尖峰嶺熱帶山地雨林林冠層喬木某些功能性狀的系統(tǒng)發(fā)育信號、關(guān)聯(lián)性及其演化模式
2017-10-13 12:32:55許格希史作民陳歡歡唐敬超馬凡強劉世榮李意德林明獻
生態(tài)學(xué)報 2017年17期
許格希, 史作民,2,*,劉 順, 陳歡歡, 唐敬超, 馬凡強, 許 涵, 劉世榮, 李意德, 林明獻
1 中國林業(yè)科學(xué)研究院森林生態(tài)環(huán)境與保護研究所國家林業(yè)局森林生態(tài)環(huán)境重點實驗室, 北京 100091 2 南京林業(yè)大學(xué)南方森林可持續(xù)經(jīng)營創(chuàng)新中心, 南京 210037 3 中國林業(yè)科學(xué)研究院熱帶林業(yè)研究所, 廣州 510520 4 中國林業(yè)科學(xué)研究院熱帶林業(yè)研究所試驗站, 樂東 572542
尖峰嶺熱帶山地雨林林冠層喬木某些功能性狀的系統(tǒng)發(fā)育信號、關(guān)聯(lián)性及其演化模式
許格希1, 史作民1,2,*,劉 順1, 陳歡歡1, 唐敬超1, 馬凡強1, 許 涵3, 劉世榮1, 李意德3, 林明獻4
1 中國林業(yè)科學(xué)研究院森林生態(tài)環(huán)境與保護研究所國家林業(yè)局森林生態(tài)環(huán)境重點實驗室, 北京 100091 2 南京林業(yè)大學(xué)南方森林可持續(xù)經(jīng)營創(chuàng)新中心, 南京 210037 3 中國林業(yè)科學(xué)研究院熱帶林業(yè)研究所, 廣州 510520 4 中國林業(yè)科學(xué)研究院熱帶林業(yè)研究所試驗站, 樂東 572542
熱帶雨林林冠層具有豐富的物種、功能和系統(tǒng)發(fā)育(譜系)多樣性,能夠顯著影響生態(tài)系統(tǒng)生物地球化學(xué)循環(huán),調(diào)節(jié)大氣水熱平衡,緩解氣候變化與人為干擾的負(fù)面作用。因此熱帶雨林林冠層功能和譜系生態(tài)學(xué)研究是目前群落生態(tài)學(xué)研究的熱點之一。選取海南尖峰嶺熱帶山地雨林3個1 hm2樣地的林冠層為研究對象,利用樣地林冠樹種清查數(shù)據(jù)基于APG III系統(tǒng)發(fā)育結(jié)構(gòu)重建樣地林冠層系統(tǒng)發(fā)育樹。利用Blomber′sK對和Pagel′sλ指標(biāo)結(jié)合系統(tǒng)發(fā)育獨立性比較法對組成林冠層喬木樹種的結(jié)構(gòu)、化學(xué)計量和水力學(xué)功能性狀(11個性狀)的系統(tǒng)發(fā)育信號、關(guān)聯(lián)性和演化模式進行研究,以探討物種親緣關(guān)系對尖峰嶺熱帶山地雨林林冠層多維功能性狀關(guān)聯(lián)性及其動態(tài)演化的影響。結(jié)果表明,有8個功能性狀(葉面積、葉厚度和潛在最大高度除外)具有顯著的系統(tǒng)發(fā)育信號(通過P<0.05的顯著性檢驗),Blomber′sK值和Pagel′sλ值分別介于0.202—0.392和0.277—0.847之間,表明尖峰嶺熱帶山地雨林林冠層功能性狀普遍存在系統(tǒng)發(fā)育保守性。在系統(tǒng)發(fā)育背景下,林冠層喬木樹種結(jié)構(gòu)、化學(xué)和水力學(xué)功能性狀在物種水平普遍存在顯著關(guān)聯(lián)性(P<0.05),表現(xiàn)出趨同或趨異進化;而且林冠層喬木隨物種分化其功能性狀分化模式大致呈水平“漏斗”狀,姐妹類群功能性狀間差異性在物種分化早期(大約1億2千萬年前至6000萬年前)明顯小于其在中后期(大約6000萬年前至今)的差異性,導(dǎo)致了林冠層性狀空間在中后期迅速膨大。然而林冠層主要功能性狀在系統(tǒng)發(fā)育樹內(nèi)部每一節(jié)點上姊妹類群分化產(chǎn)生的系統(tǒng)發(fā)育獨立性比較值絕大部分與零模型隨機模擬值并無顯著性差異。了解系統(tǒng)發(fā)育背景下林冠層功能性狀的權(quán)衡關(guān)系及其隨物種演化的分化模式與時間動態(tài)為進一步探究熱帶雨林林冠生態(tài)系統(tǒng)功能發(fā)揮奠定基礎(chǔ)。
功能性狀; 系統(tǒng)發(fā)育信號; 性狀分化; 林冠; 熱帶山地雨林; 尖峰嶺
Abstract: The tropical rainforest canopy has enormous diversity of species, function, and phylogeny. It can significantly affect biogeochemical cycles of ecosystem, which can regulate the atmospheric water-heat balance, as well as relieve the negative effect brought by climate change and human disturbance. Thus, functional and phylogenetic ecology of tropical rainforest canopy are among the most burning topics of research in community ecology. The present study was conducted in the forest canopy of three 1 hm2plots located in Jianfengling tropical montane rainforest on Hainan Island. We used inventory data set of canopy trees to construct a phylogenetic tree of forest canopy in these plots based on APG III phylogenetic structure. Indices of Blomber′sKand Pagel′sλ, combined with phylogenetic independent contrasts method, were used to study the phylogenetic signals, correlations, and evolved patterns of eleven structural, stoichiometric, and hydraulic traits (i.e. leaf area, leaf thickness, leaf dry matter content, specific leaf area, leaf total organism content, leaf nitrogen content, leaf phosphorus content, leaf potassium content, vein density, leaf water content, potential maximum height) of the canopy trees. Our goal was to explore the effect of phylogenetic relatedness between species on the correlations of multi-dimensional functional traits and their dynamic evolutionary patterns at the canopy layer in Jianfengling tropical montane rainforest. The results showed that eight out of the eleven functional traits evolved with significant phylogenetic signals (P<0.05), except for leaf area, leaf thickness, and potential maximum height; and their Blomber′sKand Pagel′sλvalues ranged from 0.202 to 0.392 and from 0.277 to 0.847, respectively. This indicated that phylogenetic conservatism commonly existed in these canopy functional traits in Jianfengling tropical montane rainforest. Furthermore, structural, stoichiometric, and hydraulic traits of canopy trees were universally associated with each other at the species level (P<0.05) after considering species′ phylogenetic relatedness, which demonstrated convergent or divergent evolution. The differentiated patterns of these functional traits displayed generally like a funnel-shape along with divergence of species. In addition, the variation of functional traits for sister nodes, occurring in the preliminary evolved phase (c. 120 to 60 million years ago), was weaker than that in the middle and latest phases (since about 60 million years ago), which indicated the rapid expansion of trait-space for the forest canopy in the latter ones. However, a majority of phylogenetic independent contrast values obtained from the divergence of sister taxa on each of the phylogenetic internal node, given of evolution of each trait, were non-significantly different against the randomly simulated values generated by a null model. In summary, understanding the trade-off relationship of functional traits and their divergent patterns and temporal dynamics with evolution is a foundation for further exploration of the ecosystem functioning for tropical rainforest canopy.
KeyWords: functional traits; phylogenetic signal; trait divergence; forest canopy; tropical montane rainforest; Jianfengling
植物功能性狀常用于表征植物生態(tài)位及其生存策略,是植物體獲取資源、分配光合產(chǎn)物及應(yīng)對氣候變化的直觀體現(xiàn)[1- 3]。植物功能性狀種間差異及其權(quán)衡對維持生態(tài)系統(tǒng)結(jié)構(gòu)、功能和動態(tài)具有重要意義[4]。森林生態(tài)學(xué)家沿環(huán)境梯度(海拔、養(yǎng)分、氣候)和演替過程在不同植被類型中對與結(jié)構(gòu)、化學(xué)和水力學(xué)等密切關(guān)聯(lián)的功能性狀進行廣泛采樣,試圖揭示森林生態(tài)系統(tǒng)的生理生態(tài)過程、物種共存機制、對氣候變化的響應(yīng)機制、植被演替動態(tài)特征等[3,5-7]。但是,以往的功能生態(tài)學(xué)研究多將物種視為彼此相互獨立,忽略了物種間的系統(tǒng)發(fā)育關(guān)系的差異性[8-10]。
近20年,隨著系統(tǒng)發(fā)育群落生態(tài)學(xué)的快速發(fā)展,局域群落尺度、區(qū)域尺度和全球尺度植物物種系統(tǒng)發(fā)育關(guān)系逐漸得到確認(rèn)[11- 13]。有越來越多的功能性狀研究在物種間系統(tǒng)發(fā)育背景(進化歷史)下分析性狀間的協(xié)同或耦合關(guān)系及其環(huán)境適應(yīng)性[3,14- 16]。有學(xué)者提出功能性狀系統(tǒng)發(fā)育生態(tài)位保守性假說,認(rèn)為近緣種應(yīng)該比親緣相疏物種在功能性狀上具有更高的相似性,其生態(tài)位更加相似[17]。因此在分析局域群落或者更大尺度植物區(qū)系功能性狀關(guān)聯(lián)性時往往需要提前檢驗功能性狀的系統(tǒng)發(fā)育信號[18],若功能性狀具有系統(tǒng)發(fā)育保守性,則應(yīng)在考慮物種間系統(tǒng)發(fā)育信息的背景下分析功能性狀間的耦合關(guān)系[19]。
光是影響熱帶雨林植物生長、發(fā)育和演化的最重要環(huán)境因子[20]。垂直方向上的光梯度常導(dǎo)致雨林植物功能性狀表現(xiàn)出明顯的種間和種內(nèi)差異性(表型可塑性),甚至是個體不同部位同一功能性狀的變異性。接受全光照的植物比長期遮蔭的植物具有更高的光合能力和單位面積葉氮含量,同時將更多的生物量分配到根系[21],而遮蔭植物將更多的光合產(chǎn)物分配到葉綠體和其他參與光捕獲器官,將更多的生物量分配到葉片和枝干,以實現(xiàn)更高的養(yǎng)分利用效率[22]。然而,至今仍然只有較少的研究回答充分光照條件下,在考慮群落組成物種系統(tǒng)發(fā)育結(jié)構(gòu)時,林冠層與植物結(jié)構(gòu)、化學(xué)和水分傳導(dǎo)相關(guān)的功能性狀的耦合關(guān)系在種間是否具有普遍性和一致性[23]。
熱帶雨林林冠層生物多樣性豐富,群落結(jié)構(gòu)復(fù)雜,對于生態(tài)系統(tǒng)生物地球化學(xué)循環(huán)、應(yīng)對干擾以及調(diào)節(jié)局域小氣候具有重要作用,其功能生態(tài)學(xué)研究是目前群落生態(tài)學(xué)研究的熱點問題之一[24]。國外對于熱帶雨林林冠功能生態(tài)學(xué)研究已經(jīng)開展有近十年的歷史[23,25],例如Asner和Martin對林冠喬木和藤本化學(xué)性狀研究發(fā)現(xiàn)喬木在葉片壽命和抵御干擾上投入更多,而藤本植物在光照和生長上的投入多于喬木[26]。但是國內(nèi)針對熱帶森林林冠層的生態(tài)學(xué)研究才剛起步[27]。基于此,本文結(jié)合海南島尖峰嶺熱帶山地雨林本底資料,對林冠層組成喬木樹種與養(yǎng)分獲取及分配、水分運輸以及抵御干擾密切相關(guān)的功能性狀進行采樣,結(jié)合林冠組成樹種系統(tǒng)發(fā)育結(jié)構(gòu)檢驗這些重要功能性狀的系統(tǒng)發(fā)育信號,利用系統(tǒng)發(fā)育獨立性比較法分析林冠層物種水平功能性狀間權(quán)衡耦合關(guān)系及其生物學(xué)意義,探討各功能性狀在系統(tǒng)發(fā)育樹上的內(nèi)部節(jié)點的分化模式及其時間動態(tài),以期為尖峰嶺熱帶山地雨林林冠系統(tǒng)發(fā)育結(jié)構(gòu)及其物種共存機制研究奠定基礎(chǔ)。
1 研究地概況
尖峰嶺林區(qū)(18°20′—18°57′ N,108°41′—109°12′ E)是海南島主要林區(qū)之一,是我國現(xiàn)存面積較大、保存完整的熱帶原始森林之一,具有非常高的植物多樣性。尖峰嶺林區(qū)現(xiàn)存的熱帶原始雨林主要分布在海拔650—1100 m的熱帶山地雨林中,年均溫19.8℃,年均降水量約2651.3 mm,最冷月和最熱月平均溫度為10.8℃和27.5℃,土壤類型主要為磚黃壤和潛黃壤[28]。
尖峰嶺生物多樣性動態(tài)監(jiān)測樣地(簡稱為“尖峰嶺大樣地”)位于尖峰嶺熱帶山地雨林區(qū)的五分區(qū)原始林內(nèi),樣地東西長1000 m,南北寬為600 m,總面積為60 hm2。尖峰嶺大樣地原點(西南角)坐標(biāo)為18°43′41.0″N,108°53′59.6″,海拔為870 m[29]。樣地內(nèi)植物調(diào)查于2013年初完成,共記錄木本、藤本胸徑DBH≥1 cm的植物484427株(包括萌生、分支、枯立木、倒木),其中存活植株439676株,存活藤本植株32183株,枯立木和倒木植株12568株。存活植株除極少數(shù)未確定種名的植株外(61株),其他植株分屬62科155屬290種,主要以樟科、殼斗科、茜草科和棕櫚科為最具優(yōu)勢科,同時榆科的白顏樹和山礬科山礬屬物種在大樣地中也經(jīng)常分布[30]。
2 研究方法
2.1 樣地選擇
根據(jù)尖峰嶺大樣地的地形地貌特征結(jié)合研究目的和生物學(xué)統(tǒng)計要求,在尖峰嶺大樣地西南-東北對角線上沿海拔梯度選取相距距離最遠(yuǎn)的3個1 hm2樣地。3塊樣地中具有明顯的海拔、坡度、凹凸度差異,局域生境異質(zhì)性相對明顯。將3塊1 hm2樣地劃分為75個20 m×20 m的樣方。3塊樣地中共計存活的胸徑DBH≥1 cm的木本植株22561株(不含萌生、分支),分屬56科128屬236種,分別占尖峰嶺大樣地胸徑(DBH)≥1 cm木本和藤本植物科屬種的91.8%、83.1%和81.4%,林冠優(yōu)勢種見重要值列表(表1)。
表1尖峰嶺熱帶山地雨林研究樣地林冠優(yōu)勢木本植物種類(重要值≥2.0)組成
Table1Compositionofdominantwoodyspecieswithimportantvaluelagerthan2.0ofcanopylayerforstudiedplotsinJianfenglingtropicalrainforest

物種名Species重要值Importancevalue相對胸高斷面積Relativebasalarea相對密度Relativedensity相對頻度Relativefrequency大葉蒲葵Livistonasaribus(Loureiro)MerrillexChevalier4.099.591.051.63白顏樹GironnierasubaequalisPlanchon3.705.223.792.10厚殼桂Cryptocaryachinensis(Hance)Hemsley3.572.705.952.07香果新木姜子NeolitseaellipsoideaC.K.Allen3.331.246.732.02海南蕈樹AltingiaobovataMerrill&Chun3.287.781.051.01海南韶子Nepheliumtopengii(Merrill)H.S.Lo3.172.115.332.07油丹AlseodaphnehainanensisMerrill2.684.301.951.79東方瓊楠BeilschmiediatungfangensisS.K.Lee&L.F.Lau2.651.974.151.82黃葉樹XanthophyllumhainanenseHu2.322.602.411.96東方肖欖PlateaparvifoliaMerrill&Chun2.031.492.611.99
2.2 樣品采集
2014年7—10月份進行采樣。根據(jù)Swenson等在熱帶雨林的研究,將DBH≥15 cm的喬木構(gòu)成的子群落定義為林冠層[31]。參考Pérez-Harguindeguy等功能手冊指南及結(jié)合林冠層的特點,最終選擇11個結(jié)構(gòu)、化學(xué)和水力學(xué)功能性狀[32]。這些功能性狀分別為葉面積(LA)、葉厚度(LT)、葉干物質(zhì)含量(LDMC)、比葉面積(SLA)、葉脈密度(VD)、葉含水量(LWC)、葉總有機碳含量(TOC)、葉氮含量(LNC)、葉磷含量(LPC)、葉鉀含量(LKC)和潛在最大樹高(Hmax)。
選取生長到冠層并能接受全光照的喬木植株,在其樹冠中上層向陽面采集能夠被陽光直接照射到的15片以上(15—150片)成熟、無病蟲害葉片,并快速裝袋帶回實驗室。3個1 hm2樣地共采集林冠物種個體樣品637份,分屬41科77屬131種,占3塊被研究樣地所有林冠群落喬木樹種(145種)的90.3%。對于林冠群落常見樹種,每個樹種采集5個以上個體(5—14個);對于樣地內(nèi)樹高普遍超過25m的樹種,例如紅錐(Castanopsishystrix)、海南槽裂木(Pertusadinametcalfii)和臀果木(Pygeumtopengii)等,則至少采集1個以上個體;對于林冠偶見種或稀有種則盡量采集所有個體(2—5個)。
2.3 功能性狀測定
將帶回實驗室的鮮葉洗凈擦干后用百分之一天平稱量鮮重(g);利用數(shù)顯游標(biāo)卡尺測量葉片厚度,每次測量選取5片葉片疊加、避開主脈和側(cè)脈在葉片中心位置處進行測量,每個個體至少對10張(10—25)葉片進行測量,利用測量總和除以被測葉片數(shù)量得到單葉厚度(mm)。將葉片平展利用Epson 10000x掃描儀(Seiko Epson Co., Nagano, Japan)掃描葉片電子圖像,通過ImageJ軟件(https://imagej.nih.gov/)計算葉面積(cm2)。隨后,將掃描后的樣品裝入信封并在65℃的條件下烘干72 h直至恒重。樣品烘干后取出稱量干重(mg),以計算葉干物質(zhì)含量(mg/g)和比葉面積(cm2/g)。
另外,選取5片鮮葉避開主脈和側(cè)脈在葉片基部、中部和頂端各裁剪直徑約為6 mm的葉盤(圓盤)。隨機選取8個葉盤在5%濃度的NaOH溶液里進行水煮腐蝕直至透明,利用2%濃度的次氯酸鈉進行漂白,并轉(zhuǎn)移到50%的乙醇和100%的無水乙醇里進行脫水,之后轉(zhuǎn)移到番紅精-乙醇溶液里進行充分染色,取出后轉(zhuǎn)移到二甲苯溶液中漂色,這樣葉脈完成染色。從中選取3個處理效果最好的葉盤用加拿大樹脂制作永久切片。在顯微鏡(Leica S8APO, Leica Microsystems Ltd., Wetzlar, Germany)下對切片放大20倍選取葉脈-背景對比最明顯的區(qū)域進行拍照,利用Leaf GUI軟件[33]處理葉脈照片并計算葉脈密度(mm/mm2)。
將烘干葉片磨碎、過篩以備化學(xué)功能性狀測定所用。利用大約0.15 g葉片粉末樣品通過高溫外熱重絡(luò)酸鉀氧化-容量法測量葉全碳含量(mg/g);利用0.3 g左右樣品通過凱氏定氮法(Kjeltec 2200, FOSS, Sweden)測定葉氮含量(mg/g);利用大約0.3 g葉片粉末樣品通過鉬銻抗比色法(Mo-Sb colorimetric method, UV- 755B, 上海精密科學(xué)儀器有限公司, 上海)測定葉磷含量(mg/g);再取約0.3 g樣品利用原子吸收分光光度法(atomic absorption spectrophotometry, Z- 2300, Hitachi Ltd., Tokyo, Japan)測定葉鉀含量(mg/g)。
葉水分含量(g H2O/cm2)根據(jù)葉鮮重、干重和葉面積計算而得,計算公式為:LWC=(鮮重×1000-干重)/葉面積。潛在最大樹高(m)來源于Flora of China (http://foc.eflora.cn/)和海南植物志檢索。
2.4 系統(tǒng)發(fā)育樹構(gòu)建
將3塊1 hm2樣地中胸徑DBH≥15 cm包含所有喬木物種的物種名錄信息輸入到Phylomatic平臺(http://phylodiversity.net/phylomatic/),基于APG III分類系統(tǒng)發(fā)育框架(phylomatic tree version:zanne2014)[12]獲得林冠群落科水平的系統(tǒng)發(fā)育結(jié)構(gòu)。在科水平下出現(xiàn)的屬多分枝以及在屬水平下的種多分枝問題利用R語言ape包里multi2di函數(shù)對系統(tǒng)發(fā)育樹多分枝(polytomies)進行隨機分解[34],最終生成二分枝結(jié)構(gòu)的系統(tǒng)發(fā)育樹。基于Congruification算法[35]估算林冠群落系統(tǒng)發(fā)育樹的枝長,利用Phylocom軟件(version 4.2)中的BLADJ模塊將這些進化枝長添加到進化樹上,生成具有進化枝長的系統(tǒng)發(fā)育樹(ultrametric tree)[36]。
2.5 數(shù)據(jù)處理與分析
尖峰嶺熱帶山地雨林林冠功能性狀進化受種間親緣關(guān)系影響程度可用功能性狀系統(tǒng)發(fā)育信號及其顯著性檢驗進行量度,其中Blomberg′sK(以下簡稱為K值)[37]和Pagel′sλ(以下簡稱為λ值)[38]是目前廣為采用的兩個系統(tǒng)發(fā)育信號量度指標(biāo)。當(dāng)K>1,表示觀測到的群落物種功能性狀進化比預(yù)期布朗運動(Brownian Motion)隨機進化結(jié)果具有更強的系統(tǒng)發(fā)育信號,即功能性狀比預(yù)期更易變;當(dāng)K<1表示物種功能性狀進化比預(yù)期的隨機進化具有更弱的系統(tǒng)發(fā)育信號,即功能性狀更不容易發(fā)生變化(系統(tǒng)發(fā)育保守性)。功能性狀的系統(tǒng)發(fā)育信號檢驗是通過將觀測得到的K值與系統(tǒng)發(fā)育樹物種進行999次隨機置換產(chǎn)生的模擬K值進行由小到大排序,排序值小于等于25或大于等于975則表明功能性狀在給定系統(tǒng)發(fā)育結(jié)構(gòu)下具有顯著的系統(tǒng)發(fā)育信號(顯著性水平α=0.05)。λ值通常介于0跟1之間,λ值越接近于0表明姊妹類群(sister taxa)性狀分化時間越長,其性狀進化比隨機布朗運動下的功能性狀進化產(chǎn)生更大變異性,具有更弱的系統(tǒng)發(fā)育信號;較大的λ值表示物種間功能性狀進化與預(yù)期布朗運動性狀隨機進化更加相似,即功能性狀具有相對更強的系統(tǒng)發(fā)育信號(系統(tǒng)發(fā)育保守性,功能性狀更加相似)。功能性狀λ值的顯著性通過似然比檢驗(likelihood ratio test)比較伴隨λ值產(chǎn)生的對數(shù)似然值(log-likelihood values)與λ值等于0時產(chǎn)生的對數(shù)似然值來實現(xiàn)[39]。
系統(tǒng)發(fā)育獨立性比較(phylogenetic independent contrasts, PICs)是在考慮物種間系統(tǒng)發(fā)育關(guān)系的基礎(chǔ)上量度功能性狀間的相互關(guān)系的有效方法,最早由Felsenstein提出[40],直到今天仍然廣泛應(yīng)用于功能生態(tài)學(xué)的研究當(dāng)中[41]。PICs通過生成每個功能性狀的系統(tǒng)發(fā)育樹內(nèi)部節(jié)點(internal nodes)的比較值(contrast values)來實現(xiàn)[42]。對于群落中的N個物種,則可生成(N- 1)個系統(tǒng)發(fā)育節(jié)點(phylogenetic nodes)比較值。這些比較值是由任意兩個系統(tǒng)發(fā)育節(jié)點下的姐妹類群(末端節(jié)點就是2個物種)的功能性狀差異性及其共有枝長權(quán)重而得。因此,可以從系統(tǒng)發(fā)育樹末梢(terminal)通過逐層遞推,計算系統(tǒng)發(fā)育樹上每個節(jié)點的比較值。利用每個節(jié)點比較值結(jié)合系統(tǒng)發(fā)育樹枝長信息,分析功能性狀分化與物種進化時間的關(guān)系。與某一功能性狀群落整體系統(tǒng)發(fā)育信號計算及顯著性檢驗相似,利用每個節(jié)點計算得到的比較值與隨機999次模擬生成的比較值進行排序可檢驗每個節(jié)點上功能性狀的分化產(chǎn)生的差異性比預(yù)期隨機產(chǎn)生的值顯著的大或小。
對林冠群落喬木樹種功能性狀值進行對數(shù)轉(zhuǎn)換,林冠主要功能性狀系統(tǒng)發(fā)育信號計算及其顯著性檢驗通過R語言中的phytools包[43]來實現(xiàn),利用ape包[34]計算功能性狀的獨立性比較值。將各個功能性狀兩兩之間對應(yīng)的節(jié)點比較值(統(tǒng)計學(xué)上相互獨立)分別進行Pearson相關(guān)性分析,正相關(guān)則表明某兩個功能性狀趨同進化,反之則是趨異進化。對系統(tǒng)發(fā)育樹上各個節(jié)點功能性狀比較值與其相應(yīng)的枝長信息進行回歸分析,若具有更長進化歷史(系統(tǒng)發(fā)育枝長更長)的物種性狀節(jié)點比較值更大而末端節(jié)點比較值更小,則表明群落性狀空間(trait space)在早期迅速被物種填充,且隨著時間推移逐漸被填滿,同時對每個節(jié)點姊妹類群功能性狀差異性進行顯著性檢驗[42]。
所有的數(shù)據(jù)處理和圖形繪制均在R里完成[44]。
3 結(jié)果與分析
3.1 物種系統(tǒng)發(fā)育關(guān)系及其功能性狀差異
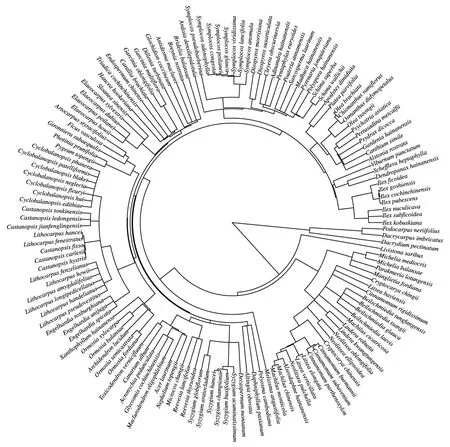
圖1 海南尖峰嶺熱帶山地雨林研究樣地林冠群落基于APG Ⅲ系統(tǒng)發(fā)育樹Fig.1 Phylogeny of canopy communities based on APG Ⅲ system in the studied plots of Jianfengling tropical montane rainforest on Hainan Island
從系統(tǒng)發(fā)育樹可以明確林冠群落組成喬木樹種的親緣關(guān)系,物種在系統(tǒng)發(fā)育樹上枝長越長則表示該物種的進化歷史更長(圖1)。從圖中可以看出尖峰嶺熱帶山地雨林林冠群落主要以真雙子葉植物為主,其中冬青科冬青屬的幾個種分化時間最短;同時擁有原始的基部被子植物木蘭科和樟科。此外,這些喬木樹種的冠層功能性狀種間差異量級具有差異性,其中LA、LPC、LKC和VD種間差異明顯,這些功能性狀物種水平最大值分別是最小值的大約38、33、11、10倍;LT、SLA、LNC、Hmax、LDMC以及LWC種間差異較明顯,最大值大致為物種最小值的3—4倍;TOC種間差異相對較小,在植物體中相對穩(wěn)定(表2)。
各功能性狀系統(tǒng)發(fā)育信號及其顯著性檢驗發(fā)現(xiàn)不同功能性狀受物種間親緣關(guān)系的影響程度不同。Blomber′sK法和Pagel′sλ法對功能性狀系統(tǒng)發(fā)育信號顯著性的檢驗結(jié)果相似,表明林冠主要功能性狀種間分化比預(yù)期隨機分化還要保守,表明親緣相近物種較親緣相疏物種具有更高的功能性狀相似性(表2)。因此,在分析功能性狀間的關(guān)聯(lián)性時有必要考慮物種間進化關(guān)系的影響。
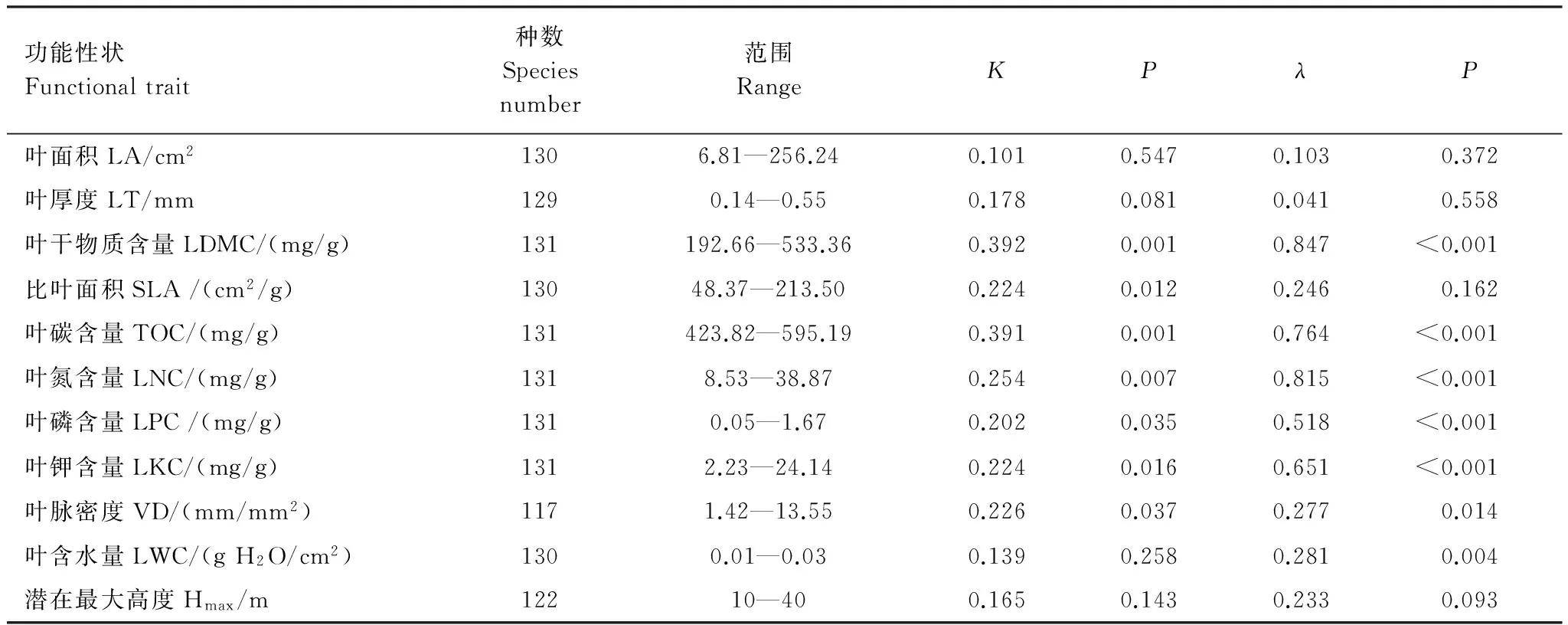
表2 海南尖峰嶺熱帶山地雨林林冠喬木主要功能性狀的系統(tǒng)發(fā)育信號
LA: leaf area; LT: leaf thickness; LDMC: leaf dry matter content; SLA: specific leaf area: TOC: leaf total organism content; LNC: leaf nitrogen content; LPC: leaf phosphorus content; LKC: leaf potassium content; VD: leaf vein density; LWC: leaf water content; Hmax: potential maximum height
3.2 林冠功能性狀系統(tǒng)發(fā)育獨立性比較及其關(guān)聯(lián)性
綜合姊妹類群功能性狀值及其進化枝長信息計算系統(tǒng)發(fā)育節(jié)點比較值,發(fā)現(xiàn)尖峰嶺熱帶山地雨林林冠群落物種水平功能性狀間大多表現(xiàn)為或者趨同進化或者趨異進化,且功能性狀間關(guān)聯(lián)程度不同。其中,LA與LT、LPC、LKC、LWC顯著正相關(guān)(LA與LKC相關(guān)性最強,相關(guān)系數(shù)為0.60),物種間功能性狀表現(xiàn)為趨同進化,同時與LDMC、SLA和LNC存在顯著負(fù)相關(guān)關(guān)系,表現(xiàn)為趨異進化。LT還表現(xiàn)為與LDMC、SLA、LNC和VD趨異進化(LT與SLA負(fù)相關(guān)關(guān)系最強,相關(guān)系數(shù)為-0.72),而與LWC趨同進化(相關(guān)系數(shù)為0.82)。LDMC表現(xiàn)出與SLA、LPC、LKC、VD和LWC趨異進化(LDMC與LKC和LWC密切相關(guān)),而與TOC和VD趨同進化(但相關(guān)性較弱);SLA除了與LA、LT和LDMC存在趨異進化外,還與TOC和LWC存在趨異進化,同時與養(yǎng)分元素LNC、LPC和LKC趨同進化;TOC除了與LA、LDMC和SLA存在進化上的較弱關(guān)聯(lián)外,與其他功能性狀不存在顯著的進化關(guān)聯(lián)。LNC還與LPC、VD和Hmax存在顯著的趨同進化關(guān)系,但與LWC存在顯著的趨異進化關(guān)系。LPC還與LKC存在極顯著的趨同進化關(guān)系,VD還與LWC存在顯著的趨異進化關(guān)系(表3)。其余未表現(xiàn)出顯著相關(guān)性的功能性狀意味著在考慮物種間系統(tǒng)發(fā)育非獨立性后功能性狀間不存在關(guān)聯(lián)性。
3.3 林冠樹種功能性狀分化模式及其時間動態(tài)
尖峰嶺熱帶山地雨林林冠群落組成喬木樹種不同功能性狀在系統(tǒng)發(fā)育樹上隨物種分化(各內(nèi)部節(jié)點)發(fā)生分化的模式相似但量級明顯不同。該地區(qū)林冠群落樹種功能性狀分化模式隨物種的分化大致呈現(xiàn)水平“漏斗”狀,絕大多數(shù)物種功能性狀分化發(fā)生在大約1億2千萬年前(-120 Ma)至3000萬年前(-30 Ma)。在該階段,物種分化后功能性狀種間差異較小(系統(tǒng)發(fā)育獨立性比較值較小),性狀空間相對較小。而自-30 Ma以來發(fā)生的物種分化,其功能性狀分化導(dǎo)致的性狀種間差異明顯,性狀空間迅速擴大,這在所有研究性狀均得到體現(xiàn)。此外,不同功能性狀隨物種分化其性狀差異量級不同(所有功能性狀均經(jīng)對數(shù)轉(zhuǎn)換),其中以LA隨物種分化差異量級最大,TOC差異量級最小,前者是后者的33倍以上。LDMC、SLA和LNC量級相似,比LA差異約小一個量級。其他功能性狀系統(tǒng)發(fā)育節(jié)點差異性與LA節(jié)點差異同一量級(圖2)。
表3海南尖峰嶺熱帶山地雨林林冠喬木主要功能性狀系統(tǒng)發(fā)育獨立性比較值Pearson相關(guān)性
Table3PearsoncorrelationamongPICsvaluesformajorfunctionaltraitsofcanopytreesofJianfenglingtropicalmontanerainforestonHainanIsland
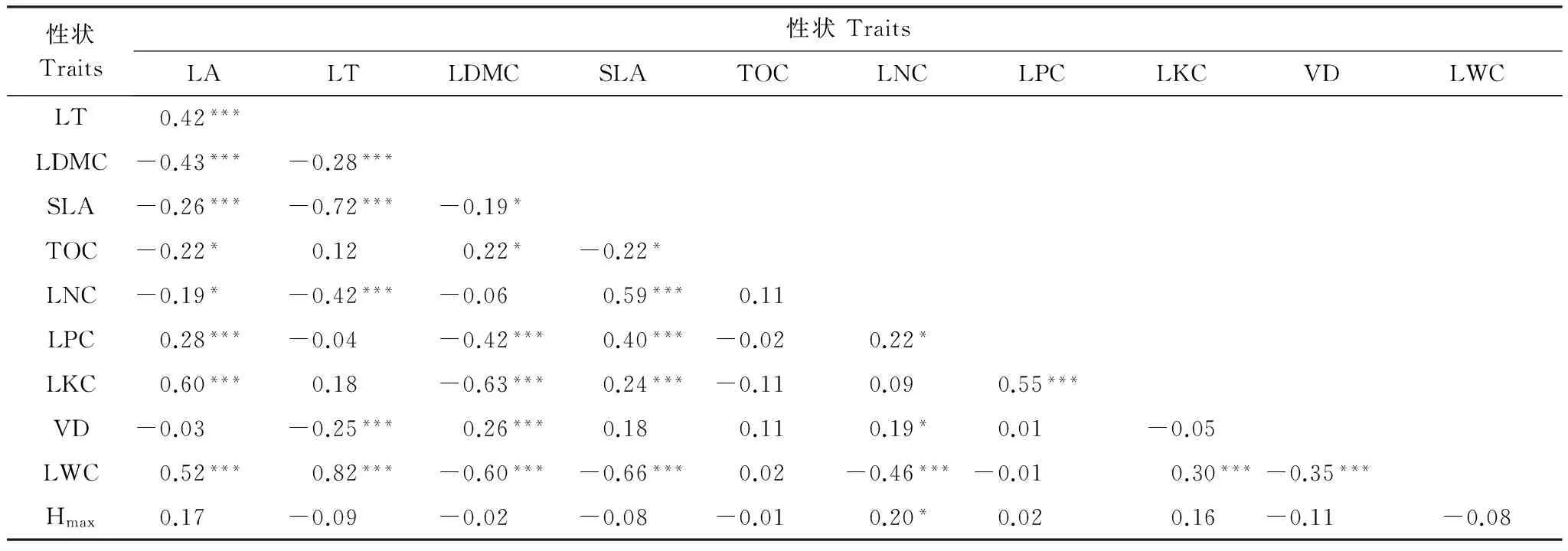
性狀Traits性狀TraitsLALTLDMCSLATOCLNCLPCLKCVDLWCLT0.42***LDMC-0.43***-0.28***SLA-0.26***-0.72***-0.19*TOC-0.22*0.120.22*-0.22*LNC-0.19*-0.42***-0.060.59***0.11LPC0.28***-0.04-0.42***0.40***-0.020.22*LKC0.60***0.18-0.63***0.24***-0.110.090.55***VD-0.03-0.25***0.26***0.180.110.19*0.01-0.05LWC0.52***0.82***-0.60***-0.66***0.02-0.46***-0.010.30***-0.35***Hmax0.17-0.09-0.02-0.08-0.010.20*0.020.16-0.11-0.08
大寫字母代表的功能性狀見表2;***P<0.001;***P<0.01;*P<0.05
但是,針對各功能性狀系統(tǒng)發(fā)育節(jié)點差異性的顯著性檢驗發(fā)現(xiàn)除了LA、LPC、LKC和Hmax在個別節(jié)點上表現(xiàn)為具有顯著性外,其他功能性狀在系統(tǒng)發(fā)育節(jié)點上的分化模式中與零模型預(yù)期的性狀隨機差異無顯著不同,表明系統(tǒng)發(fā)育樹上每個節(jié)點姊妹類群分化時功能性狀的分化在統(tǒng)計學(xué)上并未表現(xiàn)出顯著的保守性(圖3)。
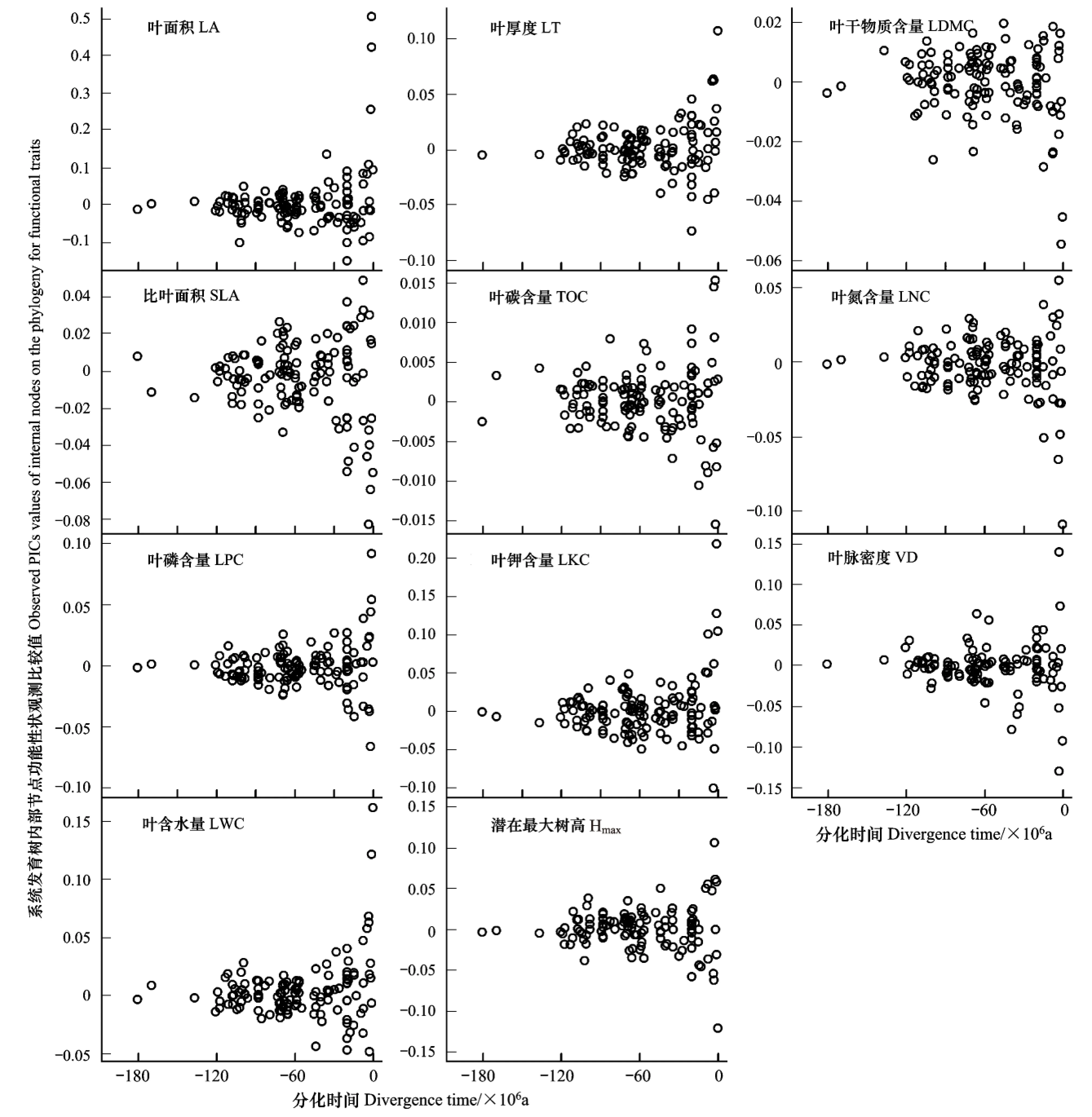
圖2 海南尖峰嶺熱帶山地雨林林冠喬木主要功能性狀分化模式及其時間動態(tài)Fig.2 Patterns and temporal dynamics of major traits divergence for canopy trees in Jianfengling tropical montane rainforest on Hainan IslandLA: leaf area; LT: leaf thickness; LDMC: leaf dry matter content; SLA: specific leaf area; TOC: leaf total organism content; LNC: leaf nitrogen content; LPC: leaf phosphorus content; LKC: leaf potassium content; VD: leaf vein density; LWC: leaf water content; Hmax: potential maximum height
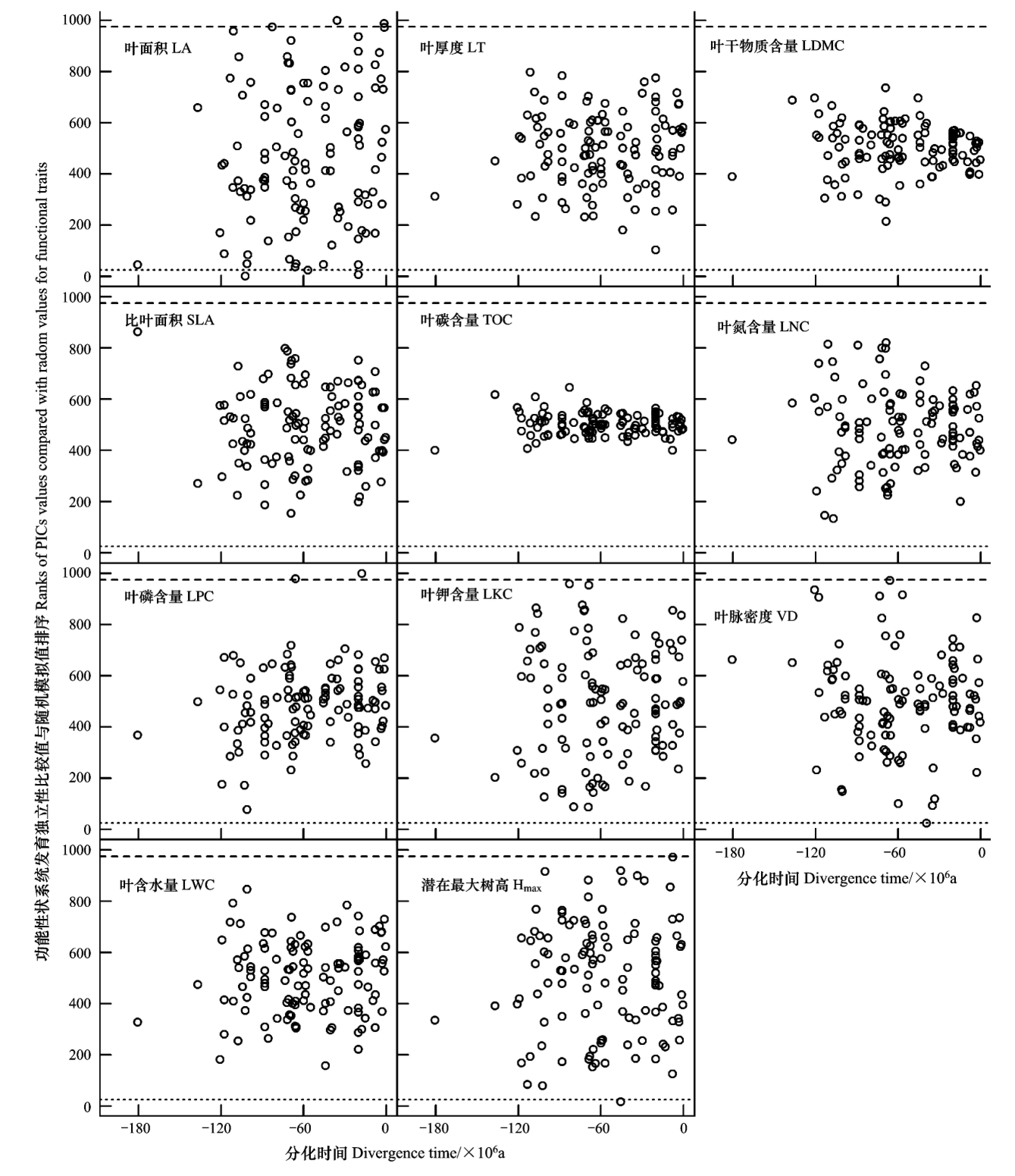
圖3 海南尖峰嶺熱帶山地雨林林冠喬木主要功能性狀姐妹類群節(jié)點分化差異性檢驗Fig.3 Test of variation generated by sister taxa divergence of major functional trais for canopy trees in Jianfengling tropical montane rainforest on Hainan Island各圖中散點代表各功能性狀在系統(tǒng)發(fā)育樹各內(nèi)部節(jié)點的系統(tǒng)發(fā)育獨立性比較值;虛線和短劃線分別代表具有系統(tǒng)發(fā)育信號和“反信號”的臨界值,分別低于或高于這兩個臨界值表明功能性狀分化差異性概率比預(yù)期隨機分化概率高或低
4 結(jié)論
本文針對生物多樣性極其豐富的海南島尖峰嶺熱帶山地雨林林冠層樹種某些與植物生理生態(tài)過程密切相關(guān)的功能性狀的系統(tǒng)發(fā)育保守性,功能性狀間物種水平權(quán)衡(trade-off)關(guān)系以及功能性狀隨物種演化的分化模式及其時間動態(tài)進行研究,為進一步探究群落物種共存機制奠定基礎(chǔ)。本研究主要結(jié)論如下:
(1)熱帶山地雨林林冠層組成樹種葉片功能性狀普遍具有系統(tǒng)發(fā)育保守性,即親緣相近物種比親緣相疏物種其功能性狀具有更高的相似性。
(2)林冠層葉片功能性狀在物種水平大多表現(xiàn)出協(xié)同進化模式,存在普遍的權(quán)衡關(guān)系。
(3)林冠層組成樹種隨著物種的演化其功能性狀分化呈水平“漏斗”狀,在物種演化初期(大約1億2千萬年前至6000萬年前)功能性狀種間差異較小,性狀空間相對較小;在物種演化中后期(大約6000萬年前至今)功能性狀種間差異逐漸增大,性狀空間逐漸擴展,并在演化后期隨著生物競爭加劇性狀空間迅速膨大。
5 討論
海南尖峰嶺熱帶山地雨林林冠層在結(jié)構(gòu)、化學(xué)和水力學(xué)性狀上的投入與其在資源獲取上存在權(quán)衡。遺傳特性、非生物過濾或隨機過程共同導(dǎo)致林冠某些功能性狀隨物種進化其分化模式表現(xiàn)出一定的相似性,這表現(xiàn)在功能性狀彼此之間在物種水平上普遍具有趨同或趨異現(xiàn)象。林冠層樹種對于葉片養(yǎng)分含量的投入越多,植物光合能力越強(更高的SLA),有利于獲取更多的資源產(chǎn)生更多的光合產(chǎn)物。有研究發(fā)現(xiàn)更低的SLA葉片具有更長的壽命[45],因此植物在葉片器官上的投入可能更多,這與本研究中發(fā)現(xiàn)林冠喬木樹種LT與SLA顯著負(fù)相關(guān)一致。而林冠樹種隨著對VD投入的增加,葉片對于昆蟲等食草動物的適口性變差,且葉片不容易受蒸發(fā)引起的水分喪失的脅迫,表現(xiàn)為植物葉片傾向于更薄且含水量更低[8]。表征植物光合作用能力的葉片經(jīng)濟性狀(例如SLA、LNC和LPC)與水力學(xué)性狀(例如VD和LWC)之間不存在或存在微弱的關(guān)聯(lián)性,這支持了Li等提出的葉片經(jīng)濟學(xué)和水力學(xué)相關(guān)性狀“模塊化”效應(yīng)或不同功能群性狀存在相對獨立軸向的假說[46]。林冠層主要樹種葉片功能性狀與樹高幾乎不存在任何關(guān)聯(lián),這可能是由于林冠層樹種普遍能夠捕獲充足的光照,導(dǎo)致樹高不是有效的表征種間及種內(nèi)個體差異性的性狀。
利用植物功能性狀解釋群落物種共存機制時普遍需要考慮物種的系統(tǒng)發(fā)育背景。在物種水平,簡單的對研究功能性狀進行相關(guān)或回歸分析必然會喪失物種重要的進化信息,結(jié)果在一定(或很大)程度上會與客觀功能性狀間的關(guān)系產(chǎn)生偏差。這種人為誤差在物種多樣性極高的熱帶森林功能性狀研究中更應(yīng)被納入考慮范圍[17]。本研究發(fā)現(xiàn)尖峰嶺熱帶山地雨林林冠層11個功能性狀普遍表現(xiàn)出顯著的系統(tǒng)發(fā)育信號(Blomber′sK值和Pagel′sλ值分別介于0.101—0.392和0.103—0.847),功能性狀隨物種分化而分化的過程具有系統(tǒng)發(fā)育保守性,表明親緣相近物種較親緣相疏物種具有更高的性狀相似性。這與Yang等在云南西雙版納和哀牢山熱帶雨林的研究結(jié)論相同[15]。然而,系統(tǒng)發(fā)育生態(tài)位保守性假說近年來受到一些生態(tài)學(xué)家的挑戰(zhàn)[47-48],經(jīng)常可以在群落中找到親緣相近但某些功能性狀差異顯著的植物類群,這些學(xué)者認(rèn)為環(huán)境過濾引起的植物表型可塑性以及隨機過程的影響可能超過了物種自身的遺傳作用對功能性狀的影響,進而導(dǎo)致近親物種產(chǎn)生性狀相異的現(xiàn)象[49]。許格希等研究發(fā)現(xiàn)尖峰嶺熱帶山地雨林林冠層某些功能性狀在土壤相對肥沃的生境條件下由于樹種競爭作用常導(dǎo)致功能性狀具有更高的多樣性;相反在相對貧瘠的生境下環(huán)境過濾對功能性狀的塑造作用更強,導(dǎo)致功能性狀具有更高的相似性[50]。
海南島尖峰嶺熱帶山地雨林林冠層物種水平功能性狀的分化隨物種分化大致呈現(xiàn)出水平“漏斗”狀,林冠層性狀空間是動態(tài)變化的多面體,在物種演化初期(1億2千萬年前—6000萬年前)生物競爭相對較弱,種間性狀差異較小,因此性狀空間相對亦較小。而在物種演化中后期(6000萬年前—至今),性狀空間隨種間競爭增強而逐漸膨脹。Swenson等認(rèn)為群落的性狀空間會在物種分化初期被迅速填充,而物種分化較晚的物種只能在原有性狀空間的基礎(chǔ)上去占據(jù)剩余的空間,性狀空間似乎是個靜態(tài)的“容器”[42]。本文的研究結(jié)果不認(rèn)為群落物種演化初期性狀空間會迅速被填充,反而因為資源的相對豐富且群落組成物種較少,物種傾向于具有更加相似的功能性狀。在物種分化的中期(6000萬年前—3000萬年前),隨著物種功能性狀的多元化,群落的性狀空間才會被迅速填充;在物種分化后期(3000萬年前至今),由于林冠層物種競爭排除作用激烈,導(dǎo)致后來的物種只能占據(jù)性狀空間的剩余部分或者利用性狀空間的邊緣部分并逐漸擴展群落的性狀空間以實現(xiàn)在群落中生存。
致謝:感謝所有參與海南尖峰嶺大樣地建設(shè)的科研工作者、老師、學(xué)生和當(dāng)?shù)毓と说男燎诟冻觥8兄x中國科學(xué)院植物研究所曾剛博士對于葉片解剖及葉脈觀測提供的指導(dǎo)與幫助。感謝熱帶林業(yè)研究所海南尖峰嶺試驗站楊懷博士、羅水興工程師、師國政工程師、南京森林警官學(xué)院周澗青警官、西北農(nóng)林科技大學(xué)的杜鳴溪和李明旭同學(xué)以及林澤彥對野外樣品處理提供的無私幫助,同時還要感謝黃啟雄、周建生和余遠(yuǎn)慶等當(dāng)?shù)鼐用袢虆⑴c并協(xié)助野外采樣工作的開展。
[1] McGill B J, Enquist B J, Weiher E, Westoby M. Rebuilding community ecology from functional traits. Trends in Ecology & Evolution, 2006, 21(4): 178- 185.
[2] Lasky J R, Uriarte M, Boukili V K, Erickson D L, Kress W J, Chazdon R L. The relationship between tree biodiversity and biomass dynamics changes with tropical forest succession. Ecology Letters, 2014, 17(9): 1158- 1167.
[3] Letcher S G, Lasky J R, Chazdon R L, Norden N, Wright S J, Meave J A, Pérez-García E A, Muoz R, Romero-Pérez E, Andrade A, Andrade J L, Balvanera P, Becknell J M, Bentos T V, Bhaskar R, Bongers F, Boukili V, Brancalion P H S, César R G, Clark D A, Clark D B, Craven D, DeFrancesco A, Dupuy J M, Finegan B, González-Jiménez E, Hall J S, Harms K E, Hernández-Stefanoni J L, Hietz P, Kennard D, Killeen T J, Laurance S G, Lebrija-Trejos E E, Lohbeck M, Martínez-Ramos M, Massoca P E S, Mesquita R C G, Mora F, Muscarella R, Paz H, Pineda-García F, Powers J S, Quesada-Monge R, Rodrigues R R, Sandor M E, Sanaphre-Villanueva L, Schüller E, Swenson N G, Tauro A, Uriarte M, van Breugel M, Vargas-Ramírez O, Viani R A G, Wendt A L, Williamson G B. Environmental gradients and the evolution of successional habitat specialization: a test case with 14 Neotropical forest sites. Journal of Ecology, 2015, 103(5): 1276- 1290.
[4] Soliveres S, Smit C, Maestre F T. Moving forward on facilitation research: response to changing environments and effects on the diversity, functioning and evolution of plant communities. Biological Reviews, 2015, 90(1): 297- 313.
[5] Díaz S, Kattge J, Cornelissen J H C, Wright I J, Lavorel S, Dray S, Reu B, Kleyer M, Wirth C, Prentice I C, Garnier E, B?nisch G, Westoby M, Poorter H, Reich P B, Moles A T, Dickie J, Gillison A N, Zanne A E, Chave J, Joseph Wright S, Sheremet′ev S N, Jactel H, Baraloto C, Cerabolini B, Pierce S, Shipley B, Kirkup D, Casanoves F, Joswig J S, Günther A, Falczuk V, Rüger N, Mahecha M D, Gorné L D. The global spectrum of plant form and function. Nature, 2016, 529(7585): 167- 171.
[6] Wright I J, Reich P B, Cornelissen J H C, Falster D S, Groom P K, Hikosaka K, Lee W, Lusk C H, Niinemets ü, Oleksyn J, Osada N, Poorter H, Warton D I, Westoby M. Modulation of leaf economic traits and trait relationships by climate. Global Ecology and Biogeography, 2005, 14(5): 411- 421.
[7] Anderson-Teixeira K J, Davies S J, Bennett A C, Gonzalez-Akre E B, Muller-Landau H C, Wright S J, Abu Salim K, Almeyda Zambrano A M, Alonso A, Baltzer J L, Basset Y, Bourg N A, Broadbent E N, Brockelman W Y, Bunyavejchewin S, Burslem D F R P, Butt N, Cao M, Cardenas D, Chuyong G B, Clay K, Cordell S, Dattaraja H S, Deng X, Detto M, Du X, Duque A, Erikson D L, Ewango C E N, Fischer G A, Fletcher C, Foster R B, Giardina C P, Gilbert G S, Gunatilleke N, Gunatilleke S, Hao Z, Hargrove W W, Hart T B, Hau B C H, He F, Hoffman F M, Howe R W, Hubbell S P, Inman-Narahari F M, Jansen P A, Jiang M, Johnson D J, Kanzaki M, Kassim A R, Kenfack D, Kibet S, Kinnaird M F, Korte L, Kral K, Kumar J, Larson A J, Li Y, Li X, Liu S, Lum S K Y, Lutz J A, Ma K, Maddalena D M, Makana J R, Malhi Y, Marthews T, Mat Serudin R, McMahon S M, McShea W J, Memiaghe H R, Mi X, Mizuno T, Morecroft M, Myers J A, Novotny V, De Oliveira A A, Ong P S, Orwig D A, Ostertag R, Den Ouden J, Parker G G, Phillips R P, Sack L, Sainge M N, Sang W, Sri-Ngernyuang K, Sukumar R, Sun I F, Sungpalee W, Suresh H S, Tan S, Thomas S C, Thomas D W, Thompson J, Turner B L, Uriarte M, Valencia R, Vallejo M I, Vicentini A, Vr?ka T, Wang X, Wang X, Weiblen G, Wolf A, Xu H, Yap S, Zimmerman J. CTFS-ForestGEO: a worldwide network monitoring forests in an era of global change. Global Change Biology, 2015, 21(2): 528- 549.
[8] Wright I J, Westoby M. Differences in seedling growth behaviour among species: trait correlations across species, and trait shifts along nutrient compared to rainfall gradients. Journal of Ecology, 1999, 87(1): 85- 97.
[9] Wright I J, Reich P B, Westoby M. Strategy shifts in leaf physiology, structure and nutrient content between species of high-and low-rainfall and high-and low-nutrient habitats. Functional Ecology, 2001, 15(4): 423- 434.
[10] Wright I J, Reich P B, Westoby M, Ackerly D D, Baruch Z, Bongers F, Cavender-Bares J, Chapin T, Cornelissen J H C, Diemer M, Flexas J, Garnier E, Groom P K, Gulias J, Hikosaka K, Lamont B B, Lee T, Lee W, Lusk C, Midgley J J, Navas M L, Niinemets U, Oleksyn J, Osada N, Poorter H, Poot P, Prior L, Pyankov V I, Roumet C, Thomas S C, Tjoelker M G, Veneklaas E J, Villar R. The worldwide leaf economics spectrum. Nature, 2004, 428(6985): 821- 827.
[11] Zhu Y, Comita L S, Hubbell S P, Ma K P. Conspecific and phylogenetic density-dependent survival differs across life stages in a tropical forest. Journal of Ecology, 2015, 103(4): 957- 966.
[12] Zanne A E, Tank D C, Cornwell W K, Eastman J M, Smith S A, FitzJohn R G, McGlinn D J, O′Meara B C, Moles A T, Reich P B, Royer D L, Soltis D E, Stevens P F, Westoby M, Wright I J, Aarssen L, Bertin R I, Calaminus A, Govaerts R, Hemmings F, Leishman M R, Oleksyn J, Soltis P S, Swenson N G, Warman L, Beaulieu J M. Three keys to the radiation of angiosperms into freezing environments. Nature, 2014, 506(7486): 89- 92.
[13] Smith S A, Beaulieu J M, Stamatakis A, Donoghue M J. Understanding angiosperm diversification using small and large phylogenetic trees. American Journal of Botany, 2011, 98(3): 404- 414.
[14] Ackerly D D, Schwilk D W, Webb C O. Niche evolution and adaptive radiation: testing the order of trait divergence. Ecology, 2006, 87(S7): S50-S61.
[15] Yang J, Ci X Q, Lu M M, Zhang G C, Cao M, Li J, Lin L X. Functional traits of tree species with phylogenetic signal co-vary with environmental niches in two large forest dynamics plots. Journal of Plant Ecology, 2014, 7(2): 115- 125.
[16] Pavoine S, Vela E, Gachet S, De Bélair G, Bonsall M B. Linking patterns in phylogeny, traits, abiotic variables and space: a novel approach to linking environmental filtering and plant community assembly. Journal of Ecology, 2011, 99(1): 165- 175.
[17] Losos J B. Phylogenetic niche conservatism, phylogenetic signal and the relationship between phylogenetic relatedness and ecological similarity among species. Ecology Letters, 2008, 11(10): 995- 1003.
[18] Mouquet N, Devictor V, Meynard C N, Munoz F, Bersier L F, Chave J, Couteron P, Dalecky A, Fontaine C, Gravel D, Hardy O J, Jabot F, Lavergne S, Leibold M, Mouillot D, Münkemüller T, Pavoine S, Prinzing A, Rodrigues A S L, Rohr R P, Thébault E, Thuiller W. Ecophylogenetics: advances and perspectives. Biological Reviews, 2012, 87(4): 769- 785.
[19] Garland T Jr, Harvey P H, Ives A R. Procedures for the Analysis of Comparative Data Using Phylogenetically Independent Contrasts. Systematic biology, 1992, 41(1): 18- 32.
[20] 蔡志全, 齊欣, 曹坤芳. 七種熱帶雨林樹苗葉片氣孔特征及其可塑性對不同光照強度的響應(yīng). 應(yīng)用生態(tài)學(xué)報, 2015, 15(2): 201- 204.
[21] Evans J R, Poorter H. Photosynthetic acclimation of plants to growth irradiance: the relative importance of specific leaf area and nitrogen partitioning in maximizing carbon gain. Plant, Cell & Environment, 2001, 24(8): 755- 767.
[22] Poorter H, Nagel O. The role of biomass allocation in the growth response of plants to different levels of light, CO2, nutrients and water: a quantitative review. Australian Journal of Plant Physiology, 2000, 27(6): 595- 607.
[23] Asner G P, Martin R E, Carranza-Jiménez L, Sinca F, Tupayachi R, Anderson C B, Martinez P. Functional and biological diversity of foliar spectra in tree canopies throughout the Andes to Amazon region. New Phytologist, 2014, 204(1): 127- 139.
[24] 馬克平. 生物多樣性科學(xué)的熱點問題. 生物多樣性, 2016, 24(1): 1- 2.
[25] Poorter L, Bongers L, Bongers F. Architecture of 54 moist-forest tree species: traits, trade-offs, and functional groups. Ecology, 2006, 87(5): 1289- 1301.
[26] Asner G P, Martin R E. Contrasting leaf chemical traits in tropical lianas and trees: implications for future forest composition. Ecology Letters, 2012, 15(9): 1001- 1007.
[27] Jin D M, Cao X C, Ma K P. Leaf functional traits vary with the adult height of plant species in forest communities. Journal of Plant Ecology, 2014, 7(1): 68- 76.
[28] 李意德, 許涵, 駱土壽, 陳德祥, 林明獻. 中國生態(tài)系統(tǒng)定位觀測與研究數(shù)據(jù)集·森林生態(tài)系統(tǒng)卷: 海南尖峰嶺站(生物物種數(shù)據(jù)集). 北京: 中國農(nóng)業(yè)出版社, 2012: 9- 50.
[29] 許涵, 李意德, 駱土壽, 陳德祥, 林明獻, 吳建輝, 李艷朋, 楊懷, 周璋. 海南尖峰嶺熱帶山地雨林——群落特征、樹種及其分布格局. 北京: 中國林業(yè)出版社, 2015: 17- 17.
[30] 許涵, 李意德, 林明獻, 吳建輝, 駱土壽, 周璋, 陳德祥, 楊懷, 李廣建, 劉世榮. 海南尖峰嶺熱帶山地雨林60ha動態(tài)監(jiān)測樣地群落結(jié)構(gòu)特征. 生物多樣性, 2015, 23(2): 192- 201.
[31] Swenson N G, Enquist B J, Thompson J, Zimmerman J K. The influence of spatial and size scale on phylogenetic relatedness in tropical forest communities. Ecology, 2007, 88(7): 1770- 1780.
[32] Pérez-Harguindeguy N, Díaz S, Garnier E, Lavorel S, Poorter H, Jaureguiberry P, Bret-Harte M S, Cornwell W K, Craine J M, Gurvich D E, Urcelay C, Veneklaas E, Reich P B, Poorter L, Wright I J, Ray P, Enrico L, Pausas J G, De Vos A C, Buchmann N, Funes G, Quétier F, Hodgson J G, Thompson K, Morgan H D, Ter Steege H, Van Der Heijden M G A, Sack L, Blonder B, Poschlod P, Vaieretti M V, Conti G, Staver A C, Aquino S, Cornelissen J H C. New handbook for standardised measurement of plant functional traits worldwide. Australian Journal of Botany, 2013, 61(3): 167- 234.
[33] Price C A, Symonova O, Mileyko Y, Hilley T, Weitz J S. Leaf extraction and analysis framework graphical user interface: segmenting and analyzing the structure of leaf veins and areoles. Plant Physiology, 2011, 155(1): 236- 245.
[34] Paradis E, Claude J, Strimmer K. APE: Analyses of phylogenetics and evolution in R language. Bioinformatics, 2004, 20(2): 289- 290.
[35] Eastman J M, Harmon L J, Tank D C. Congruification: support for time scaling large phylogenetic trees. Methods in Ecology and Evolution, 2013, 4(7): 688- 691.
[36] Webb C O, Ackerly D D, Kembel S W. Phylocom: software for the analysis of phylogenetic community structure and trait evolution. Bioinformatics, 2008, 24(18): 2098- 2100.
[37] Blomberg S P, Garland T Jr, Ives A R. Testing for phylogenetic signal in comparative data: Behavioral traits are more labile. Evolution, 2003, 57(4): 717- 745.
[38] Pagel M. The maximum likelihood approach to reconstructing ancestral character states of discrete characters on phylogenies. Systematic Biology, 1999, 48(3): 612- 622.
[39] Freckleton R P, Harvey P H, Pagel M. Phylogenetic analysis and comparative data: a test and review of evidence. The American Naturalist, 2002, 160(6): 712- 726.
[40] Felsenstein J. Phylogenies and the comparative method. The American Naturalist, 1985, 125(1): 1- 15.
[41] Comas L H, Callahan H S, Midford P E. Patterns in root traits of woody species hosting arbuscular and ectomycorrhizas: implications for the evolution of belowground strategies. Ecology and Evolution, 2014, 4(15): 2979- 2990.
[42] Swenson N G. Functional and Phylogenetic Ecology in R. New York: Springer, 2014: 147- 171.
[43] Revell L J. Phytools: an R package for phylogenetic comparative biology (and other things). Methods in Ecology and Evolution, 2012, 3(2): 217- 223.
[44] R Core Team. R: A language and environment for statistical computing. Vienna, Austria: The R Foundation for Statistical Computing, 2011. http://www.R-project.org.
[45] Sterck F J, Bongers F. Crown development in tropical rain forest trees: patterns with tree height and light availability. Journal of Ecology, 2001, 89(1): 1- 13.
[46] Li L, McCormack M L, Ma C G, Kong D L, Zhang Q, Chen X Y, Zeng H, Niinemets ü, Guo D L. Leaf economics and hydraulic traits are decoupled in five species-rich tropical-subtropical forests. Ecology Letters, 2015, 18(9): 899- 906.
[47] Pavoine S, Ricotta C. Functional and phylogenetic similarity among communities. Methods in Ecology and Evolution, 2014, 5(7): 666- 675.
[48] Pyron R A, Costa G C, Patten M A, Burbrink F T. Phylogenetic niche conservatism and the evolutionary basis of ecological speciation. Biological Reviews, 2015, 90(4): 1248- 1262.
[49] Eichenberg D, Ristok C, Kr?ber W, Bruelheide H. Plant polyphenols-implications of different sampling, storage and sample processing in biodiversity-ecosystem functioning experiments. Chemistry and Ecology, 2014, 30(7): 676- 692.
[50] 許格希, 史作民, 唐敬超, 劉順, 馬凡強, 許涵, 劉世榮, 李意德. 海南尖峰嶺熱帶山地雨林林冠層樹種功能多樣性特征. 應(yīng)用生態(tài)學(xué)報, 2016, 27(11): 3444- 3454.
Phylogeneticsignals,correlations,andevolutionarypatternsofsomefunctionaltraitsforforestcanopytreesinJianfenglingtropicalmontanerainforest
XU Gexi1, SHI Zuomin1,2,*, LIU Shun1, CHEN Huanhuan1, TANG Jingchao1, MA Fanqiang1, XU Han3, LIU Shirong1, LI Yide3, LIN Mingxian4
1KeyLaboratoryonForestEcologyandEnvironmentalSciences,StateForestryAdministration,ResearchInstituteofForestEcology,EnvironmentandProtection,ChineseAcademyofForestry,Beijing100091,China2Co-InnovationCentreforSustainableForestryinSouthernChina,NanjingForestryUniversity,Nanjing210037,China3ResearchInstituteofTropicalForestry,ChineseAcademyofForestry,Guangzhou510520,China4ExperimentalStationofResearchInstituteofTropicalForestry,ChineseAcademyofForestry,Ledong572542,China
國家自然科學(xué)基金資助項目(31290223, 31570240)
2016- 06- 13; < class="emphasis_bold">網(wǎng)絡(luò)出版日期
日期:2017- 04- 24
10.5846/stxb201606131132
*通訊作者Corresponding author.E-mail: shizm@caf.ac.cn
許格希, 史作民,劉順, 陳歡歡, 唐敬超, 馬凡強, 許涵, 劉世榮, 李意德, 林明獻.尖峰嶺熱帶山地雨林林冠層喬木某些功能性狀的系統(tǒng)發(fā)育信號、關(guān)聯(lián)性及其演化模式.生態(tài)學(xué)報,2017,37(17):5691- 5703.
Xu G X, Shi Z M, Liu S, Chen H H, Tang J C, Ma F Q, Xu H, Liu S R, Li Y D, Lin M X.Phylogenetic signals, correlations, and evolutionary patterns of some functional traits for forest canopy trees in Jianfengling tropical montane rainforest.Acta Ecologica Sinica,2017,37(17):5691- 5703.
猜你喜歡
課堂內(nèi)外·初中版(科學(xué)少年)(2025年1期)2025-02-28 00:00:00
課堂內(nèi)外·初中版(科學(xué)少年)(2025年2期)2025-02-28 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中華詩詞(2022年6期)2022-12-31 06:41:24
科學(xué)大眾(中學(xué))(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
中國科技論壇(2017年7期)2017-07-25 08:49:53
媽媽寶寶(2017年2期)2017-02-21 01:21:24
國際漢語學(xué)報(2016年1期)2017-01-20 08:21:20
少兒科學(xué)周刊·少年版(2015年1期)2015-07-07 17:15:12
