因的轉錄水平隨著肝細胞脂肪變性過程的推進,出現了不同程度的改變,而以上調控基因表達異常與癌癥的發生發展密切相關。提示本研究中癌癥相關基因可能參與了脂肪肝發展為肝癌的過程。這為研究脂肪肝的病理生理機制及治療方法提供了新思路。
[1] Berlanga A, Guiu-Jurado E, Porras JA, et al. Molecular pathways in non-alcoholic fatty liver disease[J]. Clin Exp Gastroenterol, 2014,7:221-239.
[2] 苑建磊,蘭濤,周國慶,等.VEGF、Bag-1及Bcl-2在肝癌中的表達及意義[J].臨床和實驗醫學雜志,2014,13(4):268-271.
[3] 張穎超,韓喜春,賈明庫,等.癌基因c-fOS和c-jun在肝細胞型肝癌中的表達及臨床意義[J].吉林大學學報,2002,28(1):46-48.
[4] Gopinathan L, Tan SL, Padmakumar VC, et al. Loss of Cdk2 and cyclin A2 impairs cell proliferation and tumorigenesis[J]. Cancer Res, 2014,74(14):3870-3879.
[5] Mueller S, Huard J, Waldow K, et al. T160-phosphorylated CDK2 defines threshold for HGF-dependent proliferation in primary hepatocytes[J]. Mol Syst Biol, 2015,11(3):795.
[6] Rao SS, Chu C, Khotz DS. Ectopic expression of cyclin D1 prevents activation of gene transcription by myogenic basic-helix-loop-helix regulators[J]. Mol Cell Biol, 1994,14(8):5259-5267.
[7] 盧斌,戴益民.肝癌發生中細胞周期調控的異常[J].世界華人消化雜志,2001,9(2):205-208.
[8] Kawana H, Tamaru J, Tanaka T, et al. Role of p27Kip1 and cyclin-dependent kinase 2 in the proliferation of non-small cell lung cancer[J]. Am J Pathol, 1998,153(2):505-513.
[9] Park H, Kim M, Kwon G T, et al. A high-fat diet increases angiogenesis, solid tumor growth, and lung metastasis of CT26 colon cancer cells in obesity-resistant BALB/c mice[J]. Mol Carcinog, 2012,51(11):869-880.
[10] 包翠芳,魏嘉,梁佳,等.前列地爾對局灶性腦缺血大鼠p-JNK及p-c-Jun表達的影響[J].山東醫藥,2011,51(25):15-16.
[11] 倪桂蓮,崔融,陳干明,等.加巴噴丁對糖尿病神經痛大鼠痛敏和脊髓Glu及下丘腦JNK的影響[J].中華全科醫學,2015,13(11):1757-1759,1907.
Expressionchangesofcellcycleregulatorygenes,Bcl-2,andc-Junintheprocessofhumanhepaticsteatosis
BAOCheng1,BIYujing,YANGRuifu,ZHIFachao
(1NanfangHospital,SouthernMedicalUniversity,Guangzhou510515,China)
ObjectiveTo investigate the expression changes of cell cycle regulatory genes [Cyclin-dependent kinase2 (CDK2), Cyclin-dependent kinase4 (CDK4), and CyclinD1], B-cell lymphoma/leukemia-2 (Bcl-2), and c-Jun in the process of human hepatic steatosis.MethodsL02 liver cell line was incubated with mixture of oleic acid and palmitic acid by the ratio of 2:1 (OP21) for 0, 5, 10, and 20 h to build the models of human hepatocyte steatosis. Oil Red O staining was performed to evaluate the fat content of the L02 liver cells. The mRNA expression levels of CDK2, CDK4, CyclinD1, Bcl-2, and c-Jun in the L02 cells at different time points were detected by real-time PCR.ResultsMore and more orange droplets were accumulated in the L02 cells with the time goes on, and the liver cells were dyed orange at 20 h. The mRNA expression level of CDK2 increased gradually, reached the peak at 5 h (P<0.05), and then decreased gradually; the mRNA expression level of CDK4 increased gradually, reached the peak at 10 h (P>0.05), then decreased gradually, and reached the lowest point at 20 h (P>0.05); no statistically significant difference was found in the expression levels of Cyclin D1 at different time points during the process (allP>0.05); the mRNA expression level of Bcl-2 increased over time, reached the peak at 10 h (P<0.05), and then decreased; the mRNA expression level of c-Jun was higher at 5, 10, and 20 h than that at 0 h (allP<0.05), but there were no regular changes.ConclusionDuring the process of hepatic steatosis, the relative mRNA levels of the cancer-related genes (CDK2, CDK4, Bcl-2, and c-Jun) first increase and then decrease, while the expression of Cyclin D1 does not change significantly.
hepatic steatosis; cell cycle; Cyclin; Cyclin-dependent kinase; Cyclin-dependent kinase inhibitor; B-cell lymphoma/leukemia-2; c-Jun gene
10.3969/j.issn.1002-266X.2017.37.004
R575
A
1002-266X(2017)37-0012-03
國家高技術研究發展計劃(863計劃)(2015AA020702)。
包成(1990-),男,在讀碩士,主要研究方向為脂肪肝。E-mail:908729477@qq.com
智發朝(1962-),男,博士,主任醫師,主要研究方向為膽胰疾病和小腸疾病的微創治療。E-mail:zhifc41532@163.com
2017-06-13)
