新疆野生甜瓜形態學觀察及其與不同變種雜交親和性研究
2017-10-16 02:13:14李寐華樊國全王廣智張學軍伊鴻平張永兵
新疆農業科學 2017年9期
關鍵詞:新疆
李寐華,樊國全,楊 永,王廣智,張 建,張學軍,伊鴻平,張永兵
(1.新疆農業科學院哈密瓜研究中心,烏魯木齊 830091;2.新疆農業大學林學與園藝學院,烏魯木齊 830052;3.新疆農業科學院科研管理處,烏魯木齊 830091;4.浙江衢州新農都實業有限公司,浙江衢州 324022)
新疆野生甜瓜形態學觀察及其與不同變種雜交親和性研究
李寐華1,2,樊國全3,楊 永1,王廣智1,張 建4,張學軍1,伊鴻平1,張永兵1
(1.新疆農業科學院哈密瓜研究中心,烏魯木齊 830091;2.新疆農業大學林學與園藝學院,烏魯木齊 830052;3.新疆農業科學院科研管理處,烏魯木齊 830091;4.浙江衢州新農都實業有限公司,浙江衢州 324022)
目的通過形態學觀察鑒定與種內不同變種間雜交親和性研究,為明確新疆野生甜瓜分類地位和擴大甜瓜栽培品種遺傳基礎提供支撐。方法觀察比較兩份新疆野生甜瓜和地方品種葉片、莖、花及果實等28個形態學性狀,利用苗期接種技術鑒定新疆野生甜瓜對蔓枯病和白粉病的抗病性。同時,以亞種ssp.agrestis內var.agrestis、var.conomon、var.dudaim、var.chito和var.momordica等5個變種,亞種ssp.melo內var.cantalupensis、var.inodorus和var.flexuosus等3個變種為試材,通過人工授粉雜交后坐果率和結籽率的觀察記錄,分析新疆野生甜瓜與種內的8個變種間的雜交親和性。結果哈密野甜瓜絕大多數性狀與栽培品種對照差異明顯,而吐魯番野甜瓜大部分性狀與對照接近。苗期人工接種甜瓜蔓枯病和白粉病菌后,哈密野甜瓜高抗蔓枯病、不抗白粉病,吐魯番野甜瓜和對照易感或高感蔓枯病和白粉病。人工授粉雜交后,新疆野生甜瓜與2個亞種的8個變種之間未發現不親和不稔現象;野生甜瓜與8個變種雜交果實的結籽率正反交間有差異,以野生甜瓜為母本時的結籽率高于父本時的;此外,哈密野甜瓜與亞種ssp.agrestis和ssp.melo雜交的平均結籽率分別為79.3%和83.4%,吐魯番野甜瓜與兩個亞種雜交的平均結籽率分別為76.1%和79.4%,野生甜瓜與兩個亞種雜交的平均結籽率相近。結論新疆野生甜瓜哈密野甜瓜的形態性狀與亞種ssp.agrestis內野生變種的性狀相近,而吐魯番野甜瓜的形態性狀接近于地方栽培品種。經苗期接種鑒定,哈密野甜瓜對甜瓜蔓枯病表現高抗。新疆野生甜瓜與甜瓜兩個亞種ssp.agrestis和ssp.melo可正常雜交且雜種后代可育,應當被劃分在C.melo.內。
野生甜瓜;形態學;結籽率;雜交親和性
0 引 言
【研究意義】甜瓜(CucumismeloL.)種內野生資源豐富,主要分布在非洲、亞洲、大洋洲和美洲等地的一些國家[1-3]。野生甜瓜在進化過程中積累了優異的基因資源,例如對生物和非生物脅迫的抗逆性和耐受性等性狀,而甜瓜栽培品種經長期的人為選擇后卻逐漸遺失了這些特異性狀[3,4]。野生甜瓜形態學和農藝特征的鑒定,以及和種內不同亞種的變種間雜交親和性研究,不僅能明確野生甜瓜資源在種內的分類地位,還可擴大甜瓜栽培品種的遺傳基礎,對于甜瓜的分類研究和育種實踐均具有重要的意義。【前人研究進展】甜瓜是葫蘆科(Cucurbitaceae)甜瓜屬(Cucumis)內遺傳變異最大的一個種,甜瓜的植株、葉片和果實的形態變異類型非常豐富,尤其是果實的形態性狀如果實形狀、果皮顏色、果肉顏色、果肉風味等性狀遺傳差異明顯[5-8]。根據果實的形態性狀特點,甜瓜種被分為兩個亞種C.melossp.agrestis(Naud.) Pangalo和C.melossp.melo,亞種ssp.agrestis包括所有的野生甜瓜變種,而亞種ssp.melo包括主要的栽培甜瓜變種[9-12]。由于難以克服的雜交障礙,甜瓜與屬內其他種(C.spp.)之間的遠緣雜交仍未取得成功[13-16]。利用種內的野生甜瓜與栽培變種雜交后代群體,研究人員分析鑒定出甜瓜白粉病和病毒病、蚜蟲和白粉虱等病蟲害的抗性基因和數量性狀基因座(QTL,quantitative trait loci)[17-19],并獲得多個控制甜瓜果實品質和農藝經濟性狀的QTL[20]。A.Roy等[2]對印度北部的野生甜瓜資源進行系統的形態學和農藝學觀察后,發現了具有抗根結線蟲和高度耐旱特異性狀的野生甜瓜,為栽培甜瓜抗根結線蟲和耐旱性育種研究奠定基礎。我國甜瓜種質資源豐富、分布廣泛,被認為是甜瓜的次級多樣化中心[21]。在我國黃河流域和西北地區的新疆均發現了野生甜瓜,初步的觀察表明這些野生甜瓜的結實力強、果實小、商品性差,還發現野生甜瓜與當地栽培品種間可正常授粉雜交并坐果[22]。利用形態性狀數據進行聚類分析后,發現新疆野生甜瓜與栽培地方品種的親緣關系很遠,可能具有完全不同的遺傳背景[23]。【本研究切入點】準確的生物學特性和系統分類的鑒定,有助于種質資源在甜瓜育種中的應用研究[24]。20世紀在新疆收集到兩份野生甜瓜哈密野甜瓜和吐魯番野甜瓜,并觀察記載了這兩份野生甜瓜的部分形態性狀[25,26]。對新疆野生甜瓜缺乏系統的形態學和農藝學特征的鑒定,對新疆野生甜瓜與種內兩個亞種的不同變種間的雜交親和性也缺乏相應的研究,這些都不利于野生甜瓜種質的保存和利用。研究新疆野生甜瓜形態學觀察及其與不同變種雜交親和性。【擬解決的關鍵問題】觀察新疆野生甜瓜形態學和農藝學性狀,鑒定新疆野生甜瓜對蔓枯病和白粉病的抗病性,通過人工授粉雜交分析新疆野生甜瓜與種內兩個亞種的不同變種間的雜交親和性,明確新疆野生甜瓜的系統分類地位補充理論知識,為擴大我國甜瓜栽培品種的遺傳背景提供支撐。
1 材料與方法
1.1 材 料
形態學和農藝學對照材料皇后,是20世紀80年代由新疆農業科學院利用雜交育種方法選育的常規品種。野生甜瓜哈密野甜瓜在新疆哈密地區哈密市南湖鄉收集,吐魯番野甜瓜在吐魯番地區鄯善縣辟展鄉東湖村收集。屬于亞種ssp.agrestis的變種包括var.agrestis、var.conomon、var.dudaim、var.chito和var.momordica,變種agrestis為引自印度的PI 164797,變種conomon為引自韓國和日本的PI 161375和“婆殺”,變種dudaim為引自印度的PI 124449,變種chito為引自津巴布韋的PI 482421,變種momordica為引自土耳其的PI 164975;屬于亞種ssp.melo的變種包括var.cantalupensis、var.inodorus和var.flexuosus,變種cantalupensis為引自阿富汗和伊朗的PI 126165和PI 140814,變種inodorus為引自西班牙和土耳其的PI 401650和PI 164976,變種flexuosus為引自伊拉克的PI 435288。所有供試甜瓜的種子均由新疆農業科學院哈密瓜研究中心提供。
1.2 方 法
1.2.1 形態學性狀觀察
新疆野生甜瓜和對照材料按照隨機區組種植,單行區,每行定植10株,行距3.5 m,株距0.4 m,2次重復。每個小區隨機取5株調查和測量田間性狀,果實成熟時每個地方品種至少采收3個果實進行調查和測量。測量16個質量性狀和12個數量性狀,葉片、莖、花及果實等的長度和直徑用直尺測量,重量用電子秤(型號ACS-30)測量,可溶性固形物含量用手持測糖儀(型號DR-103)測量。
1.2.2 病害苗期接種和調查
新疆野生甜瓜和對照的種子播種前經65℃的高溫處理48 h,播種基質為經過高溫處理的完全腐熟的椰糠。蔓枯病(Gummy Stem Blight, GSB)和白粉病(Powdery Mildew, PM)兩種病害的每份試驗材料分別種植10株,重復3次,完全隨機設計試驗。植株2~3片真葉完全展開時接種病害。參照Zhang等[27]的方法苗期接種蔓枯病原菌,調查植株莖部癥狀并對蔓枯病害進行分級。參照咸豐等[19]的方法苗期接種白粉病原菌,調查每株基部3片葉癥狀并對白粉病害進行分級。接種后相對濕度保持在80%以上,2周后調查統計病情。
1.2.3 人工授粉雜交
新疆野生甜瓜和8份變種育苗后,按照株行距0.4 m × 3.5 m定植,所有試驗材料按完全隨機排列定植。定植約3周后開始整枝,摘除供試材料第8節位之前的全部側蔓,從第9節開始留取雌花用于人工授粉。授粉開始時,第1 d下午將次日開放的雌花人工去雄后套袋,并收集每份材料次日開放的雄花恒溫保濕保存,第2 d清晨人工授粉雜交后套袋隔離。2份新疆野生甜瓜分別與8份變種間正反雜交,每個雜交組合授粉5個植株,每株授粉2朵雌花、共10朵雌花,每個雜交組合3次重復。每份材料取3株用于自然授粉,用作統計結籽率的對照。
1.3 數據處理
參照周曉慧等[28]的方法劃分試材對蔓枯病的抗感表型,參照咸豐等[19]的方法劃分對白粉病的抗感表型。蔓枯病和白粉病的病情指數按以下公式計算:
病情指數=Σ(級值×株數)/(最高級值×總株數)×100%.
授粉2周后調查記錄坐果情況。授粉4周后采收果實調查統計果實的結籽率,以各試材自然授粉的果實結籽率為對照。
坐果率=坐果數/授粉數 ×100%.
結籽率=飽滿種子數/(飽滿種子數 + 秕種子數)×100%.
利用軟件SPSS.10.1對果實的結籽率進行方差分析。
2 結果與分析
2.1 新疆野生甜瓜形態特征
研究表明,相對于地方栽培品種,野生甜瓜哈密野甜瓜的植株長勢較旺,基部分枝力較強、分枝數量多,坐果能力極強,單株平均坐果數量超過40個。哈密野甜瓜的葉片有淺葉裂,而對照地方栽培品種“皇后”的葉片全緣,哈密野甜瓜子房上的絨毛少,而對照的子房絨毛濃密。哈密野甜瓜的果形、果皮顏色、果肉顏色和果肉風味等質量性狀與對照的差異非常明顯,其中果肉風味呈酸味表現尤為獨特,而其他質量性狀如雄花著生、花瓣顏色、性別表達方式、種皮顏色、脫蒂和裂果性與對照的相似。哈密野甜瓜的大部分數量性狀與對照的差異顯著,除了主蔓節長略大于對照外,其他數量性狀如果實縱徑、單果重量和種子百粒重等的數值與對照相差數倍甚至幾十倍。
吐魯番野甜瓜植株的長勢適中,基部分枝數量和坐果能力與對照的差異不明顯。吐魯番野甜瓜子房上的絨毛適中,而對照的子房絨毛濃密。吐魯番野甜瓜的果形呈橄欖形、果皮顏色為綠色,而對照的果實成熟時呈橢圓或棒狀、果皮顏色為淡黃或黃色。吐魯番野甜瓜果肉顏色、果肉風味和種皮顏色等質量性狀與對照的差異較明顯,而其他質量性狀和對照的基本一致。吐魯番野甜瓜的大部分數量性狀值略低于對照的,除單果重量、可溶性固形物、種子縱徑和種子百粒重的數值明細低于對照外。此外,吐魯番野甜瓜數量性狀的值均介于哈密野甜瓜與對照的數值之間。
野生甜瓜和對照幼苗接種蔓枯病菌后,最初在子葉上出現癥狀,真葉上表現癥狀較晚,多數葉片的葉緣處出現淺褐色“V”形病斑。接種一周后,植株莖部開始發病,莖部病斑表面有橙色或淺黃色液體溢出,發病后期病斑逐漸延伸擴大。接種14 d后調查莖部病害,哈密野甜瓜病情指數為14%表現高抗蔓枯病,吐魯番野甜瓜和對照的病情指數高達80%均表現為高感病害。苗期人工接種甜瓜白粉病菌后,在植株基部真葉頁面上出現癥狀,初期白色的病斑菌落很小,發病后期菌落逐漸擴大并形成連片病斑,白色菌絲和病孢子的數量也逐漸增多,形成較厚的白色病斑而布滿葉片。接種7 d后調查葉部病害,哈密野甜瓜和吐魯番野甜瓜的病情指數分別為74%和78%,均表現為感病,而對照的病情指數大于80%表現為高感。表1,圖1
2.2新疆野生甜瓜與不同變種雜交坐果率和果實結籽率
研究表明,哈密野甜瓜和吐魯番野甜瓜與兩個亞種內的不同變種間正反雜交均能正常坐果,各雜交組合的坐果率均大于50%,雜種后代植株的生長發育均表現正常,未發現雜交不親和及不稔現象。以野生甜瓜為母本時,哈密野甜瓜自然授粉的結籽率與各變種人工授粉雜交的差異不顯著,吐魯番野甜瓜與變種chito人工授粉雜交的結籽率顯著小于自然授粉的,而與其余變種人工授粉雜交的結籽率和自然授粉的差異不顯著;以各變種為母本時,自然授粉的結籽率與兩份野生甜瓜人工授粉雜交的差異不顯著,表明人工授粉雜交對果實結籽率的影響可忽略不計。以哈密野甜瓜為母本、吐魯番野甜瓜為父本時,與不同變種雜交后果實的結籽率差異不顯著,反之以哈密野甜瓜為父本、吐魯番野甜瓜為母本時差異顯著。新疆野生甜瓜與不同變種雜交果實的結籽率正反交間存在差異,除變種agrestis和chito外,以野生甜瓜為父本時的結籽率均低于母本時的結籽率;此外,野生甜瓜與變種conomon、dudaim、momordica和flexuosus正反交間的結籽率差異較明顯。哈密野甜瓜與亞種ssp.agrestis和ssp.melo內變種正反雜交的平均結籽率分別為79.3%和83.4%,吐魯番野甜瓜與兩個亞種內各變種正反雜交的平均結籽率分別為76.1%和79.4%,野生甜瓜與兩個亞種雜交的平均結籽率相近。表2
表1 新疆野生甜瓜與栽培品種形態性狀比較
Table 1 Morphological comparison of Hami wild, Turpan wild and C. melo cultivars
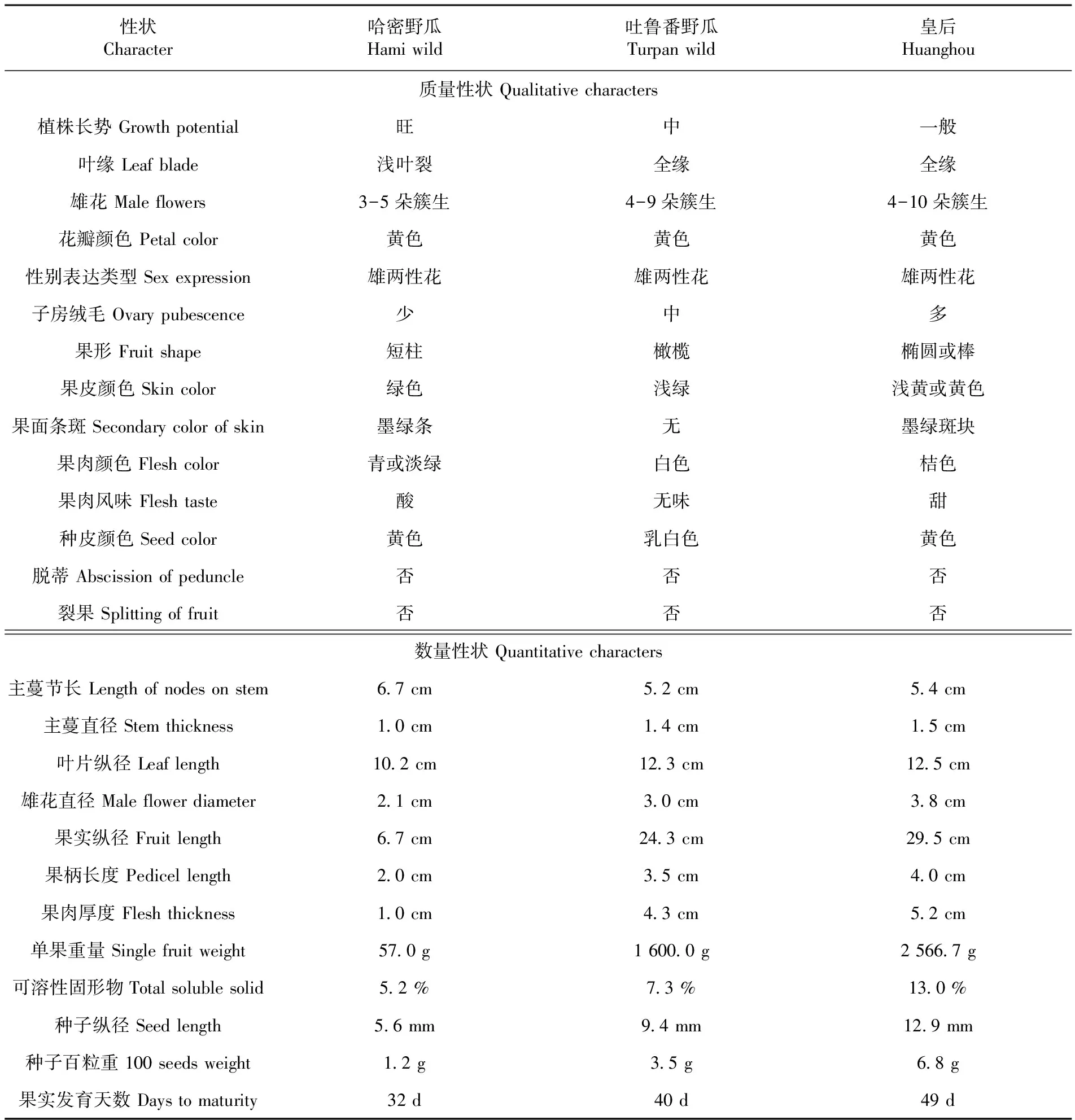
性狀Character哈密野瓜Hamiwild吐魯番野瓜Turpanwild皇后Huanghou質量性狀Qualitativecharacters植株長勢Growthpotential旺中一般葉緣Leafblade淺葉裂全緣全緣雄花Maleflowers3-5朵簇生4-9朵簇生4-10朵簇生花瓣顏色Petalcolor黃色黃色黃色性別表達類型Sexexpression雄兩性花雄兩性花雄兩性花子房絨毛Ovarypubescence少中多果形Fruitshape短柱橄欖橢圓或棒果皮顏色Skincolor綠色淺綠淺黃或黃色果面條斑Secondarycolorofskin墨綠條無墨綠斑塊果肉顏色Fleshcolor青或淡綠白色桔色果肉風味Fleshtaste酸無味甜種皮顏色Seedcolor黃色乳白色黃色脫蒂Abscissionofpeduncle否否否裂果Splittingoffruit否否否數量性狀Quantitativecharacters主蔓節長Lengthofnodesonstem6 7cm5 2cm5 4cm主蔓直徑Stemthickness1 0cm1 4cm1 5cm葉片縱徑Leaflength10 2cm12 3cm12 5cm雄花直徑Maleflowerdiameter2 1cm3 0cm3 8cm果實縱徑Fruitlength6 7cm24 3cm29 5cm果柄長度Pedicellength2 0cm3 5cm4 0cm果肉厚度Fleshthickness1 0cm4 3cm5 2cm單果重量Singlefruitweight57 0g1600 0g2566 7g可溶性固形物Totalsolublesolid5 2%7 3%13 0%種子縱徑Seedlength5 6mm9 4mm12 9mm種子百粒重100seedsweight1 2g3 5g6 8g果實發育天數Daystomaturity32d40d49d

A:哈密野甜瓜植株和坐果;B:哈密野甜瓜(左)、吐魯番野甜瓜(中)和對照(右)的雄花;C:哈密野甜瓜(左)、吐魯番野甜瓜(中)和對照(右)的雌花;D:哈密野甜瓜果實;E:吐魯番野甜瓜果實;F:對照果實,標尺:5 cm
A: Plant and fruit-setting of ‘Hami wild’; B: Male flowers of ‘Hami wild’,‘Turpan wild’ and control; C: Female flowers of ‘Hami wild’,‘Turpan wild’ and control; D: Fruit of ‘Hami wild’; E: Fruit of ‘Turpan wild’; F: Fruit of control, scale: 5 cm
圖1 新疆野生甜瓜和地方品種形態性狀
Fig.1 Morphological comparison of Xinjiang wild melon and local landrace
表2 新疆野生甜瓜與兩個亞種內變種間正反雜交果實結籽率
Table 2 Seed-setting rate of reciprocal cross between Xinjiang wild and varieties of ssp. agresitis and melo

亞種Subspecies變種Varieties哈密野瓜Hamiwild吐魯番野瓜Turpanwild母本Maternalparent(%)父本?Paternalparent(%)母本Maternalparent(%)父本Paternalparent(%)ssp.agresitisagrestis79 2(96 0?)92 9a79 1ab(96 0?)87 4conomon89 4(90 8?)81 7ab81 7a(90 8?)54 0dudaim84 4(84 0?)35 9c86 6a(84 0??)54 9chito94 8(91 5??)92 2a55 9b(91 5?)86 7momordica85 8(89 7?)56 2bc91 5a(89 7?)82 8ssp.melocantalupensis95 1(93 6?)87 2a92 6a(93 6?)72 5inodorus87 6(83 2?)79 2ab92 0a(83 2?)70 5flexuosus89 0(86 4?)62 3abc94 2a(86 4?)54 4CK90 7-87 1a-
注:不同字母表示在5%水平上差異顯著。*括號內數值表示母本自然授粉果實結籽率
Note: Treatments with different letter indicated significant difference at 5% level.*Figures in brackets indicated seed-setting rate of maternal parent after natural pollination
3 討 論
收集保存的兩份新疆野生甜瓜中,哈密野甜瓜的主要形態性狀尤其是果實的性狀與地方栽培品種的差異明顯,和野外收集時記載的性狀特征一致[26]。而吐魯番野甜瓜的大部分形態性狀與地方栽培品種的比較接近,與最初野外收集時記載的原始性狀數據相差較大,吳明珠[25]認為是收集后自交繁殖期間套袋不嚴格發生了自然雜交所造成。
最初收集到兩份新疆野生甜瓜后,未對其進行系統的抗病性鑒定,僅在田間種植并經病害調查后認為兩份新疆野生甜瓜對幾種主要病害沒有抗病性[25,26]。然而,經苗期接種鑒定發現哈密野甜瓜高抗蔓枯病、不抗白粉病,而吐魯番野甜瓜卻易感染兩種病害。哈密野甜瓜最早在哈密縣南湖公社即現在的哈密市南湖鄉發現,被當地人稱作“狗瓜”,主要分布在當地低洼地或下潮地環境中[22,26],其遺傳背景中可能具有耐干旱和耐鹽堿的抗逆基因資源。
研究中,哈密野甜瓜與新疆地方栽培品種的形態差異明顯。A.Stepansky等[9]利用表型性狀和RAPD、SSR標記對23個國家的甜瓜資源進行遺傳關系分析,認為來自我國的“Gou Gua”即哈密野甜瓜與亞種ssp.melo內主要的栽培變種cantalupensis和inodorus遺傳距離較遠,而與亞種ssp.agrestis內conomon、agrestis和dudaim等變種的親緣關系卻很近。用32個表型性狀和19個SSR標記對新疆甜瓜地方種質資源進行聚類分析后,也發現哈密野甜瓜和地方栽培品種的親緣關系最遠,初步推斷其可能屬于亞種ssp.agrestis[23,29]。哈密野甜瓜的種子縱徑為5.6 mm(表1),Y.Akashi等[30]和K.Tanaka等[31]均認為種子縱徑小于9 mm的甜瓜應該屬于亞種ssp.agrestis內的變種agrestis或conomon,這和研究結果也很接近。
甜瓜與甜瓜屬(Cucumis)內其他種(C.spp.)之間存在嚴重的雜交障礙,種間雜交至今仍未取得突破[13-16]。而甜瓜種內兩個亞種ssp.agrestis和ssp.melo之間可正常雜交坐果,雜交后代自交后被用于構建作圖群體,并進行了甜瓜病蟲害抗性基因和果實品質性狀相關的QTL分析[17,18,20]。將兩份新疆野生甜瓜分別與兩個亞種ssp.agrestis和ssp.melo內8個變種進行了人工授粉雜交后,盡管不同雜交組合的果實結籽率有一定差異,但是兩份新疆野生甜瓜與8個變種間均不存在雜交障礙,雜交后代也未觀察到不稔現象。此外,新疆野生甜瓜與兩個亞種雜交的平均結籽率相近,進一步表明新疆野生甜瓜(哈密野甜瓜和吐魯番野甜瓜)應被劃分在C.melo.內。A.Stepansky等[9]認為甜瓜兩個亞種內變種間的分類界限不清晰,主要原因之一就是甜瓜種內變種間可正常雜交以及雜交后代可育。新疆野生甜瓜與各變種雜交的結籽率正反交間存在差異,除個別變種外以野生甜瓜為母本時結籽率較高,A.K.Singh和K.S.Yadava[13]研究甜瓜屬內不同種間雜交親和性時也觀察到相同的現象,屬內一些種(C.spp.)為母本時得到可育種子數量明顯高于為父本時,認為這種差異可能與生理、遺傳或者環境因素有關。新疆野生甜瓜與地方栽培品種群體間進化關系的研究,以及對新疆野生甜瓜抗逆特異基因資源的進一步挖掘,對于我國甜瓜遺傳進化和品種改良都具有重要的意義。
4 結 論
新疆野生甜瓜哈密野甜瓜的部分形態性狀與亞種ssp.agrestis內野生變種的性狀相近,而吐魯番野甜瓜的形態性狀接近于地方栽培品種,可能是吐魯番野甜瓜在保存過程中人工授粉套袋不嚴格發生了自然雜交的緣故。經苗期接種鑒定,哈密野甜瓜對甜瓜蔓枯病表現高抗、不抗白粉病,而吐魯番野甜瓜對兩種病害表現感病或高感病害。新疆野生甜瓜與甜瓜兩個亞種ssp.agrestis和ssp.melo可正常雜交且雜種后代可育,應當被劃分在C.melo.內。
References)
[1] Decker-Walters, D. S., Chung, S. M., Staub, J. E., Quemada, H. D., & López-Sesé, A. I. (2002). The origin and genetic affinities of wild populations of melon ( cucumis melo, cucurbitaceae) in North America.PlantSystematics&Evolution, 233(3-4): 183-197.
[2] Roy A, Bal S S, Fergany M, Kaur S, Singh H, Malik A A, Singh J, Monforte A J, Dhillon N P S. (2012). Wild melon diversity in India (Punjab State).GeneticResources&CropEvolution, (59): 755-767.
[3] Pitrat, M. (2013). Phenotypic diversity in wild and cultivated melons (cucumis melo).PlantBiotechnology, 30(3): 273-278.
[4] John, K. J., Scariah, S., Nissar, V. A. M., Latha, M., Gopalakrishnan, S., & Yadav, S. R., et al. (2013). On the occurrence, distribution, taxonomy and genepool relationship of cucumis callosus (rottler) cogn. the wild progenitor of cucumis melo l. from india.GeneticResources&CropEvolution, 60(3):1,037-1,046.
[5] Zhang, C., Pratap, A. S., Natarajan, S., Pugalendhi, L., Kikuchi, S., & Sassa, H., et al. (2012). Evaluation of morphological and molecular diversity among south asian germplasms of cucumis sativus and cucumis melo.ISMAgronomy, doi: 10.5402/2012/134134.
[6] Trimech, R., Zaouali, Y., Boulila, A., Chabchoub, L., Ghezal, I., & Boussaid, M. (2013). Genetic variation in tunisian melon (cucumismelo, L.) germplasm as assessed by morphological traits.GeneticResources&CropEvolution, 60(5):1,621-1,628.
[7] Hu, J., Wang, P., Su, Y., Wang, R., Li, Q., & Sun, K. (2015). Microsatellite diversity, population structure, and core collection formation in melon germplasm.PlantMolecularBiologyReporter, 33(3): 439-447.
[8] Tanaka, K., Stevens, C. J., Iwasaki, S., Akashi, Y., Yamamoto, E., & Dung, T. P., et al. (2016). Seed size and chloroplast dna of modern and ancient seeds explain the establishment of japanese cultivated melon (cucumismelo, L.) by introduction and selection.GeneticResources&CropEvolution, 63(7):1,237-1,254.
[9] Stepansky, A., Kovalski, I., & Perl-Treves, R. (1999). Intraspecific classification of melons (cucumismelo, L.) in view of their phenotypic and molecular variation.PlantSystematics&Evolution, 217(3-4): 313-332.
[10] Nesom, G. L. (2011). Toward consistency of taxonomic rank in wild/domesticated cucurbitaceae.Phytoneuron. (13):1-33.
[11] Hammer, K., & Gladis, T. (2014). Notes on infraspecific nomenclature and classifications of cultivated plants in compositae, cruciferae, cucurbitaceae, gramineae (with a remark on triticum dicoccon, schrank) and leguminosae.GeneticResources&CropEvolution, 61(8):1,455-1,467.
[12] Ali-Shtayeh, M. S., Jamous, R. M., Shtaya, M. J., Mallah, O. B., Eid, I. S., & Zaitoun, S. Y. A. (2010). Morphological characterization of snake melon ( cucumis melo, var. flexuosus ) populations from palestine.GeneticResources&CropEvolution,accepted: 1-16.
[13] Singh, A. K., & Yadava, K. S. (1984). An analysis of interspecific hybrids and phylogenetic implications in cucumis, ( cucurbitaceae ).PlantSystematics&Evolution, 147(3-4): 237-252.
[14] Alexander Beharav, & Yigal Cohen. (1995). Attempts to overcome the barrier of interspecific hybridization between and.IsraelJournalofPlantSciences, (2): 113-123.
[15] Sebastian, P., Schaefer, H., Telford, I. R., & Renner, S. S. (2010). Cucumber (cucumis sativus) and melon (c. melo) have numerous wild relatives in asia and australia, and the sister species of melon is from australia.ProceedingsoftheNationalAcademyofSciencesoftheUnitedStatesofAmerica, 107(32): 14,269-14,273.
[16] Matsumoto, Y., Miyagi, M., Watanabe, N., & Kuboyama, T. (2012). Temperature-dependent enhancement of pollen tube growth observed in interspecific crosses between wild cucumis, spp. and melon ( c. melo, l.).ScientiaHorticulturae, 138(138):144-150.
[17] Morales, M., Orjeda, G., Nieto, C., Leeuwen, H. V., Monfort, A., & Charpentier, M., et al. (2005). A physical map covering the nsv, locus that confers resistance to melon necrotic spot virus, in melon (cucumismelo, L.).Tag.theoretical&AppliedGenetics.theoretischeUndAngewandteGenetik, 111(5): 914-922.
[18] Boissot, N., Thomas, S., Sauvion, N., Marchal, C., Pavis, C., & Dogimont, C. (2010). Mapping and validation of qtls for resistance to aphids and whiteflies in melon.Theoretical&AppliedGenetics, 121(1): 9-20.
[19] 咸豐,張勇,馬建祥,等. 野生甜瓜'云甜-930'抗白粉病主基因+多基因遺傳分析[J]. 中國農業科學,2011,44(7):1 425-1 433.
XIAN Feng, ZHANG Yong, MA Jian-xiang, et al. (2011). Genetic Analysis of Resistant to Powdery Mildew with Mixed Modelof Major Gene Plus Polygene in Wild Melon Material 'Yuntian-930'[J].ScientiaAgriculturaSinica, 44(7):1,425-1,433. (in Chinese)
[20] Diaz, A., Fergany, M., Formisano, G., Ziarsolo, P., Blanca, J., & Fei, Z., et al. (2011). A consensus linkage map for molecular markers and quantitative trait loci associated with economically important traits in melon (cucumismeloL.).BmcPlantBiology, 11(1): 1-14.
[21] Malik, A. A., Vashisht, V. K., Singh, K., Sharma, A., Singh, D. K., & Singh, H., et al. (2014). Diversity among melon (cucumismeloL.) landraces from the indo-gangetic plains of india and their genetic relationship with usa melon cultivars.GeneticResources&CropEvolution, 61(6):1,189-1,208.
[22] 王堅. 中國西瓜甜瓜[M]. 北京:中國農業出版社,2000: 355-368.
WANG Jian. (2000).ChinaWatermelonandMuskmelon[M]. Beijing: China Agriculture Press: 355-368. (in Chinese)
[23] 張永兵,李寐華,吳海波,等. 新疆甜瓜地方品種資源的表型遺傳多樣性[J]. 園藝學報,2012,(39):305-314.
ZHANG Yong-bing, LI Mei-hua, WU Hai-bo, et al. (2012).Genetic diversity of melon landraces (CucumismeloL.) in Xinjiang based on phenotypic characters [J].ActaHorticulturaeSinica, (39): 305-314. (in Chinese)
[24] Renner, S. S., Schaefer, H., & Pitrat, M. (2008). Phylogenetics of Cucumis (Cucurbitaceae) as understood in 2008.Cucurbitaceae2008.ProceedingsoftheIxthEucarpiaMeetingonGeneticsandBreedingofCucurbitaceae,Avignon,France, 21-24 May.
[25] 吳明珠. 吐魯番盆地厚皮甜瓜品種資源及其利用[J]. 中國果樹,1982,(2):28-33.
WU Ming-zhu. (1982).Germplasm resources and utilization of Muskmelon in Turpan Basin [J].JournalofFruitsScience, 9(2): 28-33. (in Chinese)
[26] 林德佩. 新疆野生甜瓜的研究[J]. 新疆八一農學院學報,1984,(1):50-52.
LIN De-pei. (1984).Study on wild melon (Cucumis melo L.) in Xinjiang [J].JournalofBayiAgriculturalCollege, (1): 50-52. (in Chinese)
[27] Zhang, Y. P., Kyle, M., Anagnostou, K., & Zitter, T. A. (1997). Screening melon (cucumis melo) for resistance to gummy stem blight in the greenhouse and field.HortscienceAPublicationoftheAmericanSocietyforHorticulturalScience, 32(1):117-121.
[28]周曉慧,Joseph N. Wolukau,李英,等. 甜瓜抗蔓枯病種質資源的篩選及RAPD分析[J]. 園藝學報,2007,34(5):1 201-1 206.
ZHOU Xiao-hui, JOSEPH N. Wolukau, LI Ying, et al. (2007). Screening and RAPD analysis of gummy stem blight resistance in melon germplasm [J].ActaHorticulturaeSinica, 34(5):1,201-1,206. (in Chinese)
[29] Zhang, Y., Fan, X., Aierken, Y., Ma, X., Yi, H., & Wu, M. (2016). Genetic diversity of melon landraces ( cucumis melo, l.) in the xinjiang uygur autonomous region on the basis of simple sequence repeat markers.GeneticResources&CropEvolution: 1-13.
[30] Akashi, Y., Fukuda, N., Wako, T., Masuda, M., & Kato, K. (2002). Genetic variation and phylogenetic relationships in east and south asian melons, cucumis melo l. based on the analysis of five isoenzymes.Euphytica, 125(3): 385-396.
[31] Tanaka, K., Nishitani, A., Akashi, Y., Sakata, Y., Nishida, H., & Yoshino, H., et al. (2007). Molecular characterization of south and east asian melon, cucumis melo l. and the origin of group conomon var. makuwa and var. conomon revealed by rapd analysis.Euphytica, 153(1-2): 233-247.
Abstract:【Objective】 Through morphological observation, identification and cross compatibility between wild muskmelon and varieties inCucumismeloL. this project aims to provide a support for the classification of wild muskmelons in Xinjiang and the genetic base of melon cultivars.【Method】Twenty-eight traits of leaf, stem, flower and fruit of Xinjiang wild melon were observed and compared with local landraces, and the disease resistance of wild melon in Xinjiang to 2 fungi diseases: gummy stem blight (GSB) and powdery mildew was identified by inoculation technique at the seedling stage. Meanwhile, cross compatibility between Xinjiang wild melon and 8 varieties, in which 5 were included in ssp.agrestis, var.agrestis, var.conomon, var.dudaim, var.chitoand var.momordica, and 3 were included in ssp.melo, var.cantalupensis, var.inodorusand var.flexuosuswere analyzed, by the way of recording on fruit-setting and seed-setting rate of combinations after artificial pollination.【Result】Results obtained showed that most morphological traits of 'Hami wild' were distinct from the local cultivated control, but traits of 'Turpan wild' were similar to the control. 'Hami wild' was highly resistant to GSB but susceptible to PM, while 'Turpan wild' was susceptible or highly susceptible to GSB and PM after inoculation at seedling stage. Infertility of hybrids and incompatibility were not observed among any combinations between Xinjiang wild melon and 8 varieties. Except var.agrestisandchito, the seed-setting rate between reciprocal crosses was found differentially among combinations between Xinjiang wild melon and 8 varieties, which higher seed-setting rate was detected when Xinjiang wild melon was used as female parent. Furthermore, the seed-setting rate between 'Hami wild' and 2 subspecies, ssp.agrestisand ssp.melo, were 79.3% and 83.4%, respectively; and the seed-setting rate between 'Turpan wild' and 2 subspecies were 76.1% and 79.4%, respectively, which indicated that seed-setting rate exhibited small differences between combinations of Xinjiang wild melon with ssp.agrestisand with ssp.melo. 【Conclusion】Morphological traits observed in 'Hami wild' looked like that of feral or weedy melon in ssp.agrestis, however, 'Turpan wild' showed affinities with local landraces. 'Hami wild' was highly resistant to GSB after screening in seedlings. Xinjiang wild melon could be classified into C.melo, because of fertility of hybrids and compatibility were detected between wild melon and 2 subspecies, ssp.agrestis, ssp.melo.
Keywords: wild melon; morphology; seed-setting rate; cross compatibility
MorphologicalObservationonXinjiangWildMuskmelonsandStudyonCrossCompatibilitybetweenThemandDifferentVarietiesinCucumismeloL.
LI Mei-hua1,2, FAN Guo-quan3, YANG Yong1, WANG Guang-zhi1, ZHANG Jian4,ZHANG Xue-jun1, YI Hong-ping1, ZHANG Yong-bing1
(1.HamiMelonResearchCenter,XinjiangAcademyofAgriculturalSciences,Urumqi830091,China;2.CollegeofForestryandHorticulture,XinjiangAgriculturalUniversity,Urumqi830052,China;3.ScientificResearchManagementOffice,XinjiangAcademyofAgriculturalSciences,Urumqi830091,China;4.ZhejiangQuzhouXinnongduIndustryCo.,Ltd.,QuzhouZhejiang324022,China)
10.6048/j.issn.1001-4330.2017.09.008
S652.9
A
1001-4330(2017)09-1627-09
2017-03-19
國家自然科學基金項目“新疆甜瓜種質資源遺傳多樣性及野生甜瓜系統生物學研究”(31260480);國家現代農業產業技術體系建設專項“厚皮甜瓜育種崗”(CARS-26-04);“十二五”農村領域國家科技計劃研究任務“西甜瓜雜種優勢利用與新品種選育”(2012BAD02B03-4)
李寐華(1985-),男,湖南湘潭人,助理研究員,研究方向為西瓜甜瓜遺傳育種,(E-mail)mh0732@qq.com
張永兵(1971-),男,河南臨潁人,研究員,博士,研究方向為甜瓜遺傳育種與生物技術,(E-mail)luoybzhang@163.com
Supported by: National Natural Science Foundation of China "Study on Genetic Diversity of Melon (CucumismeloL.) Germplasm and Biological Characteristics of Wild Melon in Xinjiang" (31260480); Modern Agro-industry Technology Research System "Position of Breeding on Muskmelon"(CARS-26-04); National Science & Technology Pillar Program during the 12th Five-year Plan Period "Utilization of Heterosis and New Cultivar Selection of Muskmelon and Watermelon"(2012BAD02B03-4)
Corresponding author:ZHANG Yong-bing (1971-), male, native place: Linying Henan. Research Fellow. Ph.D, research field: Genetics, breeding and biotechnology of melon, (E-mail)luoybzhang@163.com
猜你喜歡
國畫家(2022年2期)2022-04-13 09:07:46
四川文學(2021年4期)2021-07-22 07:11:54
新作文·高中版(2021年4期)2021-07-14 11:55:06
環球人物(2021年8期)2021-05-06 08:00:43
音樂天地(音樂創作版)(2020年12期)2021-01-11 01:13:40
音樂天地(音樂創作版)(2020年7期)2020-09-11 06:17:18
華人時刊(2018年15期)2018-11-10 03:25:32
人民中國(日文版)(2015年10期)2015-04-16 03:53:48
絲綢之路(2014年9期)2015-01-22 04:24:46
新疆人文地理(2009年7期)2009-09-29 09:56:14
