池蝶蚌β-連環蛋白基因cDNA的克隆及表達特征分析
2017-09-12 01:17:18閆含笑史建伍盛軍慶王軍花洪一江
水生生物學報 2017年5期
閆含笑 史建伍 盛軍慶 王軍花 洪一江
(南昌大學生命科學學院, 南昌 330000)
池蝶蚌β-連環蛋白基因cDNA的克隆及表達特征分析
閆含笑 史建伍 盛軍慶 王軍花 洪一江
(南昌大學生命科學學院, 南昌 330000)
為了解淡水貝類性別調控與分化機制, 課題組建立了池蝶蚌(Hyriopsis schlegelii)性腺轉錄組, 在轉錄組庫中, 存在β-連環蛋白(β-catenin)基因序列。實驗對池蝶蚌β-catenin基因進行驗證, 采用RACE技術克隆其cDNA全長, 命名為Hsβ-catenin。該序列全長4386 bp, 5′-非編碼區為162 bp, 3′-非編碼區為1758 bp, 開放閱讀框為2466 bp, 編碼821個氨基酸; 該蛋白結構域主要由12個ARM重復序列組成; 二級結構中, α-螺旋占47.75%, β-折疊占1.22%, 隨機卷曲占51.04%; 三級結構中含大量α-螺旋且為右手超螺旋, 構成ARM結構域; 系統進化樹分析表明, Hsβ-catenin與軟體動物聚為一支, 然后與昆蟲類聚為一支。實時熒光定量PCR(qRT-PCR)檢測顯示, Hsβ-catenin在腸中表達量最高, 其次是斧足和精巢。Hsβ-catenin基因在12月齡和36月齡表達量較高, 且在36月齡表達量最高, 表明其可能參與池蝶蚌的性別調控與分化作用。
池蝶蚌; Hsβ-catenin; 組織表達; 克隆; 序列分析; qRT-PCR
β-連環蛋白(β-catenin)是連環蛋白家族中的一個蛋白, 于1980年作為一種黏附因子被發現[1]。βcatenin有3個結構域, N末端區包括多個絲氨酸、蘇氨酸殘基作為多種酶的磷酸化位點; 中央區是最保守的區域, 含12—14個ARM結構域(犰狳重復結構域), 每個ARM結構域形成呈三角形排列的3個環形結構, 這些結構域因帶正電荷在超螺旋結構中形成凹槽, 成為Wnt通路中許多信號分子的結合位點; C末端區含有轉錄激活區, 能夠激活下游靶基因的轉錄[2]。β-catenin蛋白廣泛存在于各類細胞中, 主要通過與其他分子形成復合物在不同組織結構中起作用[3]; 在細胞中主要有2個作用, 一是與細胞膜上的鈣黏蛋白相互作用, 參與細胞間黏附, 二是作為Wnt信號通路中的關鍵信號分子調控細胞的生長、分化與凋亡[4]。依賴β-catenin的Wnt通路被稱為經典Wnt通路, 多種Wnt通路上的信號分子與βcatenin結合形成破壞復合物, 細胞外分泌的Wnt配體作用于破壞復合物并使其降解, β-catenin濃度不斷上升, 進而促進Wnt蛋白的表達; 而沒有Wnt配體時, β-catenin不斷被磷酸化, 濃度下降, Wnt通路被阻斷[5]。β-catenin被發現通過Wnt通路對性別調控、癌癥的轉移侵襲、神經系統的發育等途徑起著重要作用。
池蝶蚌(Hyriopsis schlegelii)是優質的淡水珍珠育珠蚌, 在生長過程中存在雌雄同體現象, 但成熟后性別為單性[6]; β-catenin被發現在青鳉中具有誘導卵巢發育甚至使性腺性逆轉的作用[7], 我們在池蝶蚌性腺轉錄組中也發現有β-catenin, 是否同樣對池蝶蚌的性別分化有調控作用?因此本研究對池蝶蚌β-catenin基因進行克隆, 分析其結構特征及在不同組織和不同年齡性腺中的表達量, 以為其功能分析奠定基礎。
1 材料與方法
1.1 實驗材料
體質健壯池蝶蚌由江西省撫州市國家級池蝶蚌良種場提供, 養于實驗室水族箱中, 水溫18—25℃, 每天換已曝氣自來水2次。
1.2 實驗方法
RNA的提取和cDNA的合成 經液氮研磨后池蝶蚌組織利用Trizol試劑盒(Invitrogen, USA)一步法提取總RNA, RNA濃度和純度用核酸測定儀(Nanodrop 2000)測定, 并用DNaseⅠ(TaKaRa, Japan)消化處理總RNA以消除基因組DNA的殘留, 最后采用1%瓊脂糖凝膠電泳檢測其完整性。使用Super ScriptTMⅢ Reverse Transcriptase Kit反轉體系合成cDNA, 備用。
池蝶蚌β-catenin cDNA的克隆 依據本實驗室已有的轉錄組數據設計上、下游引物, 擴增出β-catenin 基因的中間片段。50 μL PCR反應體系為: cDNA模版1 μL, Ex Taq酶0.5 μL, 10×Ex Taq buffer 5 μL, dNTP(2.5 mmol/L)5 μL, 上下游引物各1 μL, ddH2O 36.5 μL。PCR反應條件為: 94℃預變性5min; 94℃ 30s, 59℃ 30s, 72℃ 30s, 35個循環; 72℃延伸10min。設計RACE PCR的5′端引物(GSP1和GSP2)和3′端引物(GSP3和GSP4)(表 1)。用特異性引物和通用引物UPM(Long:Short=1:5)為兩側引物、cDNA為第一輪模版進行PCR擴增; 用特異性引物和通用引物NUP作為兩側引物, 將擴增產物稀釋10—50倍作為模版進行第二輪PCR擴增,獲得PCR產物。25 μLPCR反應體系: cDNA模版1 μL, Ex Taq酶0.3 μL, 10×Ex Taq buffer 2.5 μL, dNTP (2.5 mmol/L)1.2 μL, 上下游引物各1 μL, ddH2O 18 μL。反應條件: 94℃預變性5min; 94℃ 30s, 59℃30s, 72℃ 30s, 35個循環; 72℃延伸10min。將PCR產物經1.0%瓊脂糖凝膠電泳檢驗后, 用瓊脂糖凝膠回收試劑盒(OMEGA公司)回收、純化, 并與pMD19-T載體連接, 轉入感受態細胞E. coli DH5α中, 于LB平板(含Amp+)培養, 陽性克隆經PCR鑒定后送至上海生工生物工程技術服務有限公司測序。獲得的序列經MEGA軟件分析確定為Hsβ-catenin基因。利用ORF finder(https://www.ncbi.nlm.nih.gov/orffinder/)查找此cDNA序列的最大開放閱讀框(ORF), 對翻譯所得氨基酸序列進行一、二、三級結構預測分析, 利用MEGA軟件構建分子系統樹。
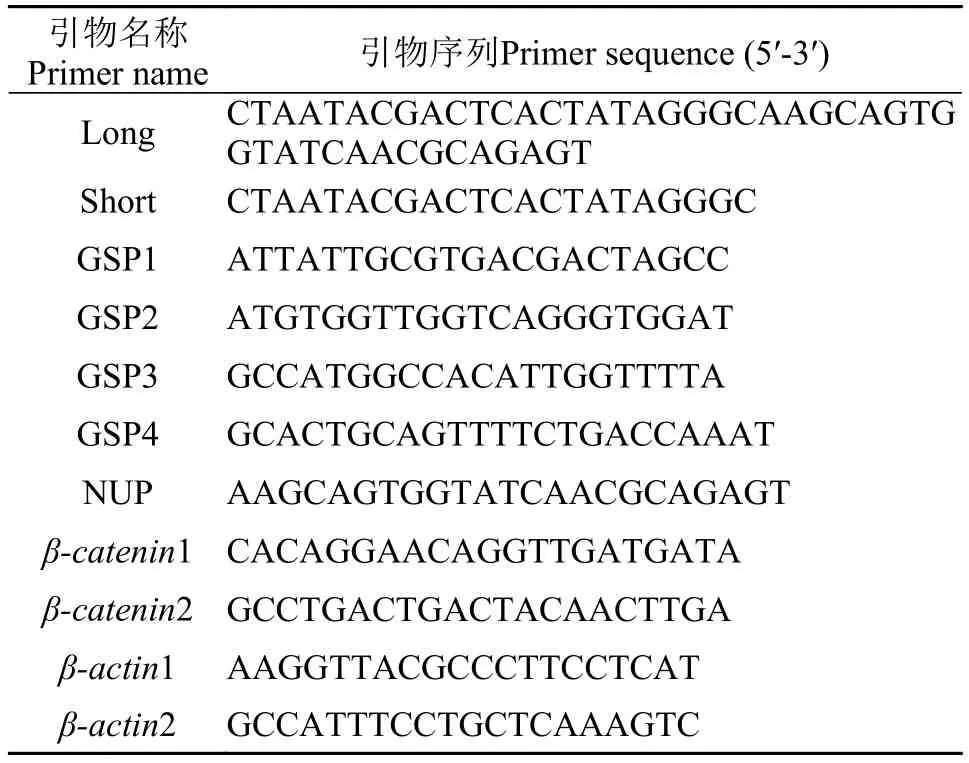
表 1 PCR相關引物序列Tab. 1 Primers used for PCR
qRT-PCR分析 采用qRT-PCR技術檢測Hsβ-catenin基因的組織轉錄差異性及不同年齡、不同性別中性腺組織的表達差異性。根據得到的Hsβ-catenin cDNA全長序列, 設計熒光定量PCR引物(表 1)進行qRT-PCR。對于組織表達實驗, 隨機選取6只成熟池蝶蚌, 雌、雄各3只采取相同組織混合法分別提取池蝶蚌卵巢、精巢、心臟、外套膜、腸、腎、肝、血和斧足各組織總RNA, 通過1%瓊脂糖凝膠電泳檢驗RNA完整性。將RNA反轉為cDNA模板; 不同年齡、不同性別池蝶蚌Hsβcatenin表達實驗方案: 選取1—4齡雌、雄性池蝶蚌各3只, 取各年齡雌、雄性腺, 按上述方法獲得cDNA模板。根據熒光定量試劑盒(嵌合檢測法, Takara)進行qRT-PCR檢測。20 μL PCR反應體系為: SYBR10 μL, 上、下游引物各0.4 μL, H2O 8.2 μL。每個樣品設置3個平行, 以蒸餾水代替模板作為陰性對照。反應條件為: 95℃預變性5min; 94℃ 30s, 58℃ 30s, 72℃ 30s, 40個循環; 72℃延伸10min。以上實驗均重復3次, SPSS軟件分析Hsβ-catenin mRNA在池蝶蚌中的表達情況, 組織表達和不同年齡性腺表達均進行單因素方差統計分析。
2 結果
2.1 池蝶蚌Hsβ-catenin 基因cDNA全長及其分析
池蝶蚌Hsβ-catenin 基因cDNA全長 將測序結果用MEGA軟件處理后拼接成完整的cDNA序列, 全長4386 bp(登錄號為KY126093), 其中5′端非編碼區162 bp, 3′端非編碼區1758 bp, ORF框2466 bp, 共編碼821個氨基酸。將拼接后的cDNA序列與實驗室已有的轉錄組序列進行比對, 二者基本相同, 證明了實驗結果的準確性。
Hsβ-catenin蛋白結構域分析 通過瑞士生物信息學研究所ExPASy服務器的SMART程序(http://smart.embl-heidelberg.de/)分析池蝶蚌Hsβcatenin蛋白結構域, 可知池蝶蚌Hsβ-catenin蛋白主要由12個ARM結構域組成。ARM結構是各物種中β-catenin蛋白的中央保守區, 從而驗證了實驗結果的準確性。
Hsβ-catenin二級結構預測 通過PredicProtein在線軟件(http://www.predictprotein.org/)對池蝶蚌Hsβ-catenin的二級結構進行預測。預測可知二級結構中α-螺旋占47.75%, 分布比較均勻且大量分布; β-折疊占1.22%, 在各處均有分布, 總量較少; 隨機卷曲占51.04%, 在兩端分布較多。根據蛋白表面暴露面積是否超過16%為標準對蛋白進行親水性和疏水性預測, 親水性蛋白含量約為49.09%, 疏水性蛋白含量約為50.91%。
Hsβ-catenin三級結構預測 通過瑞士生物信息所ExPASy服務器的SWISS-MODEL程序(http://swissmodel.expasy.org/)對池蝶蚌Hsβ-catenin的三級結構進行預測(圖 1)。圖中含大量α-螺旋且為右手螺旋, 每3個α-螺旋排列在一起形成1個超螺旋, 即ARM結構域, 中間區域共形成12個ARM重復區域; 圖中還包括少量β-折疊及大量無規則卷曲。三級結構預測結果與二級結構一致。
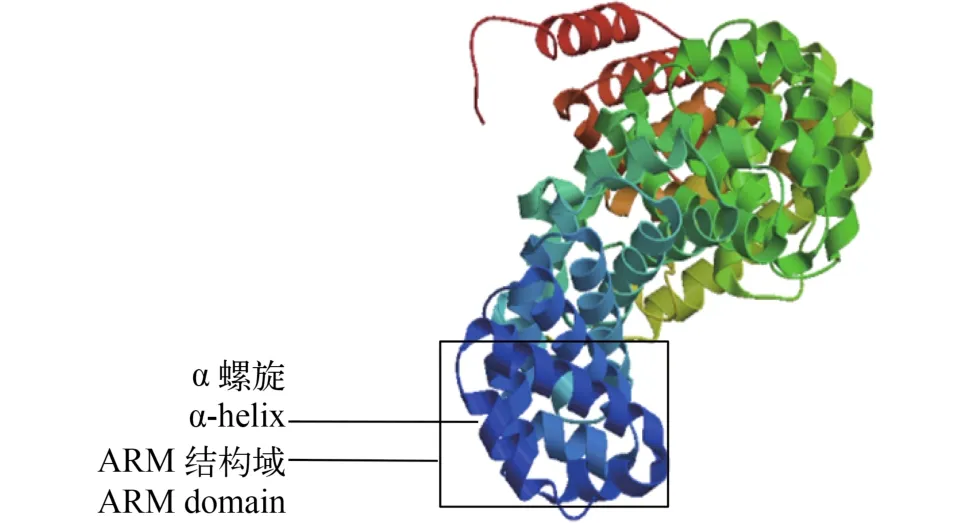
圖 1 Hsβ-catenin的三級結構預測Fig. 1 The tertiary structure prediction of Hsβ-catenin
2.2 Hsβ-catenin基因的系統發育分析
在NCBI(http://www.ncbi.nlm.nih.gov/gorf/ orfig.cgi)上查找各物種β-catenin 基因的cDNA序列,利用MEGA軟件分析可得到Hsβ-catenin基因的分子進化樹(圖 2), 由圖可知, 該基因序列比較保守,同源性較高。池蝶蚌Hsβ-catenin 基因與軟體動物的同源性很高, 構成一個分支, 其中與合浦珠母貝(Pinctada martensii)、櫛孔扇貝(Azumapecten farreri)和太平洋牡蠣(Crassostrea gigas)同源性最高,雜斑蓮花青螺(Lottia luchuana)、海蝸牛(Aplysia californica)和大西洋舟螺(Crepidula fornicata Linnaeus)次之; 與昆蟲類的同源性也很高, 如毛翼蟲(Chaetopterus variopedatus)、杜氏闊沙蠶(Platynereis dumerilii)和大斑蝶(Danaus plexippus); 與人類和魚類的同源性相對較低。
2.3 池蝶蚌Hsβ-catenin組織表達分析
用熒光定量PCR技術檢測池蝶蚌Hsβ-catenin在成熟個體9個組織中的表達(圖 3)。由圖可知, Hsβ-catenin 在各組織中均有表達, 表達量從高到低為: 腸、斧足、精巢、肝、卵巢、心臟、外套膜、腎和血, 相對Hsβ-actin表達量分別為: 8.44、4.43、3.05、2.22、1.84、1.00、0.92、0.64和0.06, 表明其在生物體各組織中具有多種生物學功能, 且在成熟池蝶蚌中精巢表達量高于卵巢, 推測Hsβ-catenin在性腺發育成熟后更多地參與其他生物學功能。

圖 2 不同物種β-catenin基因分子進化樹Fig. 2 The phylogenetic tree of β-catenin合浦珠母貝Pinctada martensii (AKJ32471.1);櫛孔扇貝Azumapecten Farreri (AFU35436.1);太平洋牡蠣Crassostrea gigas (NP_001292224.1);池蝶蚌 Aplysia californica (NP_00119 1600.1);雜斑蓮花青螺Lottia luchuana (XP_009048556.1);大西洋舟螺Crepidula fornicata Linnaeus (ADI48180.1);白氏文昌魚Branchiostoma belcheri (BAD12593.1);意蜂Apis mellifera (NP_001172034.1);內華達古白蟻Zootermopsis nevadensis (KDR13423.1);雙斑蟋Gryllus bimaculatus (BAD00045.1);大斑蝶Danaus plexippus (EHJ72882.1);嗜鳳梨果蠅Drosophila ananassae (XP_001966028.1);人類Homo sapiens (CAA61107.1)
2.4 池蝶蚌Hsβ-catenin在不同年齡性腺中的表達分析
池蝶蚌在5月齡出現性腺, 12月齡雌雄可辨, 36月齡性腺成熟。檢測Hsβ-catenin在不同年齡、不同性腺中的表達情況(圖 4), 結果顯示其在3齡(約36月齡)中的表達最高, 1齡(約12月齡)表達量次之, 2齡(約24月齡)和4齡(約48月齡)表達量最少; 在1到3齡中卵巢的表達量顯著高于精巢的表達量, 而到4齡時精巢反而略高于卵巢。
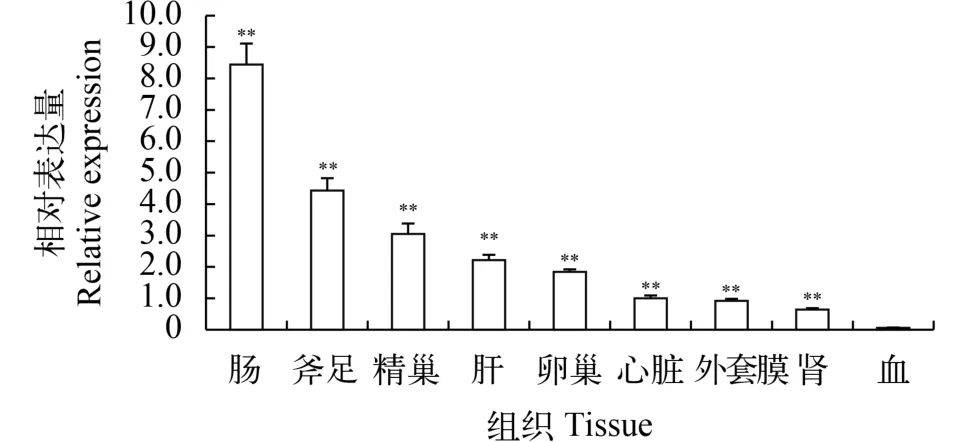
圖 3 Hsβ-catenin在各組織中的表達量
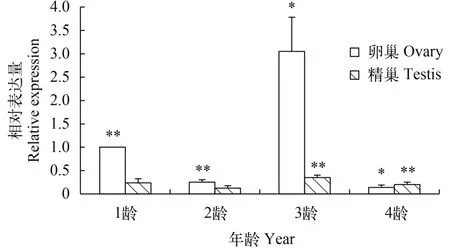
圖 4 Hsβ-catenin在不同年齡、不同性腺中的表達量
3 討論
本研究首次獲得池蝶蚌Hsβ-catenin基因的cDNA全長, 并對其結構特征和蛋白功能進行預測分析。由池蝶蚌Hsβ-catenin蛋白的結構域預測可知, 其主要由12個ARM結構域組成, 序列高度保守,有典型的N末端區和C末端區, 與各種貝類甚至其他物種同源性較高, 預測結果與ARM重復序列的性質相同。ARM重復序列是約有40個氨基酸的串聯重復序列, 通過在果蠅中起信號傳導的作用首次被發現, 動物的ARM蛋白在細胞內信號轉導和細胞骨架的調節等多種生命活動中起作用, 并參與βcatenin、腺瘤樣息肉(APC)的腫瘤抑制蛋白和核運輸因子importin α等蛋白的構建[8]。ARM結構高度保守, 僅有右手超螺旋結構, 普遍存在于真核細胞中, 如在擬南芥中起著細胞信號傳導功能的ARM重復序列對生長發育和繁殖起著重要作用[9]。β-catenin包含3個區域, 各司其職, 第一個區域為N末端區, 含有GSK-β一致性的磷酸化位點; 第二個區域為中央區, 含有ARM重復序列, 能夠與鈣黏素、APC (Adenomatous polyposis coli)和TCF/LEF (T-cell Factor/Lymphoid Enhancing Factor)相互作用;第三個區域為C末端區, 含有轉錄激活區, 對靶基因激活是所需的[10]。我們推測在經典Wnt通路中, 池蝶蚌β-catenin蛋白ARM結構域中央區可以與APC和其他信號分子結合形成破壞復合物, N末端區的磷酸化位點可與GSK-3β作用, 使破壞復合物中的β-catenin不斷被磷酸化, 無法進入細胞核中起作用; 當Wnt蛋白分泌時, GSK-3β不能磷酸化βcatenin, 從而抑制降解復合物的形成, 導致胞漿中的β-catenin不被降解并進入細胞核, 中央區與核內轉錄因子LEF/TCF家族成員結合, 而C末端的轉錄激活區促進下游靶基因表達。
在非洲爪蟾胚胎中, β-catenin出現在所有細胞中, 參與細胞間黏附功能, 特別在形態發生活躍的區域如囊胚和原腸胚的邊緣區有β-catenin高水平表達; 在原基細胞基團之間也發現β-catenin高水平表達, 說明其參與生物發育過程[11]。β-catenin可以與FHL2相互作用促進小鼠肌細胞的分化[12], Wnt經典通路的激活可以誘導小鼠腎單位的分化和腎的形成[13]。在人類[14]、果蠅[15]、非洲爪蟾[11]、羅非魚[16]等物種中也都具有的廣泛表達的特點。本研究表明, 池蝶蚌β-catenin在腸、斧足、精巢、肝、卵巢、心臟、外套膜、腎和血9個組織中均有表達,而不是僅作為性別決定和性別調控基因大量出現在精巢和卵巢中, 表明其在生物體中廣泛存在且可在不同組織中參與各種生理功能。根據在外套膜、斧足、腎及各細胞中的表達情況及其堿基序列的高度保守性, 我們推測, 其可能參與池蝶蚌肌細胞的分化與腎的形成, 并參與各組織內細胞間黏連等各種生理活動, 這與其他物種有功能類似; 但另一方面Hsβ-catenin在池蝶蚌各組織的表達量相對較低, 推測是由于Wnt信號通路在各組織中廣泛存在。
β-catenin是一種親卵巢、抗精巢的信號分子,對性別決定機制起著一定的作用并可導致性逆轉。為了研究β-catenin是否可以阻斷睪丸的發育通路, 在小鼠XY型性細胞中激活β-catenin的表達, 結果顯示XY型小鼠雄性發育通路被阻斷, 并造成雄性到雌性的性逆轉[17]。目前一些β-catenin對性腺決定和分化作用的研究認為, 哺乳動物尚未分化的性腺的發育方向是由R-spondin 1、Wnt 4和β-catenin等信號分子介導的雌性信號通路和由Sry和Sox9等信號分子介導的雄性信號通路相互競爭決定的,性腺最終朝著競爭占優勢的一方發育[16]。我們對不同性別和不同年齡池蝶蚌中Hsβ-catenin基因的表達量也進行了分析, 結果發現, 12月齡雌雄可辨時卵巢中表達量較高, 36月齡的成熟卵巢中表達量最高。由此推測, Hsβ-catenin可能在池蝶蚌雌性性別決定與分化過程中起著調控作用。另外一個有趣現象是第一次性成熟后的卵巢完成了性腺分化, Hsβ-catenin表達量反而略低于精巢, 因此我們推測其可能不再行使性腺分化調控功能。
[1]Muercheer, Wen B. New progresses of β-catenin in colon cancer [J]. Journal of North Sichuan Medical College, 2011, 26(6): 554—558 [木爾扯爾, 文彬. 結腸癌中βcatenin研究的新進展. 川北醫學院學報, 2011, 26(6): 554—558]
[2]Tan Y, Jiang N, Sun X. The research progress of β-catenin in ovarian cancer and chemoresistance [J]. Maternal and Child Health Care of China, 2015, 30(21):3746—3749 [譚影, 姜寧, 孫信. β-catenin在卵巢癌及化療耐藥中的研究進展. 中國婦幼保健, 2015, 30(21): 3746—3749]
[3]Li H L. Cloning of genes Wnt4, β-catenin, Dax1, and their correlation to the gonadal development in scallop Chlamys farreri [D]. Ocean University of China. 2013 [李海龍. 櫛孔扇貝(Chlamys farreri)Wnt4, β-catenin及Dax1基因的克隆及其與性腺發育相關性的研究. 中國海洋大學. 2013]
[4]He H. The research progress of β-catenin [J]. Practical Clinical Medicine, 2010, 11(10): 120—123 [何歡. βcatenin的研究進展. 實用臨床醫學, 2010, 11(10): 120—123]
[5]Azzolin L, Panciera T, Soligo S, et al. YAP/TAZ incorporation in the β-Catenin destruction complex orchestrates the Wnt response [J]. Cell, 2014, 158(1): 157—170
[6]Wang B H. Study on development of the gonad of Hyriopsis schlegelii [D]. Nanchang University. 2010 [王濱花. 池蝶蚌性腺發育的觀察. 南昌大學. 2010]
[7]Yu X G, Wu L M, Liu G, et al. Function in the ovary differentiation of R-spondins gene from teleost fish [C]. The 2012 Symposium of Chinese Society for Oceanology and Limnology and China Ichthyology Society. 2012 [于祥國, 吳利敏, 劉剛, 等. R-spondins基因在硬骨魚類卵巢分化中的作用研究. 中國海洋湖沼學會、中國動物學會魚類學分會2012年學術研討會. 2012]
[8]Hatzfeld M. The armadillo family of structural proteins [J]. International Review of Cytology, 1999, (186): 179—224
[9]Coates J. Armadillo repeat proteins: beyond the animal kingdom [J]. Trends in Cell Biology, 2003, 13(9): 463—471
[10]Akiyama T. Wnt/beta-catenin signaling [J]. Cytokine & Growth Factor Reviews, 2000, 11(4): 273—282
[11]Fagotto F, Gumbiner B M. Beta-catenin localization during Xenopus embryogenesis: accumulation at tissue and somite boundaries [J]. Development, 1994, 120(12): 3667—3679
[12]Martin B, Schneider R, Janetzky S, et al. The LIM-only protein FHL2 interacts with β-catenin and promotes differentiation of mouse myoblasts [J]. Journal of Cell Biology, 2002, 159(1): 113—122
[13]Kuure S, Popsueva A, Jakobson M, et al. Glycogen synthase kinase-3 inactivation and stabilization of beta-catenin induce nephron differentiation in isolated mouse and rat kidney mesenchymes [J]. Journal of the American Society of Nephrology, 2007, 18(4): 1130—1139
[14]Riggleman B, Wieschaus E, Schedl P. Molecular analysis of the armadillo locus: uniformly distributed transcripts and a protein with novel internal repeats are associated with a Drosophila segment polarity gene [J]. Genes & Development, 1989, 3(1): 96—113
[15]Helmbrecht K, Kispert A, Von W R, et al. Identification of a Wnt/beta-catenin signaling pathway in human thyroid cells [J]. Endocrinology, 2001, 142(12): 5261—5266
[16]Wu F R. Moleeular cloning, expression and characterization of several genes (Rspo1, β-catenin and TSP-1)involved in ovarian differentiation and development in Tilapia [D]. Southwest University. 2010 [吳風瑞. 羅非魚卵巢分化和發育相關基因Rspo1、β-catenin和TSP-1的克隆、表達和功能鑒定. 西南大學. 2010]
[17]Maatouk D M, Dinapoli L, Alvers A, et al. Stabilization of β-catenin in XY gonads causes male-to-female sex-reversal [J]. Human Molecular Genetics, 2008, 17(19): 2949—2955
THE STRUCTURAL FEATURE AND EXPRESSION ANALISIS OF HSβ-CATENIN cDNA AND PROTEIN FROM HYRIOPSIS SCHLEGELII
YAN Han-Xiao, SHI Jian-Wu, SHENG Jun-Qing, WANG Jun-Hua and Hong Yi-Jiang
(The College of Life Science, Nanchang University, Nanchang 330000, China)
In order to explore the mechanism of sexual development of freshwater shellfish, we established a gonadal transcriptome of Hyriopsis schlegelii. Based on the transcriptome library, this study investigated the β-catenin. The fulllength cDNA was cloned by RACE PCR, which named Hsβ-catenin. The full length of Hsβ-catenin cDNA was 4386 bp that contained an open reading frame (ORF) of 2466 bp coding a protein of 821 amino acids. The protein was mainly composed of 12 ARM repeat sequences. In the secondary structure, the α-helix accounted for 47.75% and the β-folding accounted for 1.22% were predicted. The tertiary structure had many α-helixs, namely ARM domain. Hsβ-catenin gene was clustered with homologous genes from soft animal in the phylogenetic tree. Hsβ-catenin expressed highly in intestine and modestlyin foot and testis. The expression level of Hsβ-catenin was high at the age of 12-month-old (gender differentiation) and 36-month-old (gonad mature), indicated that it might be involved in sexual development.
Hyriopsis schlegelii; Hsβ-catenin; Tissue expression; Clone; Sequence analysis; qRT-PCR
Q344+1
A
1000-3207(2017)05-0972-05
10.7541/2017.121
2016-10-19;
2017-01-14
國家自然科學基金(31660337); 江西省教育廳項目(GJJ150166)資助 [Supported by the National Natural Science Foundation of China (31660337); the Fund Project of Jiangxi Provincial Education Department (GJJ150166)]
閆含笑(1993—), 女, 山東臨沂人; 碩士研究生; 主要研究方向為水生生物學。E-mail: yanhanxiao1993@163.com
洪一江(1963—), 教授; 主要研究方向為水產動物細胞與遺傳育種。E-mail: yjhong2008@163.com
