柳枝稷體外誘導花器官逆轉與非逆轉穗芽生理特性差異研究
2017-09-13 05:18:26王勇鋒崔桂賓徐開杰孫風麗劉曙東奚亞軍
草地學報 2017年4期
關鍵詞:差異
王勇鋒, 李 毛, 崔桂賓, 徐開杰, 孫風麗, 張 超, 劉曙東, 奚亞軍*
(1. 西北農林科技大學農學院, 陜西 楊凌 712100; 2. 中國農業科學院棉花研究所, 河南 安陽 455000)
開花是一個復雜的生命進程,其發生受到光周期、溫度、春化作用、植物激素等多因素的影響[1-5]。正常情況下,植物個體成熟以后即可進行開花、授粉、結實等一系列的生命過程,然而某些植物在進入成熟期后遇到不利于開花的外界環境,如持續降雨[6]、氣溫驟變[7]、人為改變光周期[8]等,部分已分化并發育的花器官組織將停止發育或逆向發育,最終再次形成營養器官或類似營養器官的組織,這種現象被稱為花器官逆轉。花器官逆轉根據逆轉來源不同又被分為花逆轉(花的部分結構發生逆轉)和花序逆轉(花序上的小花發生逆轉)[7,9]。花器官逆轉現象最早在鳳仙花(Impatiensbalsamina)中有詳細報道,研究發現將開花期的鳳仙花從短日照環境轉移到長日照環境下,花序終端將由花轉變成葉[7,9]。模式植物擬南芥(Arabidopsisthaliana)在短日照條件下也有部分小花逆向發育形成花序的現象,然而該現象只在其Landsberg erecta生態型中發現,且逆轉頻率不高,而其突變體lfy-6和ag-1在短日照條件下發生逆轉頻率升高[8,9]。在單子葉植物中同樣有花器官逆轉相關報道,Joseph等、Wang等分別研究發現,玉米(Zeamays)突變體id1無雄花發育[10],水稻(Oryzasativa)突變體pho不能形成正常小花[6],相應的花器官均發育形成新生幼苗。
在影響花發育的眾多因素中,植物激素被認為是一種至關重要的因子。在前人的研究中發現,多數影響到花發育的環境條件(光周期、溫度等)在誘導花器官形成的過程中都會引起植物體內激素水平變化[4,5,11]。Kesy等發現短日照植物矮牽牛(Pharbitisnil.)在開花前人為打斷黑暗條件將會顯著抑制開花,研究表明該過程中植物體內生長素(IAA)和乙烯(Eth)含量急劇升高,并且外源施加IAA和Eth同樣能起到抑制開花的效果[1];Sringarm等研究表明龍眼樹(Dimocarpuslongan, Lour.)在低溫誘導開花的過程中內源異戊烯基腺嘌呤/異戊烯基腺苷(iP/iPA)和吲哚乙酸(IAA)含量急劇升高,而玉米素(ZR)和赤霉素(GA)含量卻明顯降低[3]。細胞分裂素(CTK)在花發育過程中也起到重要的作用,多數研究者認為CTK在植物開花過程中起到促進作用[5,12],然而也有研究認為,細胞分裂素含量過高時,不利于植物從營養生長向生殖生長轉變[13]。
柳枝稷(PanicumvirgatumL.)是短日照開花植物[14-15],在陜西楊凌其抽穗期為每年7-8月,不同緯度間進行引種會引起開花期的改變。之前的研究中我們發現,將柳枝稷幼穗置于含有高水平外源細胞分裂素的培養基上進行體外培養時,部分已分化的花器官停止生殖生長,轉而逆向發育,經花器官逆轉途徑形成逆轉穗芽[16]。人們對花器官逆轉的研究由來已久,然而之前的研究多集中在花器官逆轉的誘導方法以及現象描述等方面[17-18],很少有人對花器官逆轉的生理特性進行較為詳細的研究。柳枝稷體外花器官逆轉誘導穗芽與前人通過改變環境誘導花器官逆轉的方法相比具有易操作、效率更高、結果更穩定等優勢[9,19]。本文參照前人研究,結合本實驗室前期柳枝稷人工穗芽誘導相關工作,從生理學方面對柳枝稷花器官逆轉現象進行探索,以期在一定程度上揭示柳枝稷花器官逆轉發生的機理,為柳枝稷及其他植物花器官逆轉研究和花發育研究提供參考。
1 材料與方法
1.1 試驗材料
柳枝稷為異花授粉植物,同一品種不同植株間性狀存在一定差異[14,20]。為驗證體外誘導花器官在不同類型柳枝稷材料中的一致性,試驗以柳枝稷栽培品種“Alamo”兩個植株為材料,編號分別為“4-6”和“14-7”,其中“14-7”與“4-6”相比植株更高大且分蘗更集中。材料種植于溫室中,每天光照16 h,溫度為恒溫30℃。
1.2 柳枝稷穗芽誘導
當柳枝稷發育至伸長期第4期(E4期)時[21],莖端發育出1~2 cm長幼穗,取下莖端(保留幼穗上下各2 cm),先用體積分數70%的酒精消毒1 min,然后用體積分數8%的次氯酸鈉(有效氯5.5%)再次消毒1~2 min,無菌水沖洗3~4次[16]。將處理后的材料在超凈工作臺中切去兩端各0.5 cm,并將其縱向切成兩半,接種到穗芽誘導培養基(Shoot Induction Medium, SIM)或MS基本培養基(Murashige and Skoog, MS)[22]上,之后置于25℃恒溫光照培養箱中培養,光周期為20 h白天/4 h黑暗。來自于兩棵柳枝稷單株的試驗材料分別培養和研究。
MS培養基:MS培養基無機鹽和有機物 + 30 g·L-1蔗糖 + 7 g·L-1瓊脂粉,pH 5.8;
SIM培養基:MS培養基無機鹽和有機物 + 3 mg·L-16-BA + 30 g·L-1蔗糖 + 7 g·L-1瓊脂粉,pH 5.8。
1.3 穗芽繼代培養
穗芽誘導培養30 d后,將材料在超凈工作臺中取出,切除芽塊上部葉片,留下基部1~1.5 cm,然后將其接種到新鮮培養基中。繼代培養所使用培養基與穗芽誘導培養一致,仍為SIM培養基或MS基本培養基。之后再次將材料置于25℃恒溫光照培養箱中培養,光周期保持20 h白天/4 h黑暗不變。繼代培養30 d后(共培養60 d),再次將材料取出并重復一次繼代培養。
1.4 生理指標測定及方法
試驗使用裂區設計,其中培養基類型(SIM和SM)和柳枝稷材料(4-6和14-7)為主處理,誘導時間(30 d,60 d,90 d)為副處理,試驗材料每30 d繼代一次。取不同處理的柳枝稷穗芽分別進行生理指標的測定,并進行主處理和副處理的差異性分析,每個處理重復3次。測定的生理指標包括可溶性糖含量[23]、游離氨基酸含量[24]、可溶性蛋白含量、總氮含量、葉綠素含量、超氧化物歧化酶(SOD)活性、過氧化物酶(POD)活性、過氧化氫酶(CAT)活性、多酚氧化酶(PPO)活性[25]。利用酶聯免疫法對不同處理3個培養時期的柳枝稷穗芽測定生長素(IAA)、赤霉素(GAs)、細胞分裂素(ZRs)和脫落酸(ABA)含量,并進行處理間和培養時期間的差異性分析,每個樣品重復3次。
1.5 數據分析
數據整理及圖表繪制使用Excel 2013,數據統計分析使用軟件DPS 7.05。
2 結果與分析
2.1 柳枝稷穗芽誘導特點
當柳枝稷4-6和14-7在SIM培養基和MS基本培養基上進行誘導培養30 d以后,不同處理的材料在形態上出現了明顯的差異。外植體在SIM培養基上(圖1c,1e)的生長速率較MS基本培養基(圖1d,1f)更快,產生逆轉穗芽數量大;與MS培養基上的非逆轉穗芽相比,逆轉穗芽長度較短,但整體更粗壯,節間更密集;在MS基本培養基上培養30 d后,幼穗繼續生長發育,長出小花(圖1d,1f),與之相比, SIM培養基上幼穗的小花發育更為緩慢或出現花器官逆轉現象(圖1c,1e);柳枝稷4-6和14-7逆轉穗芽誘導也表現出一定的差異,14-7生長速度較4-6更快,逆轉穗芽產生數量也較4-6更多,表明其具有更強的活性。

圖1 柳枝稷穗芽誘導Fig.1 Spike buds induction of switchgrass
2.2 柳枝稷穗芽發育中葉綠素含量的變化
培養30 d后,兩種柳枝稷材料的逆轉穗芽和非逆轉穗芽葉綠素含量并未發現明顯差異;60 d后,兩種柳枝稷逆轉穗芽均發生明顯的葉綠素含量降低的現象,而與此同時,非逆轉穗芽葉綠素含量并未發生明顯下降,方差分析結果顯示逆轉穗芽葉綠素含量極顯著低于非逆轉穗芽;與此類似,培養90 d的逆轉穗芽葉綠素含量極顯著低于非逆轉穗芽(表1)。兩種類型柳枝稷葉綠素含量差異不顯著(P= 0.5365),但就不同時期而言,培養30 d的材料與培養60 d和90 d的材料差異極顯著(P< 0.01),但后兩者差異不顯著(表1)。
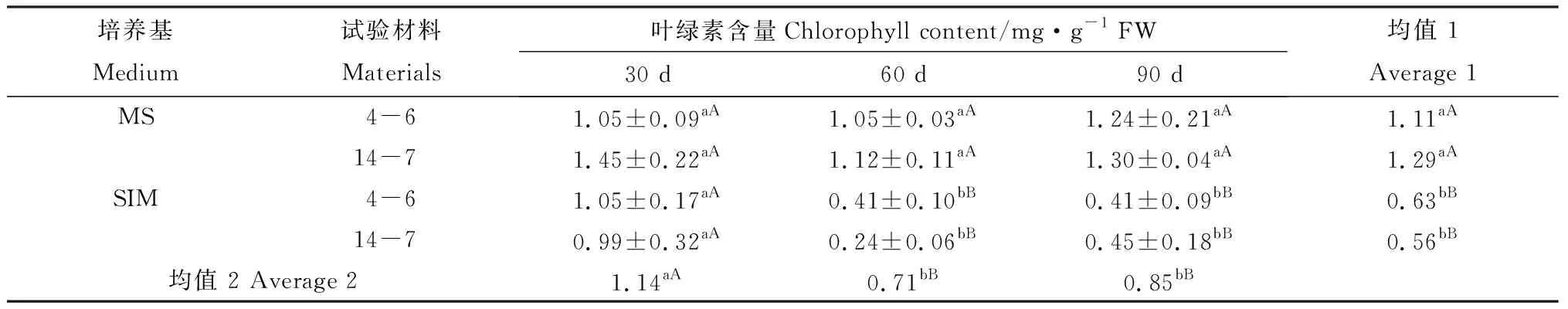
表1 柳枝稷穗芽形成期間葉綠素含量變化Table 1 Chlorophyll content variation of different switchgrass spike buds
2.3 柳枝稷穗芽發育中總糖含量的變化
兩種類型柳枝稷的逆轉穗芽和非逆轉穗芽總糖含量在培養30 d、60 d和90 d時差異均未達到顯著水平,這表明外植體在各時期不同培養基上對糖的吸收、代謝差異不明顯(表2)。兩種柳枝稷之間比較總糖含量差異未達到顯著水平(P=0.5113),但隨著培養時間的延長,總糖含量表現下降的趨勢,培養90 d的穗芽總糖含量極顯著低于培養30 d的穗芽(P< 0.01,表2)。這可能是因為多次繼代的穗芽塊具有更多的穗芽,對培養基中糖的消耗更快,從而引起后期培養基碳源供應不足。
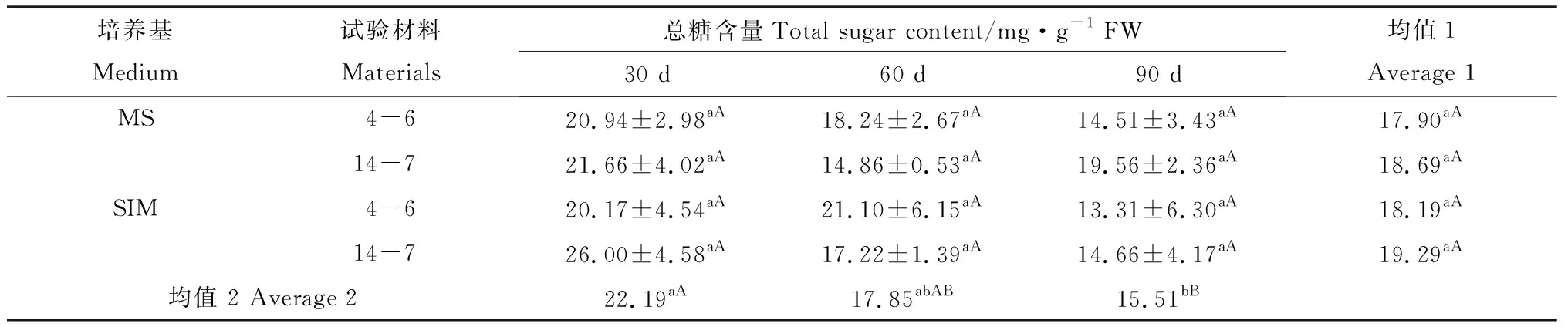
表2 柳枝稷穗芽形成期間總糖含量變化Table 2 Total sugar content variation of different switchgrass spike buds
2.4 柳枝稷穗芽發育中可溶性蛋白、游離氨基酸及總氮含量的變化
在培養30 d時,逆轉穗芽可溶性蛋白含量總體高于非逆轉穗芽,但只有柳枝稷14-7達到顯著水平(表3);與之相反,同時期的逆轉穗芽游離氨基酸的含量總體低于非逆轉穗芽,且均達到顯著水平(表4),而總氮含量差異不顯著(表5),這表明逆轉穗芽和非逆轉穗芽在培養早期對氮的吸收速率差異不明顯,但逆轉穗芽氮的代謝效率更高。培養60 d時,逆轉穗芽可溶性蛋白含量總體低于非逆轉穗芽,柳枝稷14-7達到顯著水平(表3),然而游離氨基酸和總氮含量差異不顯著(表3,4),這可能是因為繼代后逆轉穗芽數量和生長速度更快,對氮的消耗更快,從而表現出培養基氮缺乏現象。培養90 d時,逆轉穗芽可溶性蛋白含量總體低于非逆轉穗芽,游離氨基酸和總氮含量表現出相同的趨勢(表3,4),這可能是因為繼代培養對氮的消耗進一步加快,從而表現出更嚴重的培養基氮缺乏現象。兩種柳枝稷之間的可溶性蛋白、游離氨基酸和總氮含量差異均不顯著(P=0.0608,P=0.0611,P=0.4494),而逆轉穗芽與非逆轉穗芽之間均表現顯著或極顯著差異(P=0.0404,P=0.0048,P=0.0024),各時期間相比較可溶性蛋白含量整體下降,而游離氨基酸和總氮含量均為先升后降的趨勢。綜合考慮可溶性蛋白、游離氨基酸和總氮含量的變化以及SIM培養基后期氮供應不足可以推斷出逆轉穗芽具有更高的氮代謝效率。
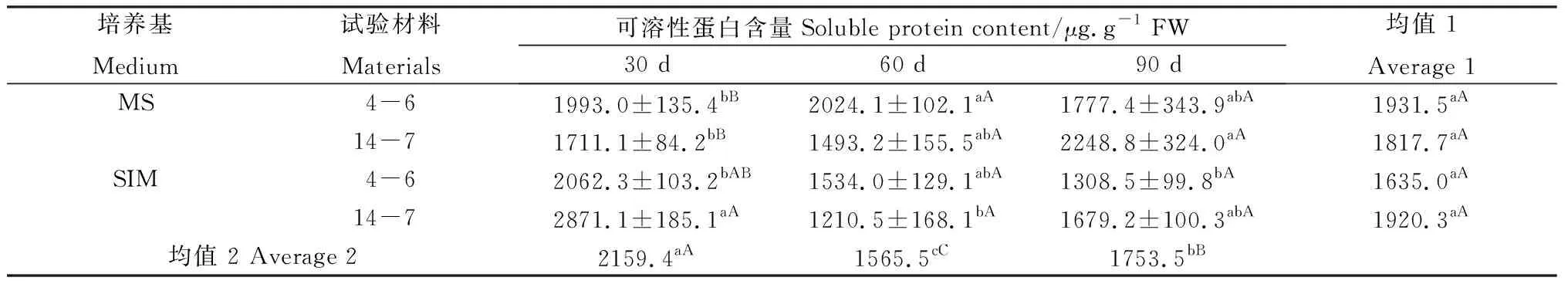
表3 柳枝稷穗芽形成期間可溶性蛋白含量變化Table 3 Soluble protein content variation of different switchgrass spike buds
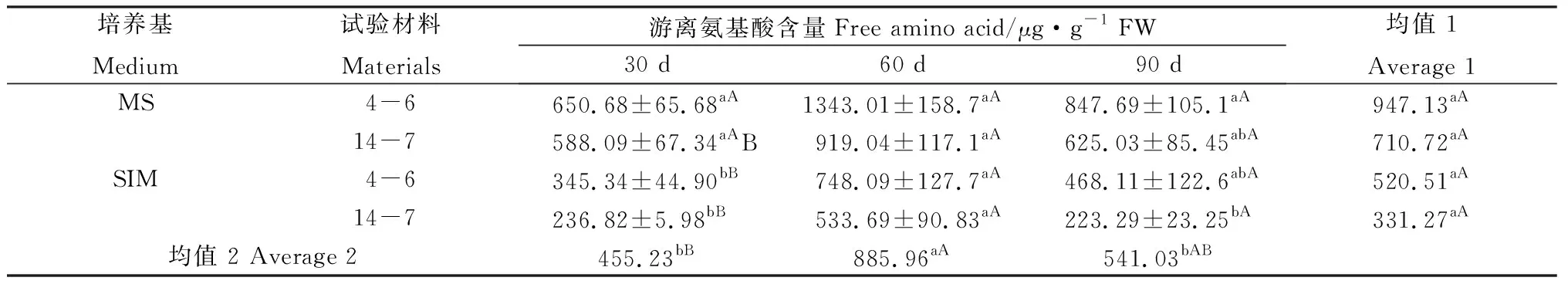
表4 柳枝稷穗芽形成期間游離氨基酸含量變化Table 4 Free amino acid content variation of different switchgrass spike buds
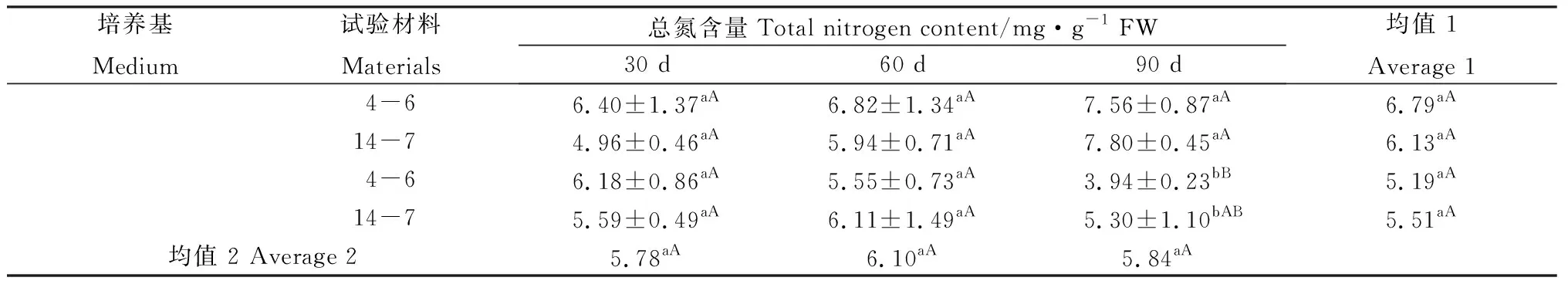
表5 柳枝稷穗芽形成期間總氮含量變化Table 5 Total nitrogen content variation of different switchgrass spike buds
2.5 柳枝稷穗芽發育中SOD、POD、CAT活性變化
在培養30 d后,兩種培養基上的逆轉穗芽和非逆轉穗芽SOD活性無顯著差異,但培養60 d和90 d后,逆轉穗芽SOD活性總體高于非逆轉穗芽(表6)。培養30 d后,柳枝稷4-6的逆轉穗芽POD活性顯著高于其他3組處理;培養至60 d時活性有所下降,與柳枝稷14-7逆轉穗芽總體低于非逆轉穗芽,但未達到顯著水平;培養90 d后,柳枝稷14-7的逆轉穗芽POD活性急劇上升,極顯著高于其他3組處理,并且逆轉穗芽POD酶活性總體高于非逆轉穗芽,這表明在繼代2次培養基碳源和氮源缺乏的情況下,POD酶產生響應,以起到保護的作用(表7)。CAT活性在培養30 d后各處理間未表現出顯著差異,但培養60 d后逆轉穗芽酶活性顯著高于非逆轉穗芽,而培養90 d后,柳枝稷14-7的逆轉穗芽CAT活性卻急劇下降,與SOD和POD酶活性變化截然相反,這表明CAT活性并未因碳源和氮源缺乏而產生響應,相反的是其活性受到明顯的抑制(表8)。兩種柳枝稷之間的SOD、POD和CAT差異均極顯著(P=0.0013,P=0.0003,P=0.0011),然而SOD和POD均表現為柳枝稷14-7活性更高,而CAT表現為4-6活性更高,這可能是不同類型抗氧化酶對碳氮缺乏響應的差異所引起;SOD各時期差異不顯著,而誘導90 d的穗芽POD和CAT活性與其他兩個時期差異均達到極顯著水平。
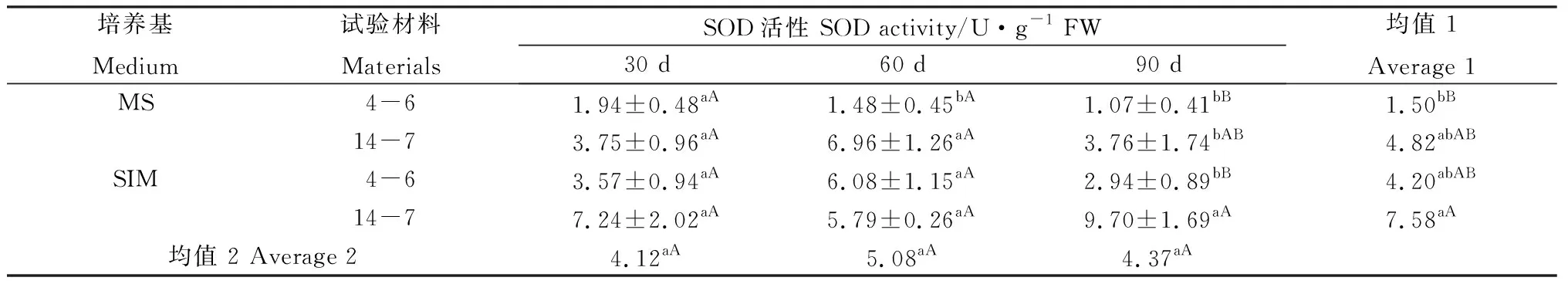
表6 柳枝稷穗芽形成期間SOD活性變化Table 6 SOD activity variation of different switchgrass spike buds

表7 柳枝稷穗芽形成期間POD活性變化Table 7 POD activity variation of different switchgrass spike buds
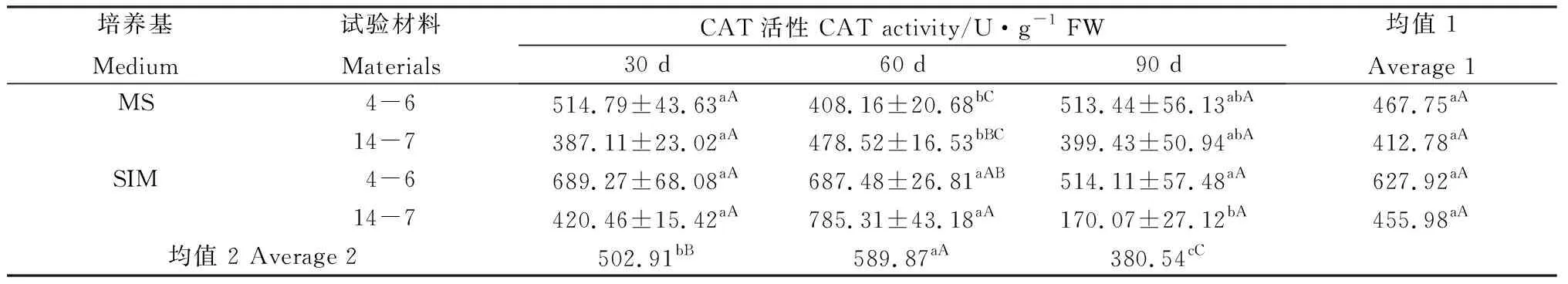
表8 柳枝稷穗芽形成期間CAT活性變化Table 8 CAT activity variation of different switchgrass spike buds
2.6 柳枝稷穗芽發育中PPO活性的變化
方差分析發現,在培養30 d、60 d和90 d時,兩種柳枝稷材料的逆轉穗芽和非逆轉穗芽PPO活性差異均不顯著,但總體上逆轉穗芽酶活性比非逆轉穗芽略高(表9)。兩種柳枝稷之間PPO酶活性無顯著差異(P= 0.7645);各培養時期間相比較發現PPO活性整體呈現先降后升的趨勢。
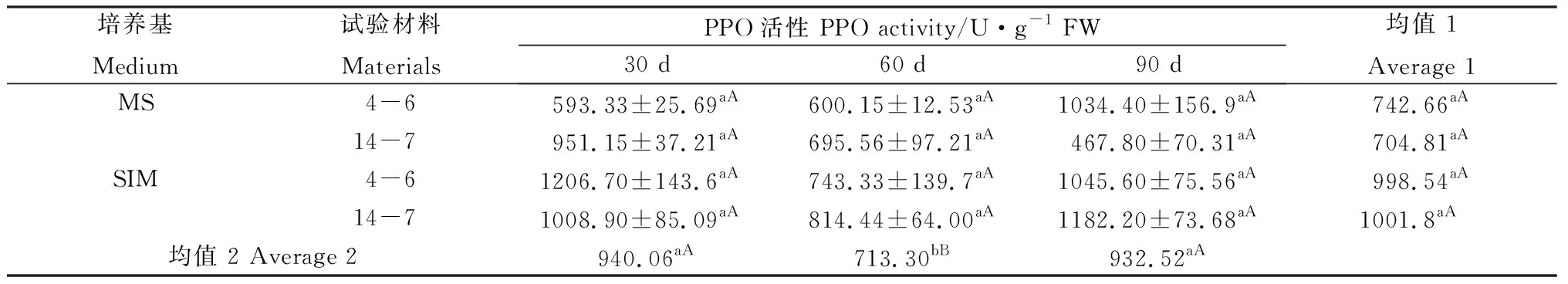
表9 柳枝稷穗芽形成期間PPO活性變化Table 9 PPO activity variation of different switchgrass spike buds
2.7 柳枝稷穗芽發育中ZR、IAA含量的變化
對兩種類型柳枝稷逆轉和非逆轉穗芽的ZR含量分析發現,培養30 d和60 d后,逆轉和非逆轉穗芽間均未體現出顯著的差異;培養90 d后柳枝稷4-6的非逆轉穗芽ZR含量顯著高于其他處理但未達到極顯著水平(表10)。材料培養30 d后,非逆轉穗芽IAA含量顯著高于逆轉穗芽,培養60 d和90 d后穗芽IAA含量雖然也有差異,但兩種柳枝稷卻并未表現出一致的趨勢;此外該結果還表明外源6-BA的施加對IAA的合成體現出一定的抑制作用。通過對IAA/ZR分析,結果與IAA含量類似,整體而言,逆轉穗芽IAA/ZR高于非逆轉穗芽(表12)。兩種柳枝稷ZR含量差異不顯著(P=0.0757),但IAA含量和IAA/ZR差異達到顯著或極顯著水平(P=0.0063,P=0.0148);不同時期相比較發現,ZR含量、IAA含量和IAA/ZR均表現先升后降的趨勢,且時期間差異均達到極顯著水平(P<0.0601)。
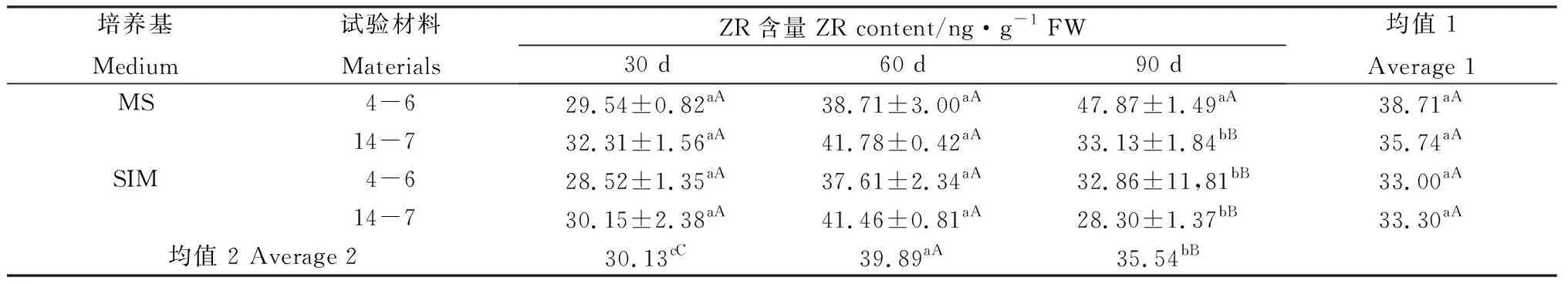
表10 柳枝稷穗芽形成期間ZR含量變化Table 10 ZR content variation of different switchgrass spike buds

表11 柳枝稷穗芽形成期間IAA含量變化Table 11 IAA content variation of different switchgrass spike buds

表12 柳枝稷穗芽形成期間IAA/ZR變化Table 12 The value of IAA/ZR variation of different switchgrass spike buds
2.8 柳枝稷穗芽發育中ABA含量的變化
培養30 d后,柳枝稷4-6的逆轉穗芽ABA含量顯著或極顯著高于其他3組處理,但其他3組處理間差異不顯著;培養至60 d時, 柳枝稷14-7的逆轉穗芽ABA含量有所上升,但4-6的逆轉穗芽ABA含量仍高于其他3組處理,且差異均達到極顯著;培養到90 d時,柳枝稷14-7的逆轉穗芽ABA含量有顯著上升,與4-6的逆轉穗芽ABA含量水平一致,并且均顯著高于MS培養基(表13)。兩種類型柳枝稷所形成的穗芽ABA含量差異極顯著(P= 0.0002),表明兩種柳枝稷的ABA響應存在一定的差異;3個培養時期相比較,差異雖未達到顯著水平,但整體呈上升趨勢,結合逆轉穗芽ABA含量更高的結果可推斷,ABA可能在穗芽發育的后期起到重要的作用。

表13 柳枝稷穗芽形成期間ABA含量變化Table 13 ABA content variation of different switchgrass spike buds
2.9 柳枝稷穗芽發育中GAs含量的變化
當培養至30 d時,兩種類型柳枝稷逆轉穗芽GAs含量均高于非逆轉,差異達到顯著或極顯著水平(表14),表明早期穗芽起始誘導的過程中GAs發揮著重要的作用;培養至60 d和90 d時,兩種類型柳枝稷逆轉穗芽GAs含量仍顯著或極顯著高于非逆轉穗芽,表明在后期逆轉穗芽發育過程中,GAs同樣行使著重要的功能(表14)。就兩種類型柳枝稷而言,14-7的GAs水平整體極顯著高于4-6,結合柳枝稷14-7穗芽生長速度較4-6快,而SIM培養基上的穗芽生長速度較MS培養基快,我們可以推斷GAs對穗芽形成時的快速生長起到重要的作用;3個培養時期比較分析表明,GAs含量隨著培養時間總體呈上升趨勢,并且培養至90 d時其水平極顯著高于前兩個時期,表明GAs在逆轉穗芽形成初期及發育后期均起到重要的作用。
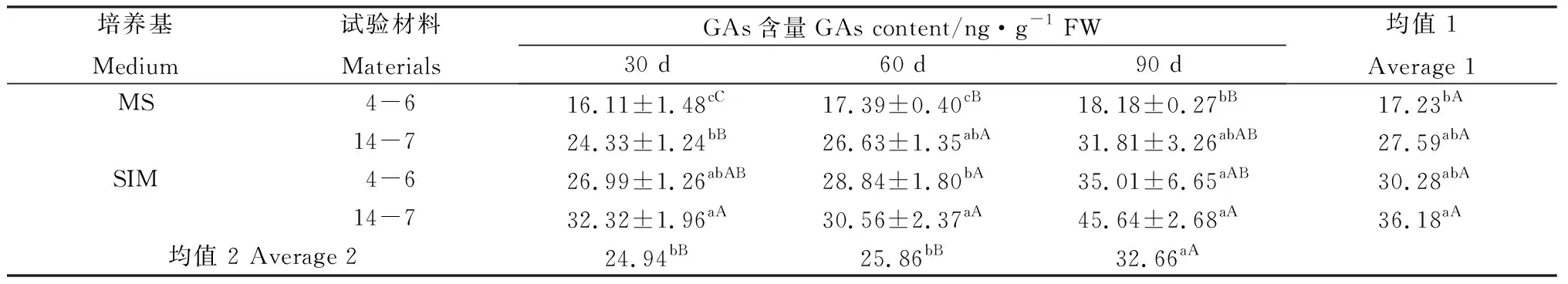
表14 柳枝稷穗芽形成期間GAs含量變化Table 14 GAs content variation of different switchgrass spike buds
3 討論與結論
在花發育的研究進程中,人們往往更關注植物從營養生長向生殖生長的轉變,而對花器官逆向發育的關注較少[26]。本試驗利用體外花器官逆轉誘導方法,通過比較SIM培養基上逆轉穗芽和MS基本培養基上非逆轉穗芽的部分生理指標和激素水平,發現逆轉穗芽具有較低的葉綠素水平,但具有更高的糖含量(前兩個時期30 d、60 d),這可能是因為體外培養過程中的碳源更多來自培養基,而逆轉穗芽葉綠素含量低并非導致其發育速度快的原因。綜合考慮兩種培養基上的穗芽可溶性蛋白、游離氨基酸和總氮含量的變化以及SIM培養基后期氮供應不足可以推斷出,與非逆轉穗芽相比,逆轉穗芽具有更高的氮代謝效率。逆轉穗芽和非逆轉穗芽抗氧化酶活性比較發現,逆轉穗芽整體抗氧化酶系統具有更高的活性。外源施加6-BA對柳枝稷內源ZR的合成無顯著影響,但對IAA的合成可能起到一定的抑制作用,這對穗芽起始誘導起到關鍵的作用。ABA可能在穗芽發育的后期階段起到重要的作用,而GAs可能在穗芽發育的整個時期都有行使重要的功能。
激素是調控植物生長發育的重要物質,在花發育和花器官逆轉過程中也起到至關重要的作用。Gaudinová等在研究病毒引起的黑醋栗(Ribesnigrum)花器官逆轉的現象中發現,被病毒感染的花器官與正常組織相比細胞分裂素含量異常升高[13]。Young等研究表明細胞分裂素在玉米雌蕊發育以及雌蕊發育方向決定中起到重要的作用[27]。這些結果都與本試驗相一致。Jack等在研究中指出赤霉素是花器官形成過程中傳遞光周期信號的重要物質,對花芽的形成起到促進作用,花器官逆轉可能通過降低植物體內赤霉素水平而發生[8]。然而本試驗發現穗芽形成過程中具有較高的赤霉素水平,這可能是因為本試驗中高濃度的外源6-BA主導柳枝稷花器官逆轉的發生,而赤霉素可能起到加快發育進程的作用。此外,Cai等和Yuan等的研究表明,茉莉酸甲酯在花發育中起到促進的作用[11,28]。
在花器官逆轉誘導條件研究中,光周期和溫度被認為是啟始花器官逆轉的重要因素[7,9]。Okamuro等通過研究光周期和激素對擬南芥花器官逆轉的影響試驗發現,LFY和AG在花分生組織保持中起到重要作用,而其突變體發生的花器官逆轉可能與光敏色素和赤霉素信號傳導有關[8]。本試驗進行過程中,外植體培養光周期為20 h白天/4 h黑暗,該條件是不利于短日照植物柳枝稷正常花器官發育的[14]。然而在相同光周期條件的MS基本培養基上無花器官逆轉現象發生,表明在柳枝稷幼穗體外培養過程中,單獨改變光周期無法誘導其花器官逆向發育,外源施加高濃度細胞分裂素是柳枝稷花器官逆轉誘導的必要條件。光周期在誘導柳枝稷花器官逆轉中是否起到一定的作用仍需進一步試驗驗證。
植物花器官逆轉現象能夠使我們從反方向研究花發育,為花發育研究和基因功能驗證提供新的思路。Wang等通過對水稻花器官逆轉現象的研究發現DEP和AFO基因在決定花的形成中起到重要的作用[6]。Ampomah-Dwamena等通過對西紅柿中的SEP同源基因TM29共抑制和RNA干擾研究發現,轉基因植株在一定程度上表現出花器官逆轉的現象,甚至在部分畸形果實上能夠長出新的葉子或芽[29]。本試驗采用體外誘導方法獲得柳枝稷花器官逆轉,通過對逆轉穗芽和非逆轉穗芽的比較研究,在一定程度上揭示了柳枝稷穗芽形成過程中的生理生化特性。同時本試驗存一定的不足之處,為確保結果一致性,所選柳枝稷材料只有栽培種Alamo的兩個單株,試驗結果具有一定的局限性,但二者結果整體表現一致,對研究花發育以及花器官逆轉能夠提供一定的參考作用。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現代語文(2016年21期)2016-05-25 13:13:50
