黃河三角洲刺槐根際與非根際細菌結構及多樣性*
2017-11-01 09:21:30丁新景敬如巖黃雅麗陳博杰馬風云
土壤學報 2017年5期
關鍵詞:差異
丁新景 敬如巖 黃雅麗 陳博杰 馬風云?
(1 山東農業大學林學院,山東農業大學生態與環境實驗室,山東泰安 271018)
(2 中國科學院西北生態環境資源研究院,蘭州 730000)
(3 中國科學院大學,北京 100049)
黃河三角洲刺槐根際與非根際細菌結構及多樣性*
丁新景1,2,3敬如巖1黃雅麗1陳博杰1馬風云1?
(1 山東農業大學林學院,山東農業大學生態與環境實驗室,山東泰安 271018)
(2 中國科學院西北生態環境資源研究院,蘭州 730000)
(3 中國科學院大學,北京 100049)
為精確分析黃河三角洲刺槐根際與非根際土壤細菌群落定殖情況,本研究采用高通量測序方法對刺槐根際與非根際土壤細菌結構及多樣性進行了研究。研究表明,根際土壤細菌共有36門214屬,非根際土壤細菌共有33門153屬。變形菌門(Proteobacteria)、放線菌門(Actinobacteria)、酸桿菌門(Acidobacteria)細菌豐度超過15%,是根際與非根際土壤中的優勢菌落。根際與非根際土壤中酸桿菌門、硝化螺旋菌門(Nitrospirae)豐度差異顯著。根際土壤中紅游動菌屬(Rhodoplanes)、溶桿菌屬(Lysobacter)、熱單胞菌屬(Thermomonas)、鏈霉菌屬(Streptomyces)及非根際土壤中紅游動菌屬、溶桿菌屬、鏈霉菌屬、Kaistobacter細菌豐度超過4%。根際土壤中固氮菌豐度顯著高于非根際土壤,解磷、解鉀細菌豐度差異不顯著。根際與非根際土壤細菌Chao豐富度分別為2 054、2 376,差異顯著。根際與非根際土壤細菌之間的權重(Weighted Unifrac)距離在0.12~0.25之間。綜上所述,黃河三角洲刺槐根際與非根際土壤細菌結構具有一定差異,多樣性差異顯著。
黃河三角洲;刺槐;根際;細菌;多樣性
微生物是土壤生態系統的重要組成部分,參與土壤中很多生理生化反應,在土壤生態系統的物質轉換、能量流動、信息傳遞中發揮著重要作用[1]。土壤微生物對植物生長具有重要影響。微生物作為植物有機殘體的主要分解者能夠通過土壤中營養元素的周轉,調節植物養分的供應,影響植物的生長、資源分配和化學組成。此外,微生物可以將土壤中植物不能直接利用的N、P轉化為有效態,因此在微生物多樣性較高的土壤中植物可以吸收更多的營養物質,對外界的脅迫具有更強的恢復能力[1]。如微生物多樣性較低土壤中植物對熱脅迫和銅脅迫的恢復力也最低,且當多樣性下降時某些特定的功能也受到極大影響[1-2]。土壤微生物中細菌數量最大、種類最多,生物學特性各異,繁殖迅速,對土壤肥力的形成、植物營養的轉化有極其重要的作用[3]。植物根際土壤是根系與細菌相互作用的媒介,在根系與細菌的共同作用下形成的復雜微生態系統[4-5]。植物根系代謝產生分泌物,其中一些物質可直接影響根際土壤細菌生長、發育和繁殖,使根際細菌數量、種類不同于非根際土壤,形成根際效應[6-7]。根際土壤細菌對植物的生長具有促進作用,如部分細菌與豆科植物根系可以聯合固氮,也有部分細菌可以通過產生激素影響植物生長[8]。根際土壤細菌的研究有利于進一步認識植物根系-細菌的共生體系協同進化機制[9]。
黃河三角洲是我國重要的濕地分布區,目前正處于快速開發建設時期,如何在經濟快速發展的同時保持良好的生態環境,實現經濟的可持續發展,是一個重要的研究課題。為改善和保護該地區的生態環境,黃河三角洲地區自20世紀80年代以來營造了大量防護林,其中刺槐林栽培面積較大,達5 000 hm2,這些防護林在該地區防風固沙、水土保持、土壤改良等方面發揮著重要的作用[10]。鑒于細菌在土壤各種生態過程中具有的重要作用,本研究選擇該地區刺槐人工林為研究對象,研究林地根際與非根際土壤細菌群落結構及多樣性差異,為人工林的科學管理以及進一步研究黃河三角洲生態系統提供可參考依據。
1 材料與方法
1.1 研究區概況
研究區位于山東省東營市河口區孤島鎮,東、北兩側臨渤海,屬暖溫帶半濕潤季風氣候。該地年平均氣溫為12.8 ℃,年均無霜期為234 d,年均日照時間約2 728 h,年均降水量為690.5 mm,全年降水分布不均,夏季降水占全年降水量的69%,超過10℃有效積溫約4 300 ℃左右[10]。
人工林為1985年營建,株行距為3 m×3 m。林木主要有刺槐(Robinia pseudoacacia)、絨毛白蠟(Fraxinus velutina)、臭椿(Ailanthus altissima)、榆樹(Ulmus pumila)、楊樹(Populs)、國槐(Sophora japonica)等。林下生長有茅草(Imperata cylindica)、荻(Triarrherca saccharifora)等草本植物。
1.2 樣品采集與處理
2014年11月在刺槐純林中設3個20 m×20 m標準地。每標準地內選取1株標準木,距樹干基部0.5 m處設東西南北4個取樣點,采集直徑小于0.2 cm的刺槐細根,用抖落法獲取細根上粘附的土壤作為根際土壤,將每個標準地獲取的根際土壤分別混勻;另外在各取樣點5~20 cm土層采集非根際土壤。將每標準地根際與非土壤分為2份,一份裝入已消毒的密封塑料袋中,液氮保存帶回實驗室,并在-70℃冷凍條件下備用;1份土樣風干過0.20 mm篩,用于測定土壤理化性質[10]。另用環刀法取樣測定土壤含水量。
1.3 土壤微生物DNA提取、測序、處理
采用CTAB方法提取樣本基因組DNA[11],之后利用瓊脂糖凝膠電泳、OD280檢測DNA的純度和濃度。在離心管中將無菌水稀釋樣品至1 ng μl-1,以稀釋后基因組DNA為模板,根據測序區域的選擇,使用帶條碼(Barcode)的特異引物,采用高效和高保真酶進行聚合酶鏈反應(PCR)。引物對應區域:16S V4區引物為515F-806R。用New England Biolabs公司的建庫試劑盒進行文庫的構建,構建好的文庫經過Qubit定量和文庫檢測合格后,使用MiSeq進行上機測序。
Illumina MiSeq(Illumina高通量測序)測序平臺根據Barcode序列將下機數據拆分為不同樣品數據,截去Barcode序列和PCR擴增引物序列;將拆分的數據使用FLASH對每個樣品的片段(reads)進行拼接,得到的拼接序列為原始標簽(Raw Tags);Raw Tags經過更嚴格過濾處理后[12],得到高質量的標簽數據(Clean Tags):Raw Tags從連續低質量值(默認質量閾值為≤ 3)堿基數達到設定長度(默認長度值為3)的第一個低質量堿基位點截斷;經過截取后得到的標簽(Tags)數據集,進一步過濾掉其中連續高質量堿基長度小于Tags長度75%的Tags[13]。經過以上處理后得到的Tags序列與數據庫(UCHIME Algorithm)進行比對,檢測嵌合體序列并最終去除其中的嵌合體序列[14]。
1.4 物種注釋與豐度計算
用Uparse軟件對所有樣品的全部有效序列(Effective Tags)聚類[15],默認97%的一致性將序列聚類成為OTUs(群落聚類),Uparse構建OTUs時會選取代表性序列(依據其算法原則,篩選OTUs中出現頻數最高的序列),將這些代表性序列集合用RDP Classifier與GreenGene數據庫進行物種注釋分析。結合物種組成信息結果,得到在該樣品中每個物種豐度。
1.5 數據處理
根據每個樣本文庫的OTUs豐度信息,利用QIIME(Version1.7.0)軟件計算樣品的α多樣性值,包括物種數、Chao指數、香農指數(Shannon index,H)、辛普森指數(Simpson index,D):
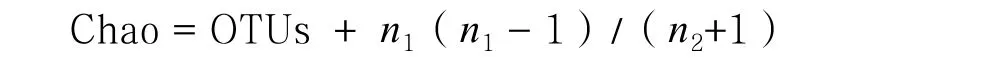
式中, OTUs為實際觀測到的 OTU數目;n1為只含有一條序列OTU數目;n2為只含有兩條序列的 OTU數目;
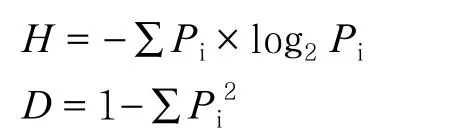
式中,Pi=ni/N,表明第i個物種的相對多度;ni為含有i條序列的 OTU數目;N為所有序列數[16]。
利用QIIME軟件分析根際和非根際土壤樣品差異性[15],即Weighted Unifrac距離。應用SPSS 22.0 軟件完成統計分析,各處理間數據采用方差分析(ANOVA),以 Duncan法檢測差異顯著性,差異水平p<0.05。
2 結 果
2.1 根際與非根際土壤理化性質
刺槐林根際與非根際土壤理化性質見表1。刺槐根際與非根際土壤含水量、電導率、pH、速效鉀、硝態氮含量差異不顯著。根際土壤中有效磷、堿解氮、有機質、可溶性有機碳、可溶性有機氮含量顯著高于非根際土壤,分別是非根際土壤的88.76%、71.19%、88.50%、88.50%、90.03%。
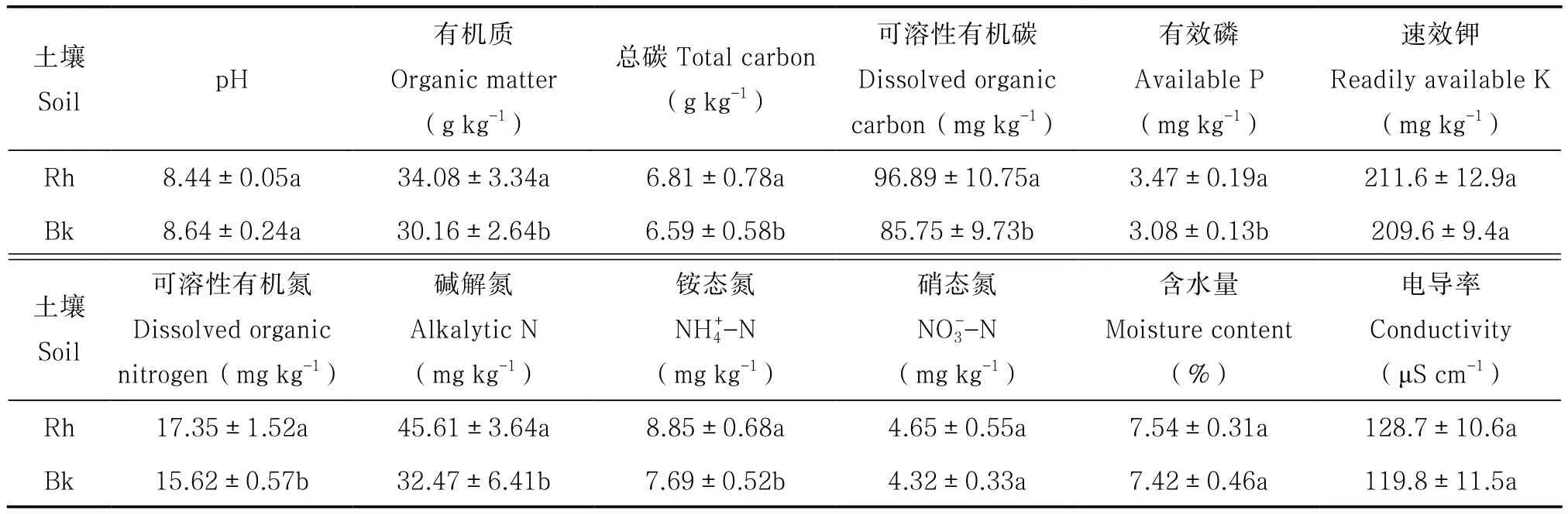
表1 刺槐林土壤理化性質Table 1 Physical and chemical properties of the soil under Robinia pseudoacacia forest
2.2 根際與非根際土壤細菌測序
經測定,黃河三角洲刺槐根際土壤3次重復分別獲得42 341、54 992、51 601條有效序列,非根際土壤3次重復分別獲得38 657、23 353、54 813條有效序列。對獲得的序列進行隨機抽樣的方法,以抽到的序列數與其所代表的OTU數目構建稀釋性曲線,如圖1,從圖中可以看出,12 360條序列時根際與非根際土壤序列稀釋性曲線均趨于平坦,更多的測序數據對發現新的OTU貢獻率較小,表明測序數據合理。
2.3 根際與非根際土壤細菌結構組成

圖1 根際與非根際土壤細菌稀疏性曲線分析Fig. 1 Sparsity curve analysis of OTUs of rhizosphere and bulk soil
根際與非根際土壤細菌分別有36門和33門,其中分別有9門和8門細菌豐度超過1%(圖2),豐度總和分別為97.37%和96.15%。變形菌門(Proteobacteria)、放線菌門(Actinobacteria)、酸桿菌門(Acidobacteria)3門細菌在根際與非根際土壤中豐度超過15%,是根際與非根際土壤中的優勢菌落。變形菌門細菌在根際土壤中豐度最高,為33.30%,顯著高于非根際土壤中的豐度。變形菌門細菌共有5綱,根際土壤中Betaproteobacteria、Alphaproteobacteria、Gammaproteobacteria 3綱細菌豐度分別為12.31%、9.46%、7.83%,顯著高于非根際;Deltaproteobacteria細菌在根際與非根際土壤中豐度分別為3.67%、4.04%,差異不顯著;根際與非根際中Epsilonproteobacteria細菌豐度低于1%。非根際土壤中酸桿菌門豐度最高,為29.29%,根際土壤中豐度為20.64%,差異顯著(p<0.05)。綠彎菌門(Chloroflexi)、芽單胞菌門(Gemmatimonadetes)、浮游菌門(Planctomycetes)、硝化螺旋菌門(Nitrospirae)、疣微菌門(Verrucomicrobia)5門細菌在根際與非根際土壤中豐度介于1.96%~6.82%,其中硝化螺旋菌門細菌在根際土壤豐度僅為2.39%,而非根際土壤中豐度為6.82%,差異顯著(p<0.05)。
根際與非根際土壤中細菌分別有214屬和153屬,豐度超過1%的屬分別有29屬和32屬(圖3),豐度總和分別為76.95%和80.08%。紅游動菌屬(Rhodoplanes)、溶桿菌屬(Lysobacter)、熱單胞菌屬(T h e r m o m o n a s)、鏈霉菌屬(Streptomyces)4屬細菌在根際土壤中豐度超過4%。紅游動菌屬、溶桿菌屬、鏈霉菌屬、Kaistobacter4屬細菌在非根際土壤中豐度超過4%。許多屬細菌在根際與非根際土壤中豐度差異顯著(p<0.05)。如新鞘氨醇桿菌屬(Novosphingobium)、氣微菌屬(Aeromicrobium)、Limnohabitans、海藻球菌屬(Phycicoccus)4屬細菌在根際土壤中豐度顯著高于非根際土壤(p<0.05),其中Limnohabitans、海藻球菌屬細菌在根際土壤中豐度超過1%,在非根際土壤中未檢測到。Kaistobacter、Candidatus Nitrososphaera、DA101、Ramlibacter、Methylibium5屬細菌在非根際土壤中豐度顯著高于根際土壤(p<0.05),其中Ramlibacter、Methylibium在非根際土壤中的豐度超過1%,但在根際土壤中未檢測到。
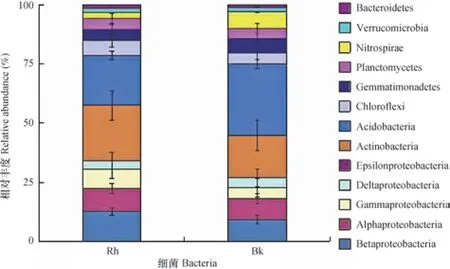
圖2 根際與非根際土壤細菌門水平相對豐度Fig. 2 Bacterial abundance of rhizosphere and bulk soil at the phylum level
2.4 根際與非根際土壤細菌功能組成
根際與非根際土壤中具有固氮功能的細菌分別有17屬、15屬(表2),根際土壤中具有固氮功能的細菌豐度是非根際土壤中的1.21倍,差異顯著(p<0.05),其中短波單胞菌屬(Brevundimonas)、成對桿菌屬(Dyadobacter)、硫桿菌屬(Thiobacillus)3屬細菌僅在根際土壤中檢測到,弗蘭克氏菌屬(Frankia)細菌僅在非根際土壤中檢測到。根際與非根際土壤中具有解磷功能的細菌分別有8屬、7屬,具有解鉀功能的細菌分別有5屬、4屬,解磷、解鉀細菌豐度差異不顯著(p>0.05)。窄食單胞菌屬(Stenotrophomonas)、芽孢桿菌屬(Bacillus)、布克氏菌屬(Burkholderia)3屬細菌具有減少植株病害的作用,3屬細菌在非根際土壤細菌豐度總和是根際土壤的3.93倍,差異顯著(p<0.05)。根際與非根際土壤中還分布有植物致病菌假黃單胞菌屬(Pseudoxanthomonas)、螺桿菌屬(Helicobacter)。

圖3 根際與非根際土壤細菌屬水平相對豐度Fig. 3 Bacterial abundance of Rhizosphere and bulk soil at the generic level

表2 根際與非根際土壤細菌功能組成Table 2 Bacterial function composition of rhizosphere and bulk soil
2.5 根際與非根際土壤細菌群落多樣性
97%序列相似度被認為是種(species)的分類水平,本研究以97%相似度水平歸并OTUs計算根際與非根際土壤細菌的α多樣性。從表3得知,非根際土壤所測得的細菌物種數、香農指數均高于根際土壤,差異不顯著(p>0.05);非根際土壤細菌Chao豐富度為2 376,顯著高于根際;根際與非根際土壤細菌辛普森指數差異不顯著(p>0.05)。

表3 刺槐根際與非根際土壤細菌多樣性分析Table 3 Bacterial diversity index of the rhizosphere and bulk soil under Robinia pseudoacacia
2.6 根際與非根際土壤細菌差異
Weighted Unifrac距離可以用來作為衡量兩個樣品間的相異系數,其值越小,表示這兩個樣品在物種多樣性方面存在的差異越小[17]。刺槐根際土壤細菌之間的Weighted Unifrac距離在0.17~0.25之間;刺槐根際土壤細菌之間的Weighted Unifrac距離顯著高于非根際(p<0.05);根際與非根際土壤細菌之間Weighted Unifrac距離整體偏大(表4)。非根際土壤細菌之間差異較小,根際土壤細菌之間差異較大。
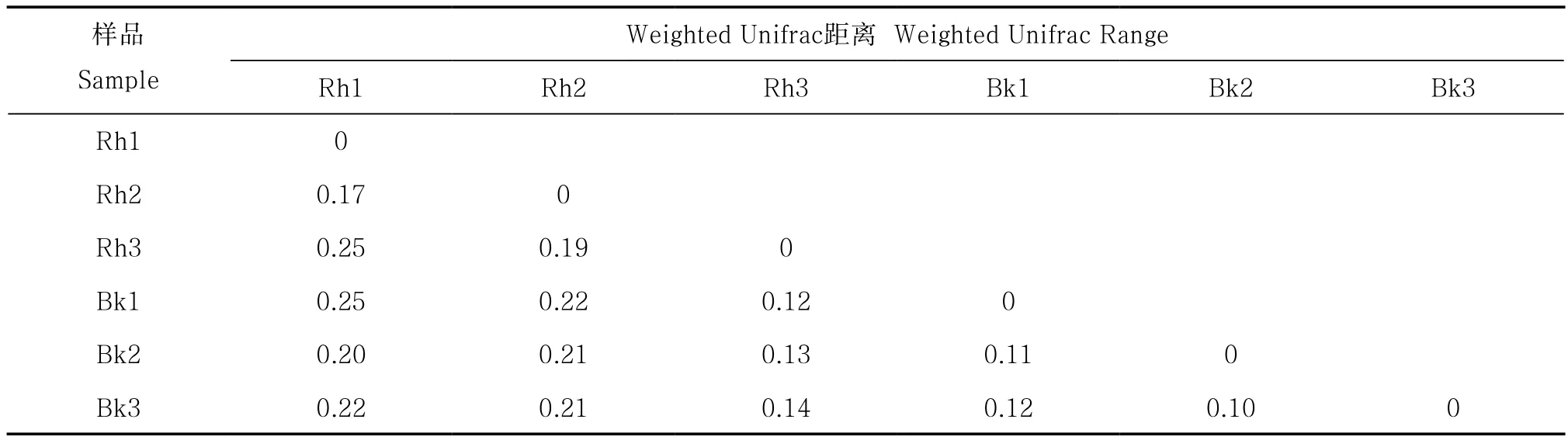
表4 根際與非根際土壤細菌Weighted Unifrac距離Table 4 Bacterial Weighted Unifrac Range between rhizosphere and bulk soil
3 討 論
3.1 根際與非根際土壤細菌群落結構
黃河三角洲刺槐林土壤中,變形菌門(Proteobacteria)、放線菌門(Actinobacteria)、酸桿菌門3門細菌土壤中豐度均超過15%,是土壤中的優勢群落。王伏偉[15]、柳春林[16]、楊菁[18]等對砂姜黑土、赤紅壤、砂壤細菌群落研究后發現變形菌門、放線菌門、酸桿菌門3門細菌是上述3種土壤中的優勢群落,杜瀅鑫等[19]研究表明變形菌門、放線菌門、酸桿菌門3門細菌在大慶鹽堿地中同樣占據優勢,與本研究結果相同。說明變形菌門、放線菌門、酸桿菌門3門細菌適應性廣,能夠存在于各種不同的土壤中,環境變化對這3門細菌分布影響不大[15]。
門水平上變形菌門、酸桿菌門、硝化螺旋菌門3門細菌在根際與非根際土壤中豐度差異顯著;屬水平上新鞘氨醇桿菌屬(Novosphingobium)、氣微菌屬(Aeromicrobium)、Limnohabitans、海藻球菌屬(Phycicoccus)、Kaistobacter、Candidatus、Nitrososphaera、DA101、Ramlibacter、Methylibium9屬細菌在根際與非根際土壤中相對豐度差異顯著。汪其同等[5]研究表明楊樹根際和非根際土壤中8屬相對豐度較高的細菌豐度存在顯著差異。細菌在根際與非根際土壤中豐度差異可能是由于植物根系生長代謝及根系分泌物差異導致根際與非根際土壤的透氣性、濕度、pH、土壤顆粒大小、土壤緊實度及土壤N、P、K、有機質含量等不同,不同理化性質的土壤中細菌類群不同,因此根際與非根際土壤細菌群落結構存在差異[20]。
3.2 根系對細菌群落多樣性的影響
一般由于植株根系代謝為根際土壤細菌提供較多的碳源,使得根際土壤細菌數量、多樣性高于非根際,如楊樹人工林根際細菌多樣性高于非根際[5];本溪山櫻根際與非根際解磷細菌群落存在明顯的根際效應[21]。黃河三角洲刺槐根際土壤細菌Chao指數為2 054,而非根際為2 376,根際土壤細菌多樣性低于非根際土壤。這應該與土樣采集時間有關。試驗土樣采于11月,該地區11月份降水較少,土壤中水分蒸發強烈,刺槐受到干旱脅迫,根系調整自身根系分泌物的組成和數量來積極響應土壤環境的變化[22]。根系分泌物對根際微生物具有一定的選擇性,干旱脅迫促使植物分泌物增加,植物根系對根際細菌的選擇性增強,使部分細菌豐度升高,其他細菌豐度降低,根際細菌多樣性低于非根際[5]。因此根際土壤細菌多樣性降低可以作為植物對干旱脅迫響應的一種表現[23]。
3.3 土壤理化性質對細菌群落結構的影響
許多研究表明酸桿菌門在酸性土壤中豐度較高,如柳春林等[16]研究發現鼎湖山森林土壤中酸桿菌門豐度介于53. 3%~67. 8%,隋心等[24]在三江平原研究發現土壤中酸桿菌門豐度為53%。酸桿菌門是嗜酸性細菌,酸性土壤有利于酸桿菌門的生長[25]。鼎湖山與三江平原土壤為酸性,適于酸桿菌門的生長,而黃河三角洲刺槐根際與非根際土壤中豐度分別為20.64%和29.29%,豐度偏低,可能是堿性土壤不利于酸桿菌門生長。雖然黃河三角洲刺槐林土壤中酸桿菌門細菌豐度較低,但土壤細菌多樣性(Chao指數2054~2 376)卻高于隋心等[24](Chao指數709~745)、汪其同等[5](Chao指數1931~2260)研究的土壤,可能是堿性環境中酸桿菌門豐度較低,利于其他細菌的生長,從而提高了土壤的細菌多樣性。黃河三角洲刺槐林土壤中檢測出芽單胞菌門(Gemmatimonadetes),這個門的細菌在海南降香黃檀人工林土壤及鼎湖山森林土壤細菌中均未發現[16],三江平原小葉章土壤中芽單胞菌門豐度極低,僅1%[24],而芽單胞菌門在黃河三角洲刺槐根際與非根際中豐度較高,分別為4.67%和5.95%,這可能是堿性環境不利于酸桿菌門細菌生長從而使芽單胞菌門細菌豐度較高。
刺槐根際與非根際土壤含水量、電導率、pH、速效鉀含量差異不顯著,根際土壤堿解氮、有效磷、有機質含量顯著高于非根際。這可能是刺槐受干旱脅迫后根系分泌物增加引起的,根系分泌物中含有較多的低分子糖、多種氨基酸、有機酸、酮、酚和胺等物質,根系分泌物中氨基酸、有機酸能夠將土壤中無效態氮、磷轉化為有效態[6],因此刺槐根際土壤堿解氮、有效磷、有機質含量顯著高于非根際。根際土壤理化性質發生變化,增強了部分細菌競爭能力[5],如刺槐根際土壤中變形菌門、放線菌門等細菌豐度均顯著高于非根際。變形菌門、放線菌門等細菌對土壤資源消耗增長使土壤細菌群落競爭加劇,酸桿菌門、硝化螺旋菌門等細菌食物資源減少,豐度降低,根際土壤細菌多樣性低于非根際。
根際土壤為細菌提供了豐富的堿解氮、有效磷、有機質等營養物質,因此根際土壤中微生物數量多于非根際土壤[26],但由于根系分泌物對根際微生物具有一定的選擇性[5],因此刺槐根際土壤細菌多樣性小于非根際土壤。
4 結 論
黃河三角洲刺槐林根際土壤細菌豐度高于非根際土壤細菌。變形菌門、放線菌門、酸桿菌門細菌是根際與非根際土壤中的共同優勢菌落。紅游動菌屬、溶桿菌屬、熱單胞菌屬、鏈霉菌屬為根際土壤優勢群落,紅游動菌屬、溶桿菌屬、鏈霉菌屬、
Kaistobacter細菌為非根際土壤優勢群落。根際土壤中固氮菌豐度顯著高于非根際土壤,解磷、解鉀細菌豐度差異不顯著。根際與非根際土壤細菌多樣性差異顯著。非根際土壤細菌之間物種多樣性差異較小,根際土壤細菌之間差異較大。
[1] Zelles L. Fatty acid patterns of phospholipids and lipopolysaccharides in the characterisation of microbial communities in soil:A review. Biology and Fertility of Soils,1999,29:111—129
[2] Griffiths B S,Ritz K,Bardgett R D,et al. Ecosystem response of pasture soil communities to fumigationinduced microbial diversity reductions:An examination of the biodiversity-ecosystem function relationship.Oikos,2000,90(2):279—294
[3] Kennedy A C. Bacterial diversity in agroecosystems.Agriculture Ecosystems and Environment,1999,74(1):65—76
[4] Ryan P R,Jones D L,Delhaize E. Function and mechanism of organic anion exudation from plant roots.Annual Review of Plant Physiology and Plant Molecular Biology,2001,52:527—560
[5] 汪其同,朱婉芮,劉夢玲,等. 基于高通量測序的楊樹人工林根際和非根際細菌群落結構比較. 應用與環境生物學報,2015,21(5):967—973 Wang Q T,Zhu W R,Liu M L,et al. Comparison on bacterial community of rhizosphere and bulk soil of poplar plantation based on pyrosequencing(In Chinese). Chinese Journal of Applied and Environmental Biology,2015,21(5):967—973
[6] Wardle D A. A comparative assessment of factors which influence microbial biomass carbon and nitrogen levels in soil. Biological Reviews,1992,67(3):321—358
[7] Prosser J. Molecular and functional eliversity in soil micro-organisms. Plant and Soil,2002,244(1):9—17
[8] Schippers B,Bakker A W,Bakker P. Interactions of deleterious and beneficial rhizosphere microorganisms and the effect of cropping practices. Annual Review of Phytopathology,2003,25(1):339—358
[9] Berg G,Grube M,Schloter M,et al. Unraveling the plant microbiome:Looking back and future perspectives. Frontiers in Microbiology,2014,5,DOI:10.3389/fmicb.2014.00148
[10] 李萍,趙庚星,高明秀,等. 黃河三角洲土壤含水量狀況的高光譜估測與遙感反演. 土壤學報,2015,52(6):1262—1272 Li P,Zhao G X,Gao M X,et al. Hyperspectral estimation and remote sensing retrieval of soil water regime in the Yellow River Delta( In Chinese). Acta Pedologica Sinica,2015,52(6):1262—1272
[11] 劉璐,何尋陽,謝強,等. 桂西北喀斯特常見豆科植物根瘤菌的遺傳多樣. 應用生態學報,2015,26(12):3663—3669 Liu L,He X Y,Xie Q,et al. Genetic diversity of rhizobia isolated from common legumes in the Karst area Northwest Guangxi(In Chinese). Chinese Journal of Applied Ecology,2015,26(12):3663—3669
[12] Bokulich N A,Subramanian S,Faith J J,et al.Quality-filtering vastly improves diversity estimates from Illumina amplicon sequencing. Nature Methods,2013,10(1):57—59
[13] Caporaso J G,Kuczynski J,Stombaugh J,et al.QIIME allows analysis of high-throughput community sequencing data. Nature Methods,2010,7(5):335—336
[14] 游偲,張立猛,計思貴,等. 枯草芽孢桿菌菌劑對煙草根際土壤細菌群落的影響. 應用生態學報,2014,25(11):3323—3330 You C,Zhang L M,Ji S G,et al. Impact of biocontrol agent Bacillus subtilis on bacterial communities in tobacco rhizospheric soil(In Chinese). Chinese Journal of Applied Ecology,2014,25(11):3323—3330
[15] 王伏偉,王曉波,李金才,等. 施肥及秸稈還田對砂姜黑土細菌群落的影響. 中國生態農業學報,2015,23(10):1302—1311 Wang F W,Wang X B,Li J C,et al. Effects of fertilization and straw incorporation on bacterial communities in lime concretion black soil(In Chinese). Chinese Journal of Eco-Agriculture,2015,23(10):1302—1311
[16] 柳春林,左偉英,趙增陽,等. 鼎湖山不同演替階段森林土壤細菌多樣性. 微生物學報,2012,52(12):1489—1496 Liu C L,Zuo W Y,Zhao Z Y,et al. Bacterial diversity of different successional stage forest soils in Dinghushan( In Chinese). Acta Microbiologica Sinica,2012,52(12):1489—1496
[17] Li T,Long M,Gatesoupe F J,et al. Comparative analysis of the intestinal bacterial communities in different species of carp by pyrosequencing. Microbial Ecology,2014,69(1):25—36
[18] 楊菁,周國英,田媛媛,等. 降香黃檀不同混交林土壤細菌多樣性差異分析. 生態學報,2015,35(24):8117—8127 Yang J,Zhou G Y,Tian Y Y,et al. Differential analysis of soil bacteria diversity in different mixed forests ofDalbergia odorifera(In Chinese). Acta Ecologica Sinica,2015,35(24):8117—8127
[19] 杜瀅鑫,謝寶明,蔡洪生,等. 大慶鹽堿地九種植物根際土壤微生物群落結構和功能多樣性. 生態學報,2016,36(3):740—747 Du Y X,Xie B M,Cai H S,et al. Structural and functional diversity of rhizosphere microbial community of nine plant species in the Daqing Saline-alkali soil region(In Chinese). Acta Ecologica Sinica,2016,36(3):740—747
[20] Steenwerth K L,Jackson L E,Calderon F J,et al.Soil microbial community composition and land use history in cultivated and grassland ecosystems of coastal California. Soil Biology & Biochemistry,2002,34(11):1599—1611
[21] 于翠,呂德國,秦嗣軍,等. 本溪山櫻根際與非根際解磷細菌群落結構及動態變化. 應用生態學報,2006,17(12):2381—2384 Yu C,Lü D G,Qin S J,et al. Community structure and its dynam ics of phosphobacteria inCerasus sachalinensisrhizosphere and non-rhizosphere(In Chinese). Acta Ecologica Sinica,2006,17(12):2381—2384
[22] Walker T S,Bais H P,Grotewold E,et al. Root exudation and rhizosphere biology. Plant Physiology,2003,132(1):44—51
[23] 吳林坤,林向民,林文雄. 根系分泌物介導下植物-土壤-微生物互作關系研究進展與展望. 植物生態學報,2014,38(3):298—310 Wu L K,Lin X M,Lin W X. Advances and perspective in research on plant-soil-microbe interactions mediated by root exudates( In Chinese). Chinese Journal of Plant Ecology,2014,38(3):298—310
[24] 隋心,張榮濤,鐘海秀,等. 利用高通量測序對三江平原小葉章濕地土壤細菌多樣性的研究. 土壤,2015,47(5):919—925 Sui X,Zhang R T,Zhong H X,et al. Study on bacterial diversity ofDeyeuxia angustifoliawetland by application of High-throughput sequencing technology in Sanjiang Plain(In Chinese). Soils,2015,47(5):919—925
[25] Schabereiter G C,Saiz J C,Pinar G,et al. Altamira cave Paleolithic paintings harbor partly unknown bacterial communities. FEMS Microbiology Letters,2002,211(1):7—11
Bacterial Structure and Diversity of Rhizosphere and Bulk Soil ofRobinia pseudoacaciaForests in Yellow River Delta
DING Xinjing1,2,3JING Ruyan1HUANG Yali1CHEN Bojie1MA Fengyun1?
(1Forestry College of Shandong Agricultural University,Laboratory of Ecology and Environment,Shandong Agricultural University,Taian,Shandong271018,China)
(2Northwest Institute of Eco-Environment and Resources,Chinese Academy of Sciences,Lanzhou730000,China)
(3University of Chinese Academy of Sciences,Beijing100049,China)
【Objective】 As one of the three major pedogenic river deltas in China and the one that develops the fastest,the Yellow River Delta is an important reserved land resource. Developing mixed forests has been becoming a major strategy to develop the Yellow River Delta,because mixed forests can not only regulate temperature but also improve soil fertility. To improve the ecological system of the Yellow River Delta,a number of forest plantations have been established since the 1950s.Robinia pseudoacaciais the dominant tree species in most of the forest plantations,which contribute significantly to conserving soil and water and improving the ecological system of the Delta. Soil microbial community in plant rhizosphere plays an important role in plant nutrition and plant health. This research aimed to accurately evaluate how soil bacterial communities colonize in rhizosphere and in bulk soils underRobinia pseudoacaciain the Yellow River Delta. 【Method】 With the aid of the Illumina MiSeq technology,this research analyzed structure and diversity of the bacterial communities in the rhizosphere soil and the bulk soil underRobinia pseudoacaciafor comparison. The differences in soil bacterial community between rhizosphere and bulk soil were expressed via the Weighted Unifrac Range of OTUs. 【Result】Results show that the bacterial community in the rhizosphere soil consisted of 214 genera of 36 phyla and that in the bulk soil 153 genera of 33 phyla. Proteobacteria,Actinobacteria and Acidobacteria were the dominant phyla of bacteria in both rhizosphere and bulk soils with relative abundance being more than 15%. Acidobacteria and Nitrospirae varied sharply in relative abundance between the rhizosphere soil and the bulk soil. Chloroflexi,Gemmatimonadetes,Planctomycetes,Nitrospirae,and Verrucomicrobia were the dominant taxa of bacteria with relative abundance being more than 1%.Rhodoplanes,Lysobacter,ThermomonasandStreptomycesin the rhizosphere soil andRhodoplanes,Lysobacter,StreptomycesandKaistobacterin the bulk soil all exceeded 4% in relative abundance. There were 29 and 32 genera of bacteria in the rhizosphere soil and 3 in the bulk soil being more than 1% in relative abundance,respectively. Significant differences were found in relative abundance ofNovosphingobium,Aeromicrobium,Limnohabitans,Phycicoccus,aistobacter,Candidatus Nitrososphaera,DA101,Ramlibacter,andMethylibiumbetween the rhizosphere soil and the bulk soil. Relative abundance ofAzotobacterwas significantly higher in the rhizosphere soil than in the bulk soil,while no significant difference was observed in relative abundance of potassium and phosphorus-dissolving bacteria between the two soils. Chao richness of bacterial community was 2 054 and 2 376 in the rhizosphere soil and in the bulk soils,respectively,showing a significant difference. Weighted Unifrac Range mof the soil bacteria between the rhizosphere and the bulk soil was 0.12~0.25. In this study,all the findings demonstrate that there are some differences in structure between the bacterial communities in the rhizosphere soil and in the bulk soil underRobinia pseudoacaciain Yellow River Delta. 【Conclusion】Significant differences do exist in relative abundance of Proteobacteria,Acidobacteria,and Nitrospirae at the phylum level,andNovosphingobium,Aeromicrobium,Limnohabitans,andPhycicoccusat the genus level between rhizosphere and bulk soils,and significant differences are also observed in bacterial diversity between rhizosphere and bulk soils. Analysis of bacteria 16S rRNA-bacteria dataset shows differernces in structure and diversity of soil bacterial community between the rhizosphere and bulk soila in the Yellow River Delta. But further research is still needed to get more detailed information about soil microorganisms.
Yellow River Delta;Robinia pseudoacacia;Rhizosphere;Bacteria;Diversity
S154.36;S792
A
10.11766/trxb201703230510
* 國家自然科學基金項目(30970499)和世界銀行貸款項目( SEAP-KY-2)共同資助 Supported by the National Natural Science Foundation of China(No. 30970499)and the World Bank Loan Project(No. SEAP-KY-2)
? 通訊作者 Corresponding author:馬風云,副教授,主要研究方向為生態恢復和重建研究。E-mail:sdmfy@sdau.edu.cn
丁新景(1990—),男,山東菏澤人,碩士研究生,主要從事林業生態工程研究。Tel:18854889053;E-mail:18854889053@163.com
2017-03-23;
2017-05-05;優先數字出版日期(www.cnki.net):2017-06-20
(責任編輯:盧 萍)
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現代語文(2016年21期)2016-05-25 13:13:50
