耐高溫β-甘露聚糖酶畢赤酵母菌M27-8發(fā)酵條件優(yōu)化
2017-10-21 07:22:48黃魁英夏楓耿陳舒潔胡海艷
中國(guó)飼料 2017年19期
熊 進(jìn),黃魁英,夏楓耿,陳舒潔,胡海艷
(廣州市微生物研究所,廣東廣州 510663)
耐高溫β-甘露聚糖酶畢赤酵母菌M27-8發(fā)酵條件優(yōu)化
熊 進(jìn),黃魁英,夏楓耿,陳舒潔,胡海艷*
(廣州市微生物研究所,廣東廣州 510663)
為獲得產(chǎn)β-甘露聚糖酶發(fā)酵最佳條件,本研究以克隆篩選獲得的產(chǎn)耐高溫β-甘露聚糖酶的畢赤酵母菌M27-8菌株為研究對(duì)象,對(duì)不同氮源含量、葡萄糖含量、pH、發(fā)酵時(shí)間、接種量、溫度和轉(zhuǎn)速進(jìn)行優(yōu)化。結(jié)果表明:最佳產(chǎn)酶條件為6%玉米漿、3%葡萄糖、pH 5.5、發(fā)酵60 h、2%接種量、28℃、225 r/min,最大酶活達(dá)到452.7 U/mL,是優(yōu)化前酶活138 U/mL的3.3倍。綜上,此發(fā)酵培養(yǎng)基可用較廉價(jià)的玉米漿取代YPD培養(yǎng)基,從而節(jié)省成本,為工業(yè)生產(chǎn)奠定基礎(chǔ)。
β-甘露聚糖酶;畢赤酵母;發(fā)酵條件;優(yōu)化
甘露聚糖作為半纖維素的第二大組分(Scheller等,2010;St。albrand等,1993),廣泛存在于木材和各種植物組織中(Kansoh等,2004)。β-甘露聚糖酶能夠水解含有甘露聚糖的有機(jī)物,已經(jīng)廣泛應(yīng)用于飼料、造紙、醫(yī)藥、食品、精細(xì)化工等領(lǐng)域(Kansoh 等,2004;Bhat等,2000)。 甘露聚糖酶因在飼料應(yīng)用中對(duì)酶的純度要求不高,無需精制純化,通過簡(jiǎn)單的發(fā)酵生產(chǎn)并噴霧干燥獲得酶粉,一般就可作為飼料添加物混合使用(Chen等,2005)。在常用飼料原料中半纖維素含量豐富,因其在單胃動(dòng)物消化道內(nèi)會(huì)形成凝膠狀,從而影響畜禽吸收和飼料利用率(Nunes等,1992)。β-甘露聚糖酶可添至飼料中,消除其中甘露聚糖的抗?fàn)I養(yǎng)作用,提高飼料利用率,并且可促進(jìn)腸道內(nèi)有益菌群的增殖,增強(qiáng)畜禽免疫力(Cerqueira等2011;Zou 等,2006)。
目前許多工業(yè)用酶,要求具有高活性酶的同時(shí),還需要耐熱和耐酸性條件,因此,獲得耐熱性、耐酸性的β-甘露聚糖酶菌株的研究成為熱點(diǎn)。目前,表達(dá)耐高溫、耐酸性β-甘露聚糖酶的菌株應(yīng)用到工業(yè)化生產(chǎn)中的不多。本研究對(duì)構(gòu)建組成型β-甘露聚糖酶畢赤酵母高效表達(dá)體系,篩選的高酶活、耐熱性和耐酸性較強(qiáng)的β-甘露聚糖酶菌株M27-8為研究對(duì)象,對(duì)培養(yǎng)基成分及培養(yǎng)條件進(jìn)行優(yōu)化,提高該菌株的產(chǎn)酶水平,為進(jìn)一步擴(kuò)大生產(chǎn)奠定良好基礎(chǔ)。
1 材料與方法
1.1 材料與儀器 蛋白胨(貴州新華生化科技發(fā)展有限公司)、玉米漿干粉(山東康源生物科技有限公司)、酵母提取物(湖北安琪酵母股份有限公司);葡萄糖、磷酸氫鈉、磷酸氫二鉀、瓊脂等試劑為國(guó)產(chǎn)分析純。畢赤酵母菌M27-8(Pichia pastoris M27-8),由廣東省種質(zhì)資源庫(kù)提供。
DHZ-DA大容量全溫振蕩器(江蘇省太倉(cāng)市實(shí)驗(yàn)設(shè)備廠);752N紫外可見分光光度計(jì)(上海精科儀器有限公司);HR/T20M臺(tái)式高速冷凍離心機(jī)(湖南赫西儀器裝備有限公司);XW-80旋渦混合儀 (海門市其林貝爾儀器制造有限公司);PHS-3C pH計(jì)(上海儀電科學(xué)儀器股份有限公司)。
1.2 試驗(yàn)方法
1.2.1 試劑 PTM1微量元素:CuSO4·5H2O 6.0 g/L、NaI 0.08 g/L、MnSO4·H2O 3.0 g/L、Na2MoO4·2H2O 0.2 g/L、H3BO30.02 g/L、CoCl20.5 g/L、ZnCl220.0 g/L、FeSO4·7H2O 65.0 g/L、 生物素 0.2 g/L、 濃 H2SO45.0 ml/L,過濾除菌。發(fā)酵培養(yǎng)基發(fā)酵前按照4.0 mL/L比例補(bǔ)充PTM1。
0.5%甘露聚糖溶液(pH5.0):用pH 5.0磷酸二氫鉀—磷酸氫二鈉緩沖液配制0.5%甘露聚糖底物液。DNS溶液的配置參照農(nóng)業(yè)部標(biāo)準(zhǔn)DNS試劑的配制。
YPD培養(yǎng)基:酵母提取物10 g/L、蛋白胨20 g/L、葡萄糖10 g/L。
YPDS培養(yǎng)基:酵母提取物10g/L、蛋白胨20g/L、葡萄糖10 g/L、瓊脂20 g/L。
BSM培養(yǎng)基:葡萄糖 20 g/L、85%H3PO426.7 mL/L、KOH 4.13 g/L、(NH4)2SO44 g/L、CaCl20.38 g/L、K2SO418.2 g/L、MgSO4·2H2O 14.9 g/L、CaSO4·2H2O 0.93 g/L、PTM1 4.0 mL/L。
1.2.2 種子液的制備 實(shí)驗(yàn)室保存的畢赤酵母菌M27-8,轉(zhuǎn)接入含 200 μg/mL Zeocin的 YPDS平板培養(yǎng)基中,28℃培養(yǎng)3 d,涂板。將平板培養(yǎng)基活化好的菌種,挑選單菌落接種到50 mL YPD液態(tài)培養(yǎng)基中,28℃、200 r/min振蕩培養(yǎng)20~24 h,OD600達(dá)到6.0~8.0,鏡檢,視野中酵母菌飽滿、均勻、單個(gè)或兩個(gè)成串的培養(yǎng)液作為發(fā)酵種子液。
1.2.3 培養(yǎng)基成分優(yōu)化
1.2.3.1 不同氮源對(duì)產(chǎn)β-甘露聚糖酶的影響。分別采用2%、4%、6% 濃度的BSM、棉籽粉、中溫豆粉、玉米漿作為氮源,添加20 g/L的葡萄糖為碳源,以YPD培養(yǎng)基為對(duì)照。接種量2%、裝液量50 mL/250 mL,轉(zhuǎn)速200 r/min,用八層無菌紗布封口培養(yǎng),28℃培養(yǎng),72 h取樣,測(cè)定β-甘露聚糖酶酶活。
1.2.3.2 不同玉米漿干粉含量對(duì)產(chǎn)β-甘露聚糖酶的影響。按照上述最佳條件,采用3%、4%、5%、6%、7%、8%、10%不同玉米漿含量的培養(yǎng)基進(jìn)行發(fā)酵試驗(yàn),5 moL/L氫氧化鈉調(diào)節(jié)pH 5.0,其他條件同上。
1.2.3.3不同葡萄糖含量對(duì)產(chǎn)β-甘露聚糖酶的影響。按照上述最佳條件,采用1%、2%、3%、4%、5%不同葡萄糖含量的培養(yǎng)基進(jìn)行發(fā)酵試驗(yàn),5 moL/L氫氧化鈉調(diào)節(jié)pH 5.0,其他條件同上。
1.2.4 培養(yǎng)條件優(yōu)化
1.2.4.1 不同發(fā)酵液pH對(duì)產(chǎn)β-甘露聚糖酶的影響。按照上述最佳條件,用5 moL/L的氫氧化鈉和10%磷酸調(diào)節(jié)初始pH,調(diào)節(jié)pH為4.0、4.5、5.0、5.5、6.0,以不調(diào)節(jié) pH 為對(duì)照,每 12 h調(diào)節(jié)發(fā)酵液pH,其他條件同上。
1.2.4.2 不同接種量對(duì)產(chǎn)β-甘露聚糖酶的影響。按照上述最佳條件,采用1%、2%、5%、7.5%、10%接種量進(jìn)行發(fā)酵試驗(yàn),其他條件同上。
1.2.4.3 不同溫度對(duì)產(chǎn)β-甘露聚糖酶的影響。按照上述最佳條件,設(shè)置兩組分別在28℃和30℃下培養(yǎng)24 h,最后每組分別在24、28、32℃繼續(xù)培養(yǎng)至72 h,其他條件同上。
1.2.4.4 不同轉(zhuǎn)速對(duì)產(chǎn)β-甘露聚糖酶的影響。按照上述最佳條件,采用 150、175、200、225、250 r/min,其他條件同上。
1.2.5 胞外粗酶液的制備 取發(fā)酵物2~4 mL至離心管中,9000 r/min離心5 min,上清液為粗酶液。
1.2.6 β-甘露聚糖酶酶活測(cè)定 取直徑15 mm的15 mL的潔凈刻度試管分別標(biāo)記空白組、對(duì)照組、測(cè)試組,分別加入1.5 mL pH 5.0的0.5%甘露聚糖溶液作為底物,45℃水浴預(yù)熱5 min;測(cè)試組加入稀釋適當(dāng)倍數(shù)的預(yù)熱酶液0.5 mL,在45℃水浴10 min,每隔2 min輕輕搖晃均勻;10 min后立即加入2 mL DNS溶液終止反應(yīng);然后僅在對(duì)照組中加入0.5 mL酶液,立即至于沸水中煮沸15 min,顯色反應(yīng)后,冷卻至室溫后用蒸餾水定容至15 mL,顛倒混勻;以空白管調(diào)零,在分光光度計(jì)540 nm波長(zhǎng)處測(cè)吸光度。
酶活力單位(U):在45℃、pH 5.0的條件下,每分鐘水解底物產(chǎn)生1 μmol甘露糖所需的酶量為一個(gè)酶活單位。
1.2.7 甘露糖標(biāo)準(zhǔn)曲線 用pH 5.0磷酸二氫鉀—磷酸氫二鈉緩沖液配制 0、0.4、0.8、1.2、1.6、2.0 mg/mL不同濃度梯度,以甘露糖濃度為橫坐標(biāo),吸光度為縱坐標(biāo)線性擬合得到線性回歸方程:Y=0.15502X-0.039,R2=0.99845。
2 結(jié)果與分析
2.1 不同氮源對(duì)產(chǎn)β-甘露聚糖酶的影響 在培養(yǎng)基中分別添加玉米漿干粉、棉籽餅粉、中溫豆餅粉、BSM、YPD發(fā)酵72 h測(cè)定酶活。結(jié)果見圖1。
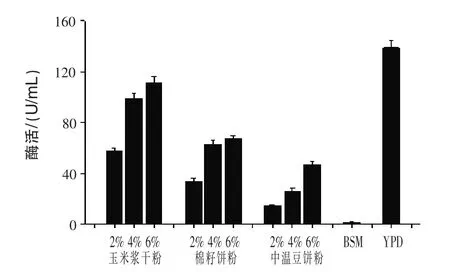
圖1 不同氮源對(duì)產(chǎn)β-甘露聚糖酶的影響
由圖1可知,發(fā)酵最佳產(chǎn)β-甘露聚糖酶培養(yǎng)基是添加氮源YPD,酶活達(dá)到138.9 U/mL;其次是玉米漿干粉,6%含量的玉米漿干粉培養(yǎng)基發(fā)酵酶活達(dá)到111 U/mL,為YPD培養(yǎng)基發(fā)酵產(chǎn)酶的80%。棉籽餅粉和中溫豆餅粉濃度也影響菌株產(chǎn)酶,當(dāng)培養(yǎng)基中添加BSM時(shí),幾乎檢測(cè)不到酶活;試驗(yàn)結(jié)果和Jia等(2012)報(bào)道的采用玉米漿干粉作為替代氮源時(shí)最佳結(jié)果相符。因玉米漿干粉相對(duì)蛋白胨和酵母膏作為氮源時(shí)廉價(jià),故在上大型發(fā)酵罐時(shí)可以將其作為YPD培養(yǎng)基的取代物,節(jié)約生產(chǎn)成本。
2.2 不同玉米漿干粉含量對(duì)產(chǎn)β-甘露聚糖酶的影響 通過發(fā)酵72 h,測(cè)定發(fā)酵液中的酶活,結(jié)果見圖2。隨著玉米漿干粉含量增加,β-甘露聚糖酶產(chǎn)量先增后減;且玉米漿含量在6%~8%時(shí),產(chǎn)酶相對(duì)平穩(wěn)。這可能是玉米漿成分復(fù)雜,顯酸性,含量過高時(shí)會(huì)抑制微生物產(chǎn)酶。最佳的玉米漿干粉含量在6%左右。
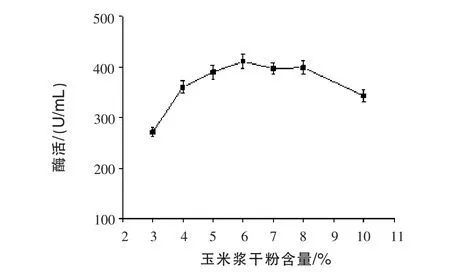
圖2 不同玉米漿干粉含量對(duì)產(chǎn)β-甘露聚糖酶的影響
2.3 不同葡萄糖含量對(duì)產(chǎn)β-甘露聚糖酶的影響通過發(fā)酵72 h,測(cè)定發(fā)酵液中的酶活,結(jié)果見圖3。隨著葡萄糖含量增加,β-甘露聚糖酶酶活逐漸升高,且在3%后不再增加。這可能是葡萄糖含量過高,會(huì)產(chǎn)生葡萄糖抑制效應(yīng),從而抑制微生物的生長(zhǎng)(Olaniyi等,2015)。最佳的葡萄糖含量為3%。
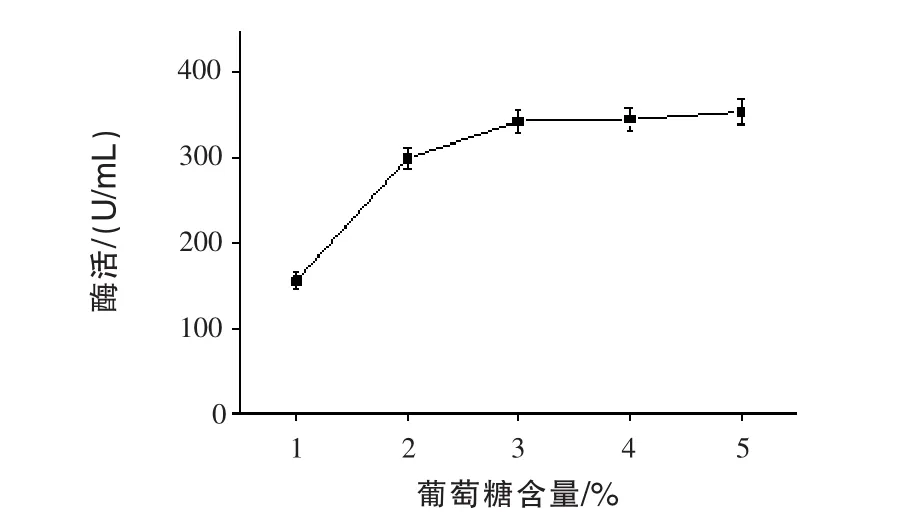
圖3 不同葡萄糖含量對(duì)產(chǎn)β-甘露聚糖酶的影響
2.4 不同發(fā)酵液pH對(duì)產(chǎn)β-甘露聚糖酶的影響通過調(diào)節(jié)0~72 h發(fā)酵液中的pH,每12 h調(diào)pH一次,以不調(diào)pH作為對(duì)照,測(cè)定發(fā)酵液中β-甘露聚糖酶酶活,結(jié)果見圖4。隨著發(fā)酵時(shí)間的延長(zhǎng),β-甘露聚糖酶酶活基本保持增長(zhǎng)的趨勢(shì)。通過發(fā)酵過程中控制不同發(fā)酵pH,只有發(fā)酵pH 4.0較自然發(fā)酵效果差,其余效果都較對(duì)照好,其中pH為5.0、5.5、6.0效果最優(yōu),且三者之間酶活差別不大,說明發(fā)酵液pH值為5.0~6.0為產(chǎn)酶的適宜濃度,是不調(diào)pH產(chǎn)酶的2.5倍左右。
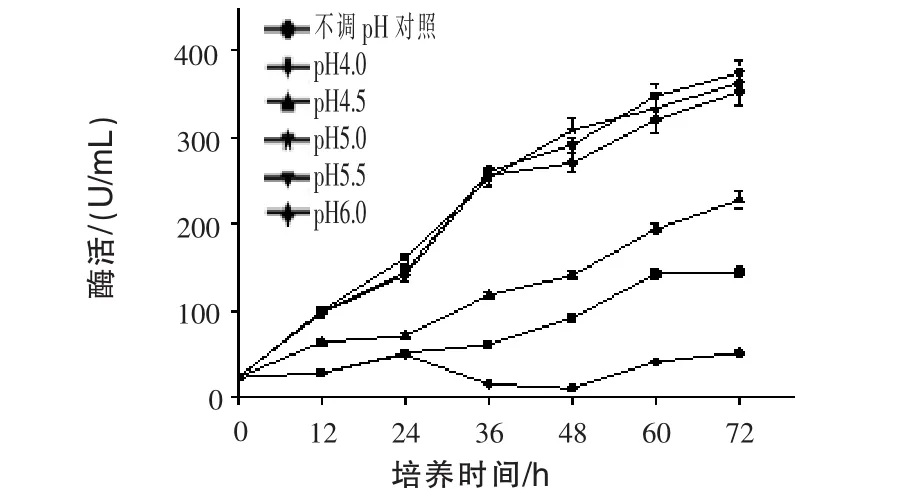
圖4 不同發(fā)酵液pH對(duì)產(chǎn)β-甘露聚糖酶的影響
2.5 不同接種量對(duì)產(chǎn)β-甘露聚糖酶的影響 通過發(fā)酵96 h,測(cè)定發(fā)酵液中的酶活,結(jié)果見圖5。隨著發(fā)酵時(shí)間的增加,產(chǎn)酶基本保持較快增長(zhǎng)的趨勢(shì);接種量不同,產(chǎn)酶差異不明顯。在發(fā)酵過程中,只有接種量為1%時(shí),前60 h和其他接種量有較大差異,說明接種量最少要控制在2%,才不會(huì)因?yàn)榫w濃度而影響產(chǎn)酶。
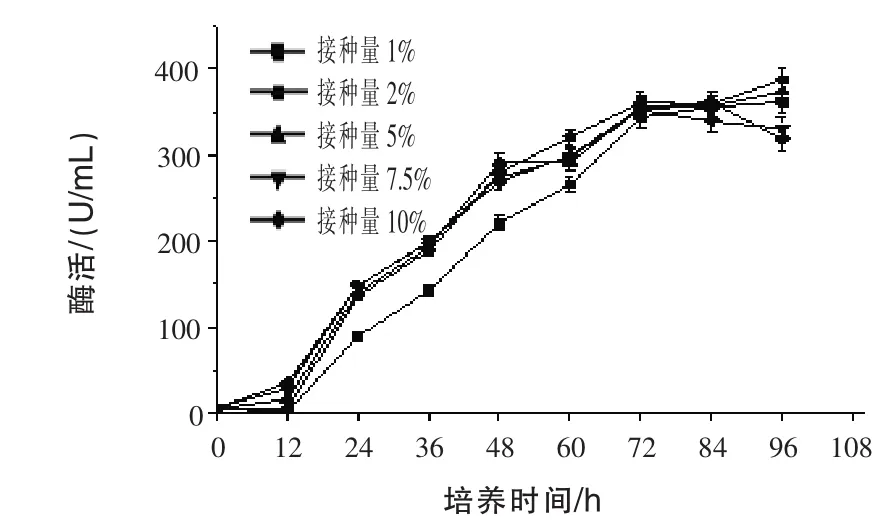
圖5 不同接種量對(duì)產(chǎn)β-甘露聚糖酶的影響
2.6 不同溫度對(duì)產(chǎn)β-甘露聚糖酶的影響 通過發(fā)酵72 h,測(cè)定發(fā)酵液中的酶活,結(jié)果見圖6。前期兩組分別在28℃和30℃培養(yǎng)24 h測(cè)定酶活,即圖中28℃培養(yǎng)起始值和30℃培養(yǎng)起始值,30℃培養(yǎng)起始值曲線較28℃培養(yǎng)起始值曲線高,這可能是30℃培養(yǎng)更適合微生物前期生長(zhǎng),較利于產(chǎn)酶,故前期采用30℃培養(yǎng)較適宜。兩組在28℃和30℃培養(yǎng)24 h后,后期分別至于24、28℃和30℃培養(yǎng)72 h測(cè)定酶活,發(fā)現(xiàn)后期培養(yǎng)溫度越高越不利于產(chǎn)酶,且前期24 h在30℃培養(yǎng)后轉(zhuǎn)入32℃培養(yǎng)至72 h時(shí),酶活相較于其他條件下降得更快,可能是由于后期發(fā)酵溫度過高,不利于微生物生長(zhǎng),導(dǎo)致蛋白質(zhì)降解,對(duì)外源蛋白質(zhì)的表達(dá)不利(Ye等,2008)。故后期采用不超過28℃培養(yǎng)較適宜。
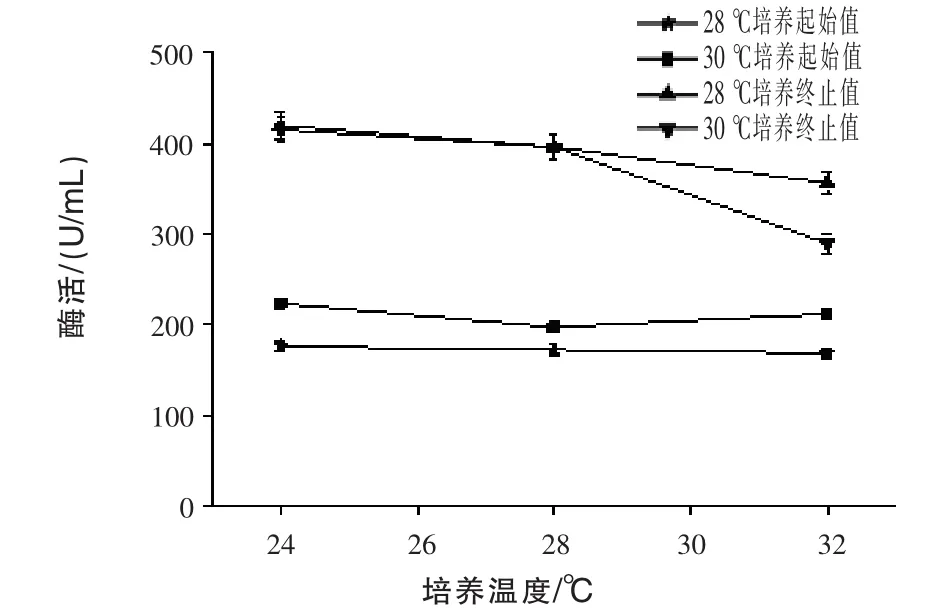
圖6 不同溫度對(duì)產(chǎn)β-甘露聚糖酶的影響
2.7 不同轉(zhuǎn)速對(duì)產(chǎn)β-甘露聚糖酶的影響 通過發(fā)酵72 h,測(cè)定發(fā)酵液中的酶活,結(jié)果見圖7。隨著時(shí)間的推移,酶活逐漸增加,且在發(fā)酵60 h之后,酶活保持穩(wěn)定;隨著轉(zhuǎn)速增加,產(chǎn)酶前期48 h內(nèi)保持增加趨勢(shì),后期逐漸平穩(wěn)。發(fā)酵過程中轉(zhuǎn)速越大,有利于促進(jìn)溶氧,促進(jìn)物質(zhì)間的傳遞轉(zhuǎn)化,有利于產(chǎn)酶。225 r/min和250 r/min在60 h后差異較小,且發(fā)酵周期縮短能夠減少生產(chǎn)成本,如果采用225 r/min,發(fā)酵60 h和72 h產(chǎn)酶并無明顯差異。故采用60 h發(fā)酵,轉(zhuǎn)速225 r/min較適宜。

圖7 不同轉(zhuǎn)速對(duì)產(chǎn)β-甘露聚糖酶的影響
3 結(jié)論
本文采用畢赤酵母組成型工程菌株M27-8進(jìn)行高密度發(fā)酵,優(yōu)化培養(yǎng)基配比及培養(yǎng)條件,得出最佳產(chǎn)酶條件。優(yōu)化后基本培養(yǎng)基組成為6%的玉米漿干粉作為氮源,3%的葡萄糖作為碳源,較采用優(yōu)化前的YPD培養(yǎng)基價(jià)格低廉,適宜規(guī)模化生產(chǎn);優(yōu)化后基本培養(yǎng)參數(shù)采用發(fā)酵pH為5.5,培養(yǎng)時(shí)間60 h,接種量2%,發(fā)酵溫度不超過28℃,轉(zhuǎn)速225 r/min,能夠達(dá)到最佳的產(chǎn)酶條件,最大酶活達(dá)到452.7 U/mL,是優(yōu)化前酶活138 U/mL的3.3倍。
[1]Bhat MK.Cellulases and related enzymes in biotechnology[J].Biotechnology Advances,2000,18(5):355 ~ 383.
[2]Chen HL,F(xiàn)an YH,Chen ME,et al.Unhydrolyzed and hydrolyzed konjac glucomannans modulated cecal and fecal microflora in Balb/c mice[J].Nutrition,2005,21(10):1059 ~ 1064.
[3]Cerqueira M A,Bourbon A I,Pinheiro A C,et al.Galactomannans use in the development of edible films/coatings for food applications[J].Trends in Food Science&Technology,2011,22(1):662 ~ 671.
[4]Jia Zheng,Wei Zhao,Ning Guo,et al.Development of an industrial medium and a novel fed-batch strategy for high-level expression of recombinant b-mananase by Pichia pastoris[J].Bioresource Technology,2012,118(1):257 ~ 264.
[5]Ghose TK.Measurement of cellulose activities[J].Pure and Applied Chemistry,1987,59(2):257 ~ 268.
[6]Kansoh AL,Nagieb ZA.Xylanase and Mannanase enzymes from Streptomyces galbus NR and their use in biobleaching of softwood kraft pulp[J].Antonie Van Leeuwonhoek,2004,85(2):103 ~ 114.
[7]Nunes C S,Malmlo..f K.Effects of guar gum and cellulose on glucose absorption,hormonal release and hepatic metabolism in the pig[J].British Journal of Nutrition,1992,68(3):693 ~ 700.
[8]Olaniyi O O,Ekundayo T C,Igbe O F,et al.Influence of Cultural and Nutritional Factors on β-Mannanase Production by Penicillium italicum under Submerged State Fermentation[J].British Microbiology Research Journal,2015,5(6):481 ~ 489.
[9]Scheller H V,Ulvskov P.Hemicelluloses[J].Annual Review of Plant Biology ,2010,61(1):263 ~ 289.
[10]St。albrand H,Siikaaho M,Tenkanen M,et al.Purification and characterization of two β -Mannanase from Trichoderma reesei[J].Biotechnology,1993,29(3):229 ~ 242.
[11]Wu D,Hao Y,Chu J,et al.Inhibition of degradation and aggregation of recombinant human comsensus interferon-a mutant expressed in Pichia pastoris with complex medium in bioreactor [J].Appl Microbiol Biotechnol,2008,80(1):1063 ~ 1071.
[12]Ye D O U,Qing-lu W,Qiao-qiao L I.Optimization of the fermentation conditions of Pichia Pastoris[J].Science and Technology of Food Industry,2008,29(6):168 ~ 171.
[13]Zou X T,Xiao X J,Xu Z R.Effect of β-Mannanase (Hemicell) on growth performance and immunity of broilers[J].Poultry Science,2006,85(12):2176 ~ 2179.■
The optimal conditions for the production of β-manganese were obtained by using Pichia pastors engineering strain M27-8.The content nitrogen sources,glucose content,pH,fermentation time,inoculum size,temperature and speed were optimized.The results showed that the optimum conditions for enzyme production were as follows:6%corn steep liquor,3%glucose,pH 5.5,fermentation 60 h,2%inoculation amount,28 ℃,225 r/min,the maximum activity reached 452.7 U/mL,which was 3.3 times of that of the optimized enzyme activity of 138 U/mL.In conclusion,substitution of YPD medium with cheaper corn pulp medium could save costs and lay the foundations for industrial production.
β-mannanase;Pichia pastoris;fermentation conditions;optimization
S816.3
A
1004-3314(2017)19-0013-04
10.15906/j.cnki.cn11-2975/s.20171903
廣州市科技計(jì)劃項(xiàng)目(201710010154)
*通訊作者
