植物乳桿菌P158產細菌素培養基及培養條件的優化
2017-11-13 01:36:04徐瓏倩胡凱弟張艾青劉書亮
食品科學 2017年22期
徐瓏倩,胡凱弟,張艾青,周 康,2,劉書亮,2,*
(1.四川農業大學食品學院,四川 雅安 625014;2.四川農業大學食品加工與安全研究所,四川 雅安 625014)
植物乳桿菌P158產細菌素培養基及培養條件的優化
徐瓏倩1,胡凱弟1,張艾青1,周 康1,2,劉書亮1,2,*
(1.四川農業大學食品學院,四川 雅安 625014;2.四川農業大學食品加工與安全研究所,四川 雅安 625014)
為提高乳酸菌細菌素產量,以藤黃微球菌、銅綠假單胞桿菌為指示菌,通過單因素和正交試驗優化植物乳桿菌P158產細菌素的培養基和培養條件。結果表明,5 種乳酸菌培養基中MRS培養基為該菌株產細菌素的適宜培養基;最佳培養條件為種子液接種量3%(V/V)、培養基初始pH 6.0、34℃靜置培養42 h;最佳培養基配方為葡萄糖添加量2 g/100 m L、酵母浸膏添加量2 g/100 mL、大豆蛋白胨添加量1.5 g/100 m L、MgSO4添加量0.058 g/100 mL、MnSO4添加量0.025 g/100 m L、FeSO4添加量0.02 g/100 m L、Tween 80添加量0.08 g/100 m L、乙酸鈉添加量0.5 g/100 m L、K2HPO4添加量0.2 g/100 m L。在此條件下,細菌素效價為1 145 IU/m L,較優化前(362 IU/m L)提高了216%。
植物乳桿菌;細菌素;培養基;培養條件
乳酸菌是一類利用可發酵性糖產生大量乳酸的細菌[1-2],在食品工業中有廣泛應用[3-4],而乳酸菌細菌素(乳酸菌素)是菌體在代謝過程中由核糖體產生的具有抑菌活性的多肽、蛋白質或蛋白質復合物[5-6]。該類物質可作為天然防腐劑(如乳酸鏈球菌素)應用于食品中[7],由于不產生抗藥性,對人體無害,其應用前景廣闊[8]。乳酸菌素產生于菌體對數生長中期,在對數后期或穩定前期達到最大值,其產量與細菌生長相關[9-12]。培養溫度、接種量、初始pH值等培養條件[13-14],以及培養基種類、碳源、氮源等營養條件[15-16]都對乳酸菌素產量有直接或間接影響。植物乳桿菌屬于我國衛生部關于“可用于食品的菌種名單”中公布的益生菌之一,能夠通過代謝產生的過氧化氫、有機酸、細菌素以及其他競爭營養物質達到抑菌作用[15,17],延長食品保質期。植物乳桿菌(Lactobacillu splantarum)P158分離于四川傳統發酵食品,可產生廣譜植物乳桿菌素,對多數革蘭氏陰性菌和革蘭氏陽性菌及少數真菌有較強抑制活性,有望作為食品生物防腐劑[18-19];但較低的產量限制了其應用。因此,研究菌株P158產細菌素的影響因素,優化發酵條件,從而提高細菌素產量,為其應用提供參考依據。
1 材料與方法
1.1 材料與試劑
1.1.1 菌種
植物乳桿菌(L. plantarum)P158[18]、藤黃微球菌(M icrococcus luteus)CICC10209、銅綠假單胞桿菌(Pseudomonas aeruginosa)ATCC27853作指示菌,由四川農業大學食品微生物實驗室提供。
1.1.2 試劑
乳酸鏈球菌素(效價106IU/g) 浙江銀象生物工程有限公司;植物蛋白胨、聚蛋白胨、胰蛋白胨、大豆蛋白胨均為生物試劑 北京奧博星生物技術有限責任公司;β-甘油磷酸二鈉、抗壞血酸、蔗糖、乳糖、半乳糖、麥芽糖、木糖、纖維二糖、山梨醇、可溶性淀粉等均為分析純試劑 成都科龍試劑廠。
1.1.3 培養基
營養肉湯培養基配方:牛肉膏3 g、蛋白胨10 g、NaCl 5 g、水1 000 m L,調至pH 7.0~7.2。
CM培養基配方:蔗糖10 g、蛋白胨10 g、酵母浸膏10 g、K2HPO410 g、NaCl 2 g、MgSO4·7H2O 0.2 g、水1 000 m L,調至pH 6.2。
MRS培養基配方:蛋白胨10 g、牛肉膏10 g、酵母浸膏5 g、葡萄糖20 g、K2HPO42 g、檸檬酸二銨2 g、乙酸鈉5 g、M gSO40.58 g、M nSO40.25 g、Tween 80 1 m L、水1 000 m L,調至pH 6.5。
M 17培養基配方:植物蛋白胨5 g、聚蛋白胨5 g、牛肉膏2.5 g、酵母浸膏5 g、β-甘油磷酸二鈉19 g、抗壞血酸0.5 g、MgSO4·7H2O 0.25 g、水1 000 m L,調至pH 7.1。
BCP培養基配方:酵母浸膏2.5 g、蛋白胨5 g、葡萄糖5 g、溴甲酚紫0.04 g、水1 000 m L,調至pH 6.8~7.0。
APT培養基配方:胰蛋白胨10 g、酵母浸膏5 g、檸檬酸鈉5 g、葡萄糖10 g、K2HPO45 g、Tween 80 1 m L、NaCl 5 g、MgSO4·7H2O 0.8 g、MnCl2·4H2O 0.14 g、FeSO4·7H2O 0.04 g、水1 000 m L,調至pH 6.7~7.0。
以上培養基添加20 g/L瓊脂粉為相應固體培養基,均于121 ℃滅菌15 min。
1.2 儀器與設備
PHS-4C+酸度計 成都世紀方舟科技有限公司;Sorvall ST 16R冷凍離心機、1300 Series A2生物安全柜美國Thermo Fisher Scientific公司;DHP-9162電熱恒溫培養箱 上海一恒科學儀器有限公司。
1.3 方法
1.3.1 指示菌活化及其菌懸液制備
分別挑取藤黃微球菌CICC10209和銅綠假單胞桿菌ATCC27853劃線于營養瓊脂斜面,37 ℃培養24 h進行活化;用5 m L無菌生理鹽水洗下菌苔,調整細胞濃度為107CFU/m L。
1.3.2 植物乳桿菌P158活化及其種子液制備
將菌株P158劃線于MRS斜面,37 ℃培養24 h進行活化;用5 m L無菌生理鹽水洗下菌苔,調整細胞濃度為108CFU/m L作為種子液。
1.3.3 發酵上清液的制備
將菌株P158種子液按體積分數1%接種至10 m L發酵培養基中,30 ℃靜置培養48 h后離心(10 000 r/m in,10 m in)得到發酵上清液。
1.3.4 抑菌實驗
采用牛津杯雙層瓊脂擴散法,并稍作改進[20]。在鋪有10 m L水瓊脂平板上放置牛津杯,吸取200 μL指示菌懸液至20 m L 50 ℃左右的無菌營養瓊脂培養基,混勻后傾注平板,放置15 min。向牛津杯內加入100 μL菌株P158發酵上清液,37 ℃正置培養16~18 h,分別測量發酵上清液對指示菌CICC10209和ATCC27853抑菌圈直徑,設置添加等量無菌MRS液體培養基為對照;同時測定發酵上清液pH值。
1.3.5 細菌素效價測定
將發酵上清液用質量分數70% (NH4)2SO4溶液沉淀,4 ℃鹽析過夜后離心(10 000 r/m in,20 m in),棄去上清液;利用20 mmol/L pH 4.5的檸檬酸緩沖液復溶后用于效價測定。效價測定方法:以藤黃微球菌10209為指示菌,選擇100、500、1 000、1 500、2 000 IU/m L 5 個濃度,采用牛津杯雙層瓊脂擴散法做抑菌實驗,以效價對數值為橫坐標,抑菌圈直徑為縱坐標,繪制效價標準曲線,然后對應標準曲線計算植物乳桿菌P158產細菌素的效價。
1.3.6 實驗設計
1.3.6.1 確定植物乳桿菌P158產細菌素適宜培養條件的單因素試驗
將菌株P158種子液按體積分數1%分別接種于CM、MRS、M 17、BCP和APT 5 種培養基中,30 ℃靜置培養48 h,測定其發酵上清液抑菌圈直徑和pH值,以添加各自使用的無菌培養基為對照,確定最適宜的培養基種類。
將菌株P158種子液按體積分數1%接入MRS培養基(pH 6.5),分別于26、30、34、38、42 ℃培養48 h,測其發酵上清液抑菌圈直徑和pH值,確定適宜的培養溫度;將菌株P158種子液按體積分數1%接入MRS培養基(pH 6.5),34 ℃靜置培養,培養12、18、24、30、36、42、48、54、60、66、72 h,測定其發酵上清液抑菌圈直徑和pH值,確定適宜的培養時間。
將菌株P158種子液分別按體積分數1%、2%、3%、4%接入MRS培養基(pH 6.5),34 ℃培養42 h后,測定其發酵上清液抑菌圈直徑和pH值,確定適宜的接種量。
將菌株P158種子液按體積分數3%接入MRS培養基,培養基初始pH值分別調至4.5、5.0、5.5、6.0、6.5、7.0、7.5、8.0,34 ℃靜置培養42 h,測定其發酵上清液抑菌圈直徑和pH值,確定適宜的初始pH值。
1.3.6.2 確定植物乳桿菌P158產細菌素適宜碳/氮源的單因素和正交試驗
在1.3.6.1節獲得的培養菌株P158培養基種類及最佳條件基礎上,以不同碳源、氮源、碳/氮源適宜比例為因素,進行單因素試驗,具體如下:以MRS培養基去除碳源為基礎,分別添加2 g/100 m L的葡萄糖、麥芽糖、半乳糖、纖維二糖、蔗糖、山梨醇、乳糖、木糖和可溶性淀粉,測定菌株P158發酵液抑菌圈直徑和pH值;以MRS培養基去除氮源為基礎,分別添加2 g/100 m L的酵母浸膏、大豆蛋白胨、胰蛋白胨、牛肉膏、蛋白胨、氨水、硝酸鈉和檸檬酸氫二銨,測定菌株P158發酵上清液抑菌圈直徑和pH值。
根據上述實驗結果,進行培養基碳/氮源適宜比例實驗,將葡萄糖作為最適碳源,按不同質量濃度0、1、2、3、4、5、7.5、10 g/100 m L添加于MRS培養基,測菌株P158發酵上清液抑菌圈直徑和pH值;將酵母浸膏按不同質量濃度0、0.5、1、1.5 g/100 m L添加于MRS培養基,測菌株P158發酵上清液抑菌圈直徑及其pH值;除酵母浸膏外,將大豆蛋白胨按不同質量濃度0、0.5、1、1.5、2、2.5、3 g/100 m L添加于MRS培養基中,測菌株P158發酵上清液抑菌圈直徑和pH值。
根據單因素試驗結果,設計L9(34)正交試驗,進一步優化培養基碳/氮源組合。
1.3.6.3 確定植物乳桿菌P158產細菌素適宜金屬離子的單因素和正交試驗
在1.3.6.2節基礎上,分別考察Mg2+、Mn2+和Fe2+對菌株P158產細菌素的影響。將MgSO4按不同質量濃度0、0.029、0.058、0.087、0.116 g/100 m L分別添加于培養基中,在最佳培養條件下培養菌株P158,測定其發酵液上清液抑菌圈直徑和pH值;將M nSO4按不同質量濃度0、0.012 5、0.025、0.037 5、0.05 g/100 m L分別添加于培養基中,在最佳培養條件下培養菌株P158,測定其發酵上清液抑菌圈直徑及其pH值;將FeSO4按不同質量濃度0、0.02、0.04、0.06、0.08 g/100m L分別添加于培養基中,在最佳培養條件下培養菌株P158,測定其發酵上清液抑菌圈直徑及其pH值。
根據單因素試驗結果,設計L9(34)正交試驗,進一步優化金屬離子添加量。
1.3.6.4 確定植物乳桿菌P158產細菌素適宜Tween 80質量濃度
在上述試驗結果基礎上,分別添加不同質量濃度0、0.01、0.02、0.03、0.04、0.06、0.08、0.1 g/100 m L Tween 80,在最佳培養條件下培養菌株P158并進行抑菌實驗,測定其發酵上清液抑菌圈直徑及其pH值。
1.4 數據處理
實驗重復3 次,取其平均值,數據采用SPSS V 22進行統計分析。
2 結果與分析
2.1 植物乳桿菌P158產細菌素適宜培養條件的確定
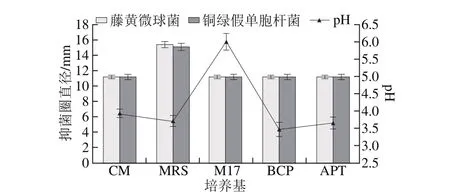
圖1 培養基對菌株P158產細菌素的影響Fig. 1 Effect of different media on production of bacteriocin by strain P158
2.1.1 乳酸菌培養基種類對植物乳桿菌P158產細菌素的影響由圖1可知,5 種培養基對菌株P158產細菌素的效果依次為MRS>APT>CM>BCP>M 17。雖然使用MRS培養基時其發酵液pH值較APT和BCP略高,但是抑菌圈直徑最大(P<0.05),故選擇MRS培養基做后續實驗。
2.1.2 培養溫度對植物乳桿菌P158產細菌素的影響
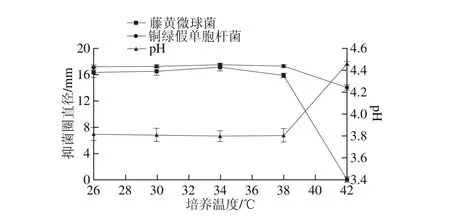
圖2 培養溫度對菌株P158產細菌素的影響Fig. 2 Effect of culture temperature on production of bacteriocin by strain P158
由圖2可知,抑菌圈直徑隨著培養溫度的升高而增大,并在34 ℃時達到最大值(P<0.05);當溫度繼續上升,抑菌圈直徑反而減小甚至為0,發酵液pH值相對較高,可見P158生長已受到影響。
2.1.3 培養時間對植物乳桿菌P158產細菌素的影響
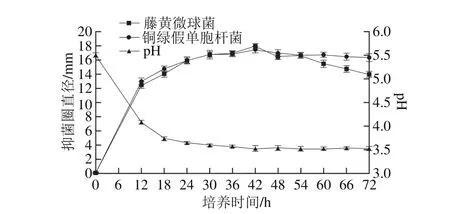
圖3 培養時間對菌株P158產細菌素的影響Fig. 3 Effect of incubation time on production of bacteriocin by strain P158
由圖3可知,隨著培養時間的延長,發酵液pH值持續下降并最終穩定在3.5左右,對2 種指示菌的抑制活性均顯著增強(P<0.05),42 h時最強;當培養超過60 h,抑菌圈直徑均開始減小;故選擇42 h作為最佳培養時間。
2.1.4 接種量對植物乳桿菌P158產細菌素的影響
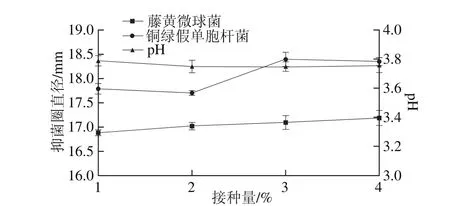
圖4 接種量對菌株P158產細菌素的影響Fig. 4 Effect of inoculum amount on production of bacteriocin by strain P158
由圖4可知,當種子液接種量為3%時,抑菌圈直徑較大。
2.1.5 初始pH值對植物乳桿菌P158產細菌素的影響
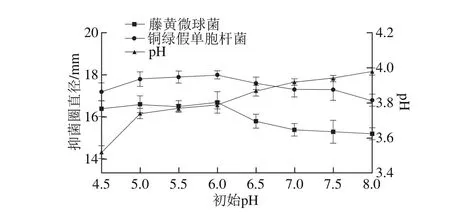
圖5 初始pH值對菌株P158產細菌素的影響Fig. 5 Effect of initial pH on p roduction of bacteriocin by strain P158
由圖5可知,隨著初始pH值的增大,藤黃微球菌和銅綠假單胞桿菌的抑菌圈直徑均呈現先增大后緩慢降低的趨勢;中性和偏堿性環境不利于乳酸菌生長故抑菌活性偏低[21](P<0.05);初始pH值為6.0時抑菌效果最好。
2.2 植物乳桿菌P158產細菌素適宜培養基成分的確定
2.2.1 碳源對植物乳桿菌P158產細菌素的影響
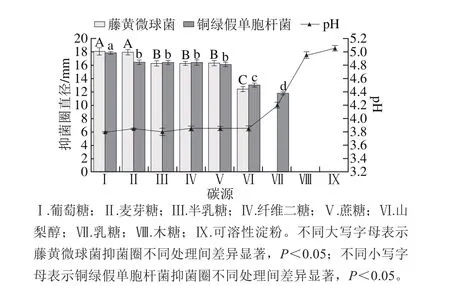
圖6 碳源對菌株P158產細菌素的影響Fig. 6 Effect of carbon sources on p roduction of bacteriocin by strain P158
由圖6可知,以葡萄糖為碳源時,菌株發酵上清液對藤黃微球菌抑制活性與添加麥芽糖相比無明顯差異(P>0.05),但顯著優于半乳糖等其他處理組(P<0.05);并且銅綠假單胞桿菌抑菌圈最大(P<0.05),pH值最低;當以乳糖為碳源時,P158對藤黃微球菌沒有活性;而添加木糖和可溶性淀粉時,藤黃微球菌和銅綠假單胞桿菌均未出現抑菌圈,此現象與Turgis等[22]研究結果相似。因此選擇葡萄糖作為最佳碳源。
2.2.2 氮源對植物乳桿菌P158產細菌素的影響
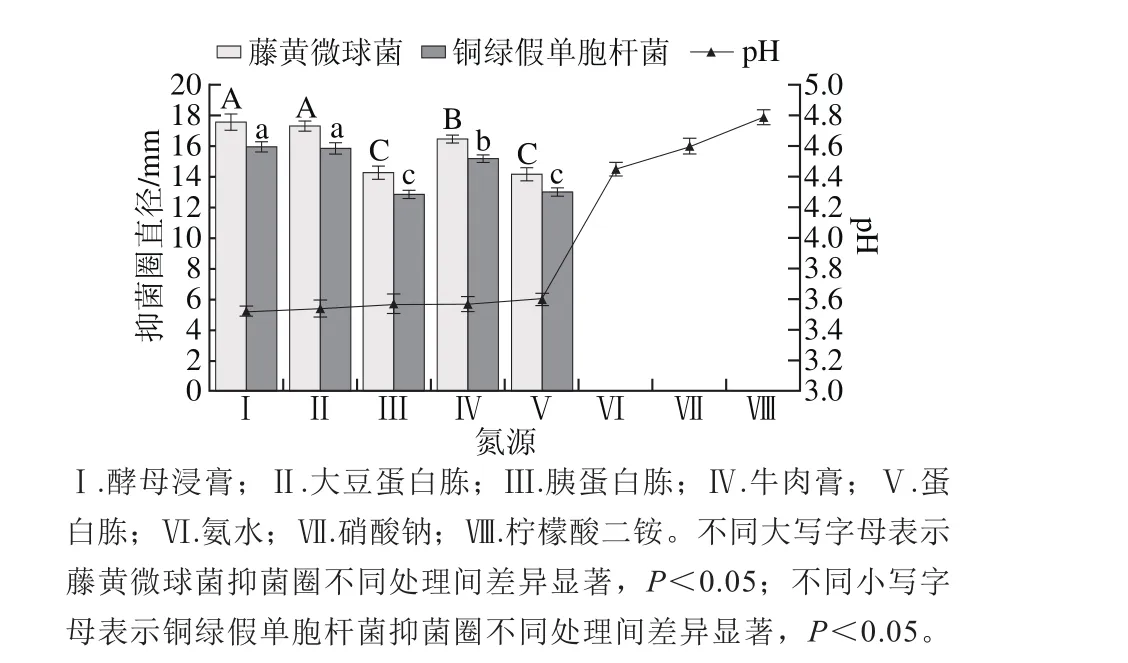
圖7 氮源對菌株P158產細菌素的影響Fig. 7 Effect of nitrogen sources on production of bacteriocin by strain P158
由圖7可知,相比于無機氮源,有機氮源更有利于菌株產細菌素,且不同氮源之間差異較大。酵母浸膏和大豆蛋白胨為氮源時,菌株P158產細菌素能力遠強于其他氮源(P<0.05),但二者之間并無顯著性差異(P>0.05);二者發酵液pH值分別為3.54與3.52,幾乎無差異。
2.2.3 培養基碳/氮源適宜比例
2.2.3.1 葡萄糖質量濃度對植物乳桿菌P158產細菌素的影響
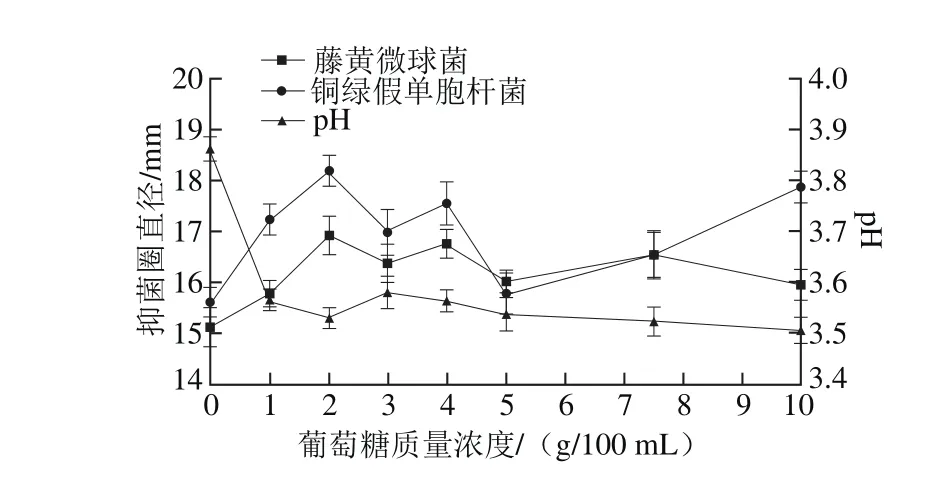
圖8 葡萄糖質量濃度對菌株P158產細菌素的影響Fig. 8 Effect of glucose concentration on production of bacteriocin by strain P158
由圖8可知,當葡萄糖質量濃度維持在1~3 g/100 m L時,2 種抑菌圈直徑均較大,pH值變化較小(維持pH 3.5左右);2 g/100 m L時效果顯著(P<0.05)。
2.2.3.2 酵母浸膏質量濃度對植物乳桿菌P158產細菌素的影響
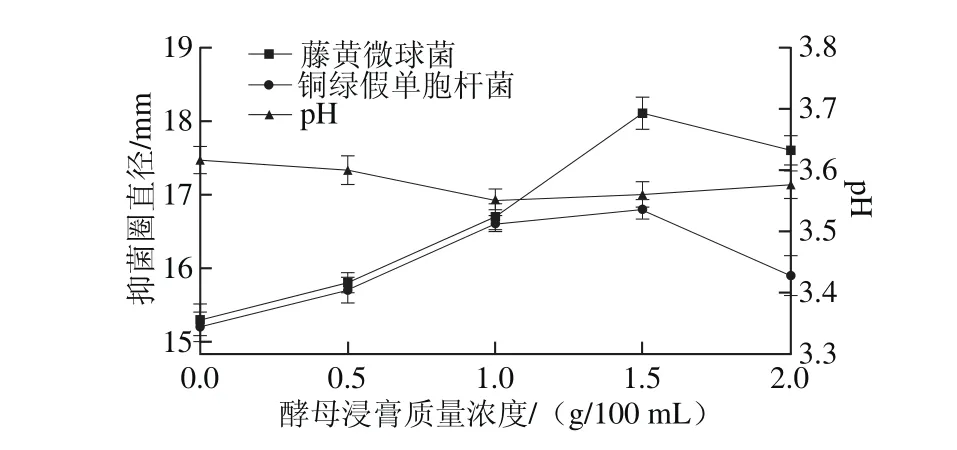
圖9 酵母浸膏質量濃度對菌株P158產細菌素的影響Fig. 9 Effect of yeast extract concentration on p roduction of bacteriocinby strain P158
由圖9可知,質量濃度的酵母浸膏對植物乳桿菌P158產細菌素影響較大;當添加1~2 g/100 m L時,抑菌圈直徑較其他實驗組大(P<0.05)。
2.2.3.3 大豆蛋白胨質量濃度對植物乳桿菌P158產細菌素的影響
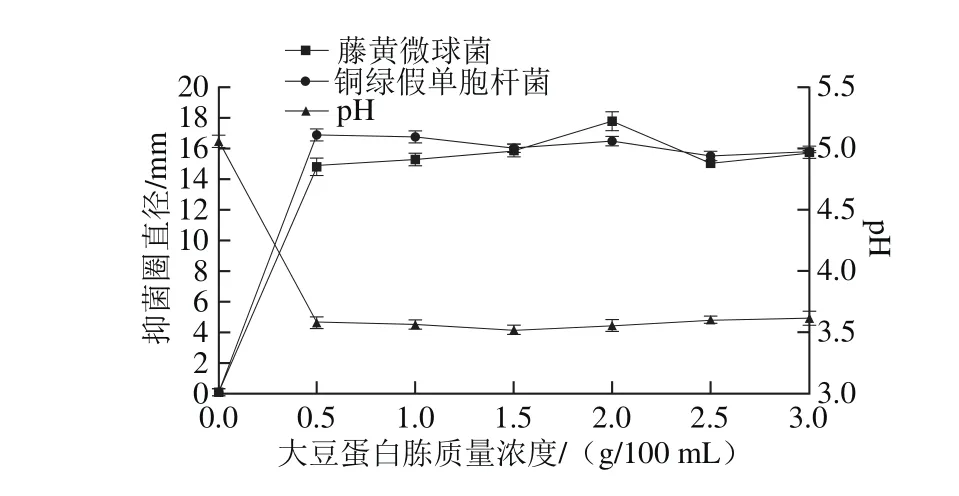
圖10 大豆蛋白胨質量濃度對菌株P158產細菌素的影響Fig. 10 Effects of soy peptone concentration on production of bacteriocin by strain P158
如圖10所示,當大豆蛋白胨添加量在1~2 g/100 m L時,P158發酵液對2 種指示菌的抑制活性較高,且pH值保持平穩,維持pH 3.5左右。
2.2.3.4 培養基碳/氮源組分正交試驗結果
根據單因素試驗結果,設計L9(34)正交試驗,進一步優化培養基碳/氮源組合,試驗設計與結果見表1,直觀分析見表2。由表2可知,影響菌株P158對2 種指示菌抑制活性的主次因素為葡萄糖>酵母浸膏>大豆蛋白胨。根據正交試驗結果,試驗號5和9對藤黃微球菌抑菌圈直徑顯著大于其他試驗組號(P<0.05);而銅綠假單胞桿菌則是試驗號4~9組優于試驗號1~3組,且第5和第9試驗號之間無顯著差異(P>0.05)。考慮到成本問題,故選擇第5組作為碳/氮源最佳配比。
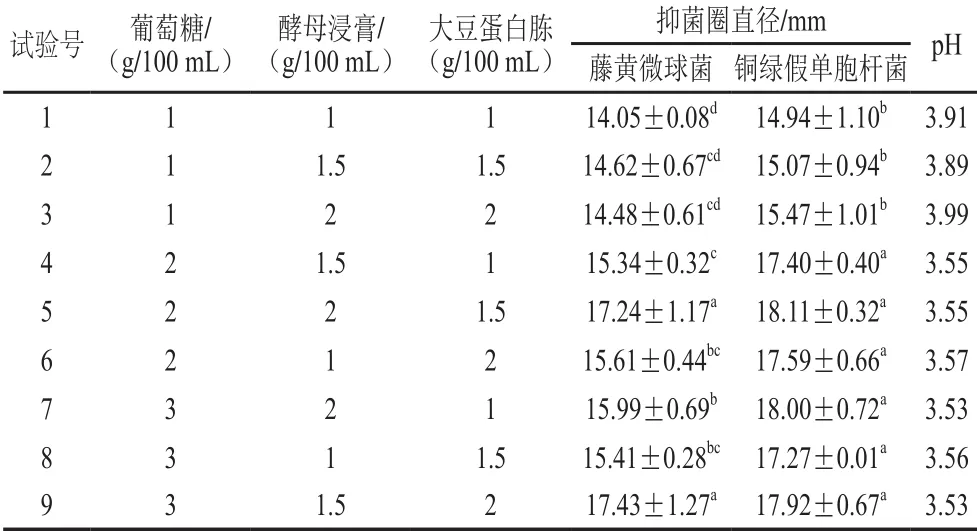
表1 培養基碳/氮源組分正交試驗優化及結果Table 1 O rthogonal array design w ith response values for the op tim ization of glucose, yeast extract and soy pep tone concentration in the medium
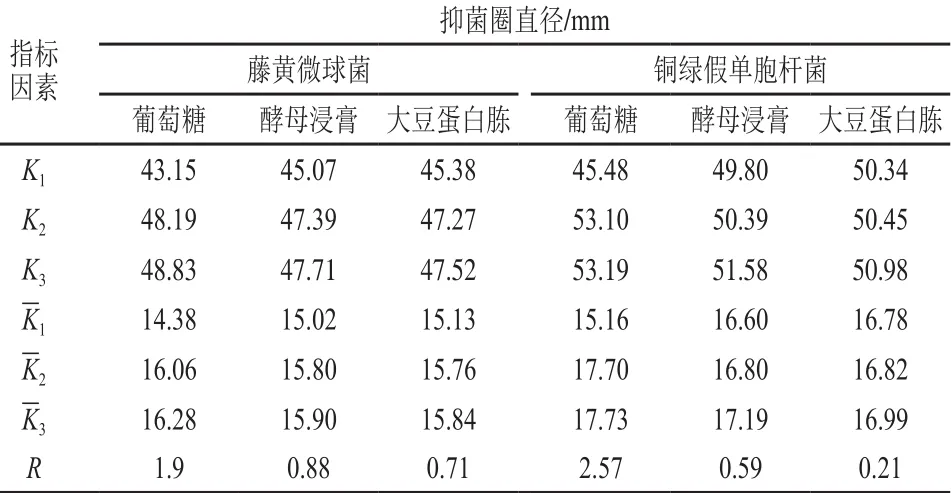
表2 培養基碳/氮源組分正交試驗直觀分析Table 2 Range analysis of bacteriostatic activity
2.2.4 金屬離子成分
2.2.4.1 單因素試驗結果
由圖11可知,隨著M g2+質量濃度逐漸增大,抑菌圈直徑表現為先增大后減小,其中銅綠假單胞桿菌抑菌圈在M g2+質量濃度為0.058 g/100 m L時達到最大(P<0.05)。當添加量為0.058~0.116 g/100 m L時,抑菌活性較強,故選擇此范圍做后續實驗。
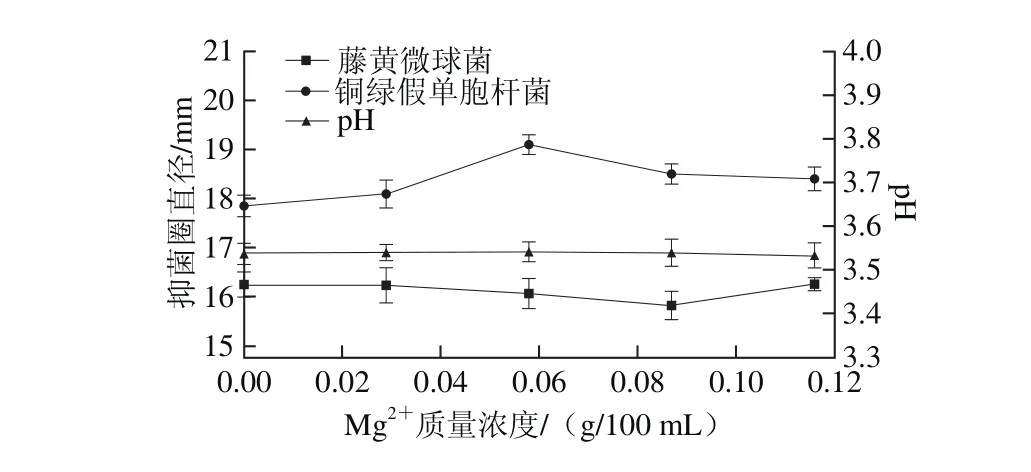
圖11 M g2質量濃度對菌株P158產細菌素的影響Fig. 11 Effect of M g2+ concentration on production of bacteriocin by strain P158
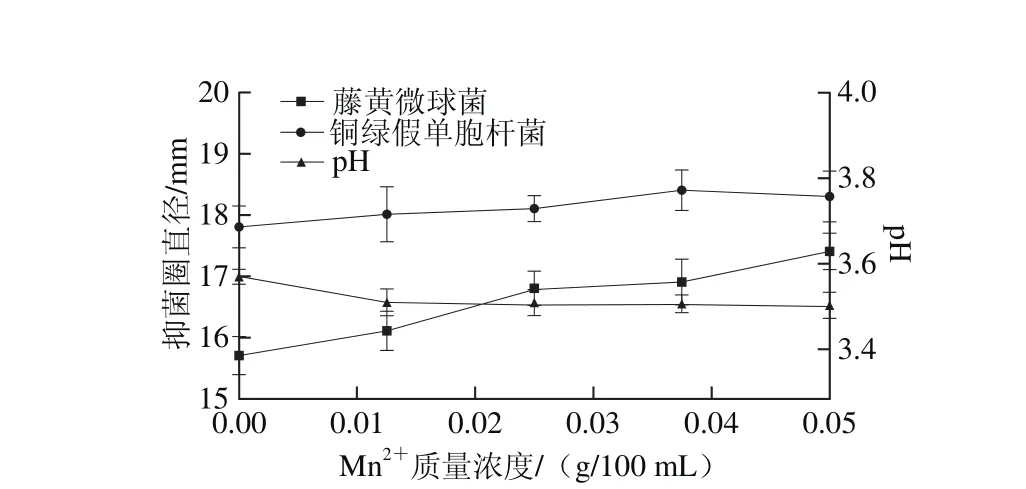
圖12 M n2質量濃度對菌株P158產細菌素的影響Fig. 12 Effect of M n2+ concentration on production of bacteriocin by strain P158
由圖12可知,Mn2+質量濃度在0.025~0.05 g/100 m L之間,菌株P158發酵上清液對2 種指示菌有較大抑制活性(P<0.05)。
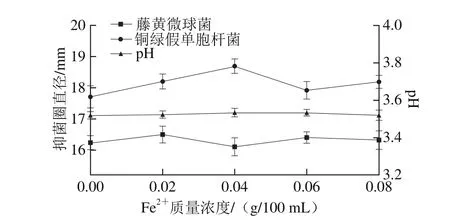
圖13 Fe2質量濃度對菌株P158產細菌素的影響Fig. 13 Effect of Fe2+ concentration on production of bacteriocin by strain P158
由圖13可知,菌株P158細菌素產量在Fe2+添加量為0.02~0.06 g/100 m L時較高。
2.2.4.2 培養基金屬離子正交試驗結果
根據單因素試驗結果,設計L9(33)正交試驗,進一步考察金屬離子比例對菌株P158產細菌的影響。試驗設計及結果見表3,直觀分析見表4。P158對藤黃微球菌抑制效果影響主次因素為Mg2+>Mn2+>Fe2+,最優組合為試驗2;而對銅綠假單胞桿菌則是M n2+>Fe2+>M g2+,試驗1效果最好;顯著性分析表明,二者差異不顯著。從成本角度考慮,選擇試驗1中各項水平作為最優組合添加于培養基中。
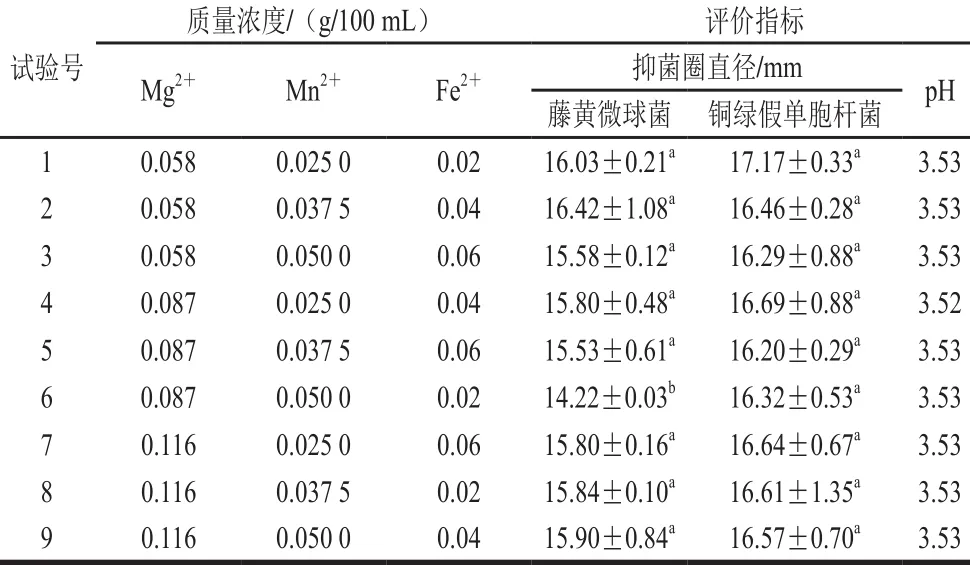
表3 培養基金屬離子正交試驗設計及結果Table 3 Orthogonal array design w ith response values for the optim ization of m etal ion concentrations in the medium
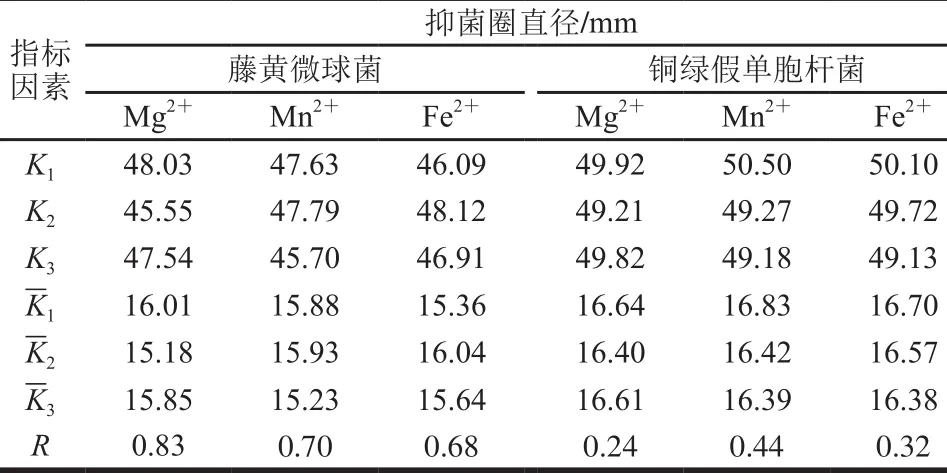
表4 培養基金屬離子正交試驗直觀分析Table 4 Range analysis of bacteriostatic activity
2.2.5 Tween 80質量濃度的影響
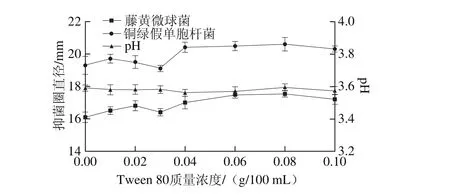
圖14 Tween 80質量濃度對菌株P158產細菌素的影響Fig. 14 Effect of Tween 80 concentration on production of bacteriocin by strain P158
由圖14可知,Tween 80對菌株P158產細菌素有一定刺激作用;其質量濃度為0.08 g/100 m L時,發酵上清液抑菌活性最強(P<0.05)。
2.3 植物乳桿菌素P158的效價結果
根據實驗結果,以效價對數值為自變量,抑菌圈直徑為因變量進行線性擬合,得到效價線性回歸方程:y=4.432 6x+3.980 7(R2=0.997 8)。分別以優化前后條件培養菌株P158,發酵液經粗提后得到細菌素粗提物,按1.3.4節分別測定其抑菌圈直徑和pH值。結果表明,優化前的抑菌圈為15.32mm,優化后為17.54mm;分別代入回歸方程計算得到效價依次為362 IU/m L和1 145 IU/m L,提高了216%;優化前后的pH值一致。該結果優于陳蕓蕓等[23]和Pal等[24]的研究。
乳酸菌素作為生物防腐劑,可提高食品安全性與品質[18]。其產量與菌體生長有關,但最佳發酵條件往往與菌體最適生長條件不一致[10-11],溫度、pH值、氮源、碳源等諸多因素對乳酸菌素產量有不同程度的影響,盡管原因尚未系統闡明[25],但優化乳酸菌素產量并提高其活性顯得尤為重要[26]。Tabbene等[14]從培養基成分入手,對枯草芽孢桿菌(Bacillus subtilis)B38產細菌素條件進行研究,結果表明乳糖、(NH4)2SO4和Mn2+對其產量影響顯著;Anthony等[27]則發現高濃度酵母浸膏和NaCl對細菌素產量有正效應,堿性環境和較高的培養溫度亦是如此;而Raza等[28]考察了不同金屬離子對Paenibacillus polymyxa SQR-21產細菌素的影響,結果表明Zn2+對產量有抑制作用,而M g2+相反,Fe2+雖然影響不顯著,但與Mg2+有協同作用。縱觀優化乳酸菌素產量的文獻,大多局限于某一方面因素;與之不同的是,本實驗不僅從培養條件,也從培養基成分方面,對植物乳桿菌P158產乳酸菌素發酵條件進行了研究,系統考察了不同因素的影響效果并作優化,為菌株的應用提供了更詳細的數據參考。此外,實驗結果表明Tween 80對植物乳桿菌P158產細菌素有一定促進作用,這與Collado等[29]報道相一致,可能是Tween 80作為一種乳化劑,改變了菌體細胞膜通透性,從而有利于營養物質和代謝產物的運輸[30]。
3 結 論
植物乳桿菌P158產細菌素的適宜培養基為MRS,其最佳培養條件為種子液接種量3%(V/V)、培養基初始pH 6、34℃靜置培養42 h。最佳培養基配方為葡萄糖質量濃度2 g/100 m L、酵母浸膏質量濃度2 g/100 m L、大豆蛋白胨質量濃度1.5 g/100 m L、M gSO4質量濃度0.058 g/100 m L、M nSO4質量濃度0.025 g/100 m L、FeSO4質量濃度0.02 g/100 m L、Tw een 80質量濃度0.08 g/100 m L、乙酸鈉質量濃度0.5 g/100 m L、K2HPO4質量濃度0.2 g/100 m L。在此條件下,細菌素產量提高了216%,效價達到1 145 IU/m L。
[1] K?N IG H, FR?HLICH J. Lactic acid bacteria[M]//Biology of M icroorganisms on Grapes, in Must and in W ine. Berlin: Springer,2009: 3-29. DOI:10.1007/978-3-540-85463-0.
[2] AXELSSON L. Lactic acid bacteria: m icrobiological and functional aspects[M]//3rd. Lactic acid bacteria: classification and physiology.New York: Marcel Dekker Inc, 2004: 1-66.
[3] CALO-MATA P, ARLINDO S, BOEHME K, et al. Current app lications and future trends o f lactic acid bacteria and their bacteriocins for the biopreservation of aquatic food products[J]. Food and Bioprocess Technology, 2008, 1(1): 43-63. DOI:10.1007/s11947-007-0021-2.
[4] LEROY F, DE VUYST L. Lactic acid bacteria as functional starter cultures for the food fermentation industry[J]. Trends in Food Science &Technology, 2004, 15(2): 67-78. DOI:10.1016/j.tifs.2003.09.004.
[5] SAVADOGO A, OUATTARA A T C, BASSOLE H N I, et al.Bacteriocins and lactic acid bacteria: a m inireview[J]. A frican Journal of Biotechnology, 2006, 5(9): 678-683.
[6] ZACHAROF M P, LOVITT R W. Bacteriocins produced by lactic acid bacteria a review article[J]. Apcbee Procedia, 2012, 2: 50-56.DOI:10.1016/j.apcbee.2012.06.010.
[7] TWOMEY D, ROSS R P, RYAN M, et al. Lantibiotics produced by lactic acid bacteria: structure, function and applications[J]. Antonie Van Leeuw enhoek, 2002, 82(1/2/3/4): 165-185. DOI:10.1023/A:1020660321724.
[8] ROSSI F, PALLOTTA M L. Bacteriocin producing cultures: a sustainable way for food safety improvement and probiotics w ith additional health promoting eff ects[J]. International Journal of Medical and Biological Frontiers, 2016, 22(1): 59-87.
[9] AVONTS L, VAN UYTVEN E, DE VUYST L. Cell grow th and bacteriocin production of probiotic Lactobacillus strains in diff erent media[J]. International Dairy Journal, 2004, 14(11): 947-955.DOI:10.1016/j.idairyj.2004.04.003.
[10] CHEIGH C I, CHOI H J, PARK H, et al. In fluence o f grow th conditions on the p roduction o f a nisin-like bacteriocin by Lactococcus lactis subsp. lactis A 164 isolated from kim chi[J].Journal of Biotechnology, 2002, 95(3): 225-235. DOI:10.1016/S0168-1656(02)00010-X.
[11] TOMáS J, BRU E, W IESE B, et al. Influence of pH, temperature and culture media on the grow th and bacteriocin production by vaginal Lactobacillus salivarius CRL 1328[J]. Journal of Applied M icrobio logy, 2002, 93(4): 714-724. DOI:10.1046/j.1365-2672.2002.01753.x.
[12] BHATTACHARYA S, DAS A. Study of physical and cu ltural parameters on the bacteriocins produced by lactic acid bacteria isolated from traditional Indian fermented foods[J]. American Journal of Food Technology, 2010, 5(2): 111-120. DOI:10.3923/ajft.2010.111.120.
[13] DELGADO A, BRITO D, PERES C, et al. Bacteriocin production by Lactobacillus pentosus B96 can be expressed as a function of temperature and NaCl concentration[J]. Food M icrobiology, 2005,22(6): 521-528. DOI:10.1016/j.fm.2004.11.015.
[14] TABBENE O, SLIMENE I B, DJEBALI K, et al. Optim ization of medium composition for the production of antim icrobial activity by Bacillus subtilis B38[J]. Biotechnology Progress, 2009, 25(5): 1267-1274. DOI:10.1002/btpr.202.
[15] DE VUYST L, LEROY F. Bacteriocins from lactic acid bacteria:production, purification, and food app lications[J]. Journal of Molecular M icrobiology and Biotechnology, 2007, 13(4): 194-199.DOI:10.1159/000104752.
[16] MALHEIROS P S, SANT’ANNA V, TODOROV S D, et al.Optim ization of grow th and bacteriocin production by Lactobacillus sakei subsp. sakei2a[J]. Brazilian Journal of M icrobiology, 2015,46(3): 825-834. DOI:10.1590/S1517-838246320140279.
[17] SOOMRO A H, MASUD T, ANWAAR K. Role of lactic acid bacteria(LAB) in food preservation and human health-a review[J]. Pakistan Journal of Nutrition, 2002, 1(1): 20-24.
[18] 張艾青, 劉書亮, 敖靈. 產廣譜細菌素乳酸菌的篩選和鑒定[J].微生物學通報, 2007, 34(4): 753-756. DOI:10.13344/j.m icrobiol.china.2007.04.014.
[19] 劉書亮, 張艾青, 田剛. 植物乳桿菌P158的生長曲線及其細菌素的特性[J]. 核農學報, 2009, 23(6): 1021-1025; 1035.
[20] 羅佩文, 劉書亮, 朱永清, 等. 植物乳桿菌SJ35的抑菌特性及復合保鮮劑配制[J]. 食品與機械, 32(7): 112-116; 201. DOI:10.13652/j.issn.1003-5788.2016.07.027.
[21] ZHOU K, ZENG Y T, HAN X F, et al. M odelling grow th and bacteriocin production by Lactobacillus plantarum BC-25 in response to temperature and pH in batch fermentation[J]. Applied Biochem istry and Biotechnology, 2015, 176(6): 1627-1637. DOI:10.1007/s12010-015-1666-3.
[22] TURGIS M, VU K D, M ILLETTE M, et al. Influence of environmental factors on bacteriocin production by human isolates of Lactococcus lactis MM 19 and Pediococcus acidilactici MM 33[J]. Probiotics and Antim icrobial Proteins, 2016, 8(1): 53-59. DOI:10.1007/s12602-015-9204-8.
[23] 陳蕓蕓, 陸兆新, 盧靜, 等. 植物乳桿菌fmb10產細菌素發酵條件的優化[J]. 食品科學, 2015, 36(17): 140-145. DOI:10.7506/spkx1002-6630-201517027.
[24] PAL A, RAMANA K V, BAWA A S. Simplifi cation and optim ization of deManRogosa Sharpe (MRS) medium for enhanced production of bacteriocin by Weissella paramesenteroides DFR-8[J]. Journal of Food Science and Technology, 2010, 47(3): 258-265. DOI:10.1007/s13197-010-0040-2.
[25] O’SULLIVAN L, ROSS R P, HILL C. Potential of bacteriocinproducing lactic acid bacteria for improvements in food safety and quality[J]. Biochim ie, 2002, 84(5): 593-604. DOI:10.1016/S0300-9084(02)01457-8.
[26] ABO-AMER A E. Optim ization of bacteriocin production by Lactobacillus acidophilus AA 11, a strain isolated from Egyptian cheese[J]. Annals o f M ic robio logy, 2011, 61(3): 445-452.DOI:10.1007/s13213-010-0157-6.
[27] ANTHONY T, RAJESH T, KAYALV IZHI N, et al. In fluence of medium com ponents and fermentation conditions on the production of bacteriocin(s) by Bacillus licheniformis AnBa9[J].Bioresource Technology, 2009, 100(2): 872-877. DOI:10.1016/j.biortech.2008.07.027.
[28] RAZA W, YANG X, WU H, et al. Evaluation of metal ions (Zn2+,Fe3+and M g2+) effect on the p roduc tion o f fusaricidin-type antifungal com pounds by Paenibacillus po lymyxa SQR-21[J].Bioresource Technology, 2010, 101(23): 9264-9271. DOI:10.1016/j.biortech.2010.07.052.
[29] COLLADO M C, HERNáNDEZ M, SANZ Y. Production of bacteriocin-like inhibitory compounds by human fecal Bifidobacterium strains[J]. Journal of Food Protection, 2005, 68(5): 1034-1040.DOI:10.4315/0362-028X-68.5.1034.
[30] PARENTE E, RICCIARDI A. Production, recovery and purifi cation of bacteriocins from lactic acid bacteria[J]. Applied M icrobiology and Biotechnology, 1999, 52(5): 628-638. DOI:10.1007/s002530051570.
Optimization of Medium and Culture Conditions for Bacteriocin Production by Lactobacillus plantarum P158
XU Longqian1, HU Kaidi1, ZHANG Aiqing1, ZHOU Kang1,2, LIU Shuliang1,2,*
(1. College of Food Science, Sichuan Agricultural University,Ya’an 625014, China;2. Institute of Food Processing and Safety, Sichuan Agricultural University, Ya’an 625014, China)
To increase the yield of bacteriocin produced by Lactobacillus plantarum P158, the medium and culture conditions for this strain were optimized using one-factor-at-a-time and orthogonal array design methods. The bacteriostatic activity of bacteriocin against Micrococcus luteus and Pseudomonas aeruginosa and pH were used as response variables.Results showed that MRS was the most suitable for bacteriocin production among five culture media for lactic acid bacteria.The optimal culture conditions were obtained as follow s: inoculum amount, 3% (V/V); initial pH, 6.0; and static culture at 34 ℃ for 42 h; the optimum medium was composed of glucose 2 g/100 m L, yeast extract 2 g/100 m L, soy peptone 1.5 g/100 m L, MgSO40.058 g/100 m L, MnSO40.025 g/100 m L, FeSO40.02 g/100 m L, Tween 80 0.08 g/100 m L, sodium acetate 0.5 g/100 m L, and K2HPO40.2 g/100 m L. Under these conditions, bacteriocin titer was 1 145 IU/m L, which was 216% higher than that (362 IU/m L) before optimization.
Lactobacillus plantarum; bacteriocin; medium; culture conditions
10.7506/spkx1002-6630-201722017
TS201.6
A
1002-6630(2017)22-0109-08引文格式:
徐瓏倩, 胡凱弟, 張艾青, 等. 植物乳桿菌P158產細菌素培養基及培養條件的優化[J]. 食品科學, 2017, 38(22): 109-116.
DOI:10.7506/spkx1002-6630-201722017. http://www.spkx.net.cn
XU Longqian, HU Kaidi, ZHANG Aiqing, et al. Optimization of medium and culture conditions for bacteriocin production by Lactobacillus plantarum P158[J]. Food Science, 2017, 38(22): 109-116. (in Chinese w ith English abstract) DOI:10.7506/spkx1002-6630-201722017. http://www.spkx.net.cn
2017-01-17
四川省農業科技成果轉化資金項目(14NZ0012);四川省科技支撐計劃項目(2015FZ0001)
徐瓏倩(1996—),女,學士,研究方向為食品微生物。E-mail:540922828@qq.com
*通信作者:劉書亮(1968—),男,教授,博士,研究方向為食品微生物與發酵工程。E-mail:lsliang999@163.com
猜你喜歡
中學生數理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
中學生數理化·中考版(2020年10期)2020-11-27 01:59:48
中國生殖健康(2019年2期)2019-08-23 08:12:08
產品可靠性報告(2017年7期)2017-09-05 09:49:12
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
汽車觀察(2016年3期)2016-02-28 13:16:26
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
