鹽堿脅迫對三個品種桑樹種子萌發的影響
2017-07-29 09:58:32趙東曉董亞茹杜建勛陳傳杰王照紅
山東農業科學 2017年7期
趙東曉+董亞茹+杜建勛+陳傳杰+王照紅+孫景詩+梁明芝
摘要:鹽堿脅迫是植物遭受的主要非生物脅迫之一。本研究將兩種中性鹽(NaCl和Na2SO4)、兩種堿性鹽(NaHCO3和Na2CO3)按1∶1混合,分別設置5個不同鹽堿濃度,對3個品種桑樹種子進行處理,測定發芽率、發芽勢、相對發芽率、相對發芽勢、發芽指數、相對鹽害率和子葉生長率等參數。結果表明:堿性鹽脅迫對桑樹種子萌發和子葉生長的抑制作用比中性鹽脅迫更強。低濃度的中性鹽溶液(﹤50 mmol/L)可促進桂優12號種子萌發,當鹽溶液濃度高于50 mmol/L時則變為抑制。綜合各項指標分析表明,中性鹽脅迫下桂優12號抗性最強,桂優62號次之,8036×農14最差;堿性鹽脅迫下桂優62號抗性最強,桂優12號次之,8036×農14最差。
關鍵詞:桑樹;品種;鹽堿脅迫;種子萌發
中圖分類號:S888.2 文獻標識號:A 文章編號:1001-4942(2017)07-0049-07
Abstract Saline alkali stress is one of the major abiotic stress in plants. In this study, two kinds of neutral salts (NaCl and Na2SO4) and two kinds of alkaline salts (NaHCO3 and Na2CO3) were mixed at 1∶1, and five different saline concentrations were set to dispose of three varieties of mulberry seeds. The germination rate, germination potential, relative germination rate, relative germination potential, germination index, relative salt damage index and cotyledon growth rate were measured. The results showed that alkaline salt stress had stronger inhibitory effect on seed germination and cotyledon growth than that of neutral salt stress. The low concentration of neutral salt solution (less than 50 mmol/L) could promote the germination of Guiyou 12, but when the concentration of salt solution was higher than 50 mmol/L, it became an inhibitory effect. Under the neutral salt stress, Guiyou 12 had the highest tolerance, followed by Guiyou 62, and 8036× Nong 14 was the worst; under the alkaline salt stress, Guiyou 62 had the highest tolerance, followed by Guiyou 12, and 8036× Nong 14 was the worst.
Keywords Mulberry; Variety; Saline alkali stress; Seed germination
土壤鹽堿化是世界農業生產中主要的非生物脅迫之一[1]。我國鹽堿地約占耕地面積的10%左右。鹽堿地生物多樣性較差、生態系統較脆弱,嚴重制約著農業生產發展[2]。我國的鹽堿地,尤其是內陸鹽堿地多為鹽土和堿土的混合,成分復雜且程度不同,使得人們很容易將鹽地和堿地混為一談,籠統地稱為鹽堿地[3]。實際上鹽脅迫和堿脅迫是兩種不同的非生物脅迫[4]。土壤鹽化主要由NaCl、Na2SO4等中性鹽造成,而土壤堿化則主要由Na2CO3、NaHCO3等堿性鹽所造成,并且由于堿脅迫下植物除了要承受與鹽脅迫相同的滲透脅迫和離子傷害以外,還要抵御高pH值的脅迫,因此土壤堿化對植物的傷害比鹽化更嚴重[5]。
桑樹(Morus alba)是家蠶的唯一飼料樹種,是傳統蠶桑產業的重要物質基礎,也是用于沙漠化及鹽堿化土地治理極具潛力的生態樹種之一[6]。目前我國蠶桑產業正從東南部地區向西北干旱、鹽堿化地區發展轉移,篩選和培育耐鹽堿桑樹品種是實現其大面積推廣應用的前提[7]。桑樹抗性育種中最常見且有效的方法是通過篩選抗逆性種質資源作為育種基礎材料,培育高抗性品種。種子萌發是植物最關鍵的一個生長階段,也是植物生活史中抗逆性最薄弱的一個環節[8]。種子的耐鹽堿性是植物耐鹽堿性早期鑒定和耐鹽堿品種選育的重要指標。鹽堿脅迫會影響植物種子的萌發。通過測定經一定濃度鹽堿溶液處理后種子的發芽率、發芽勢、發芽指數和活力指數等性狀指標評價植物的耐鹽堿性是最為簡單有效的方法。目前已有玉米[9]、豇豆[10]、水稻[11]、大豆[12]、茶花鳳仙[13]、水榆花楸[14]等植物采用該方法評價其耐鹽堿性。本研究選用不同濃度的混合中性鹽(NaCl和Na2SO4)和混合堿性鹽(Na2CO3和NaHCO3)處理3個不同品種桑種子,調查其在不同濃度中性鹽和堿性鹽脅迫下的萌發情況,綜合評價其耐鹽堿性能,以期為摸清桑種對鹽堿脅迫的響應機制和耐鹽堿性桑品種的篩選提供理論基礎。
1 材料與方法
1.1 供試材料
試驗于2016年在山東煙臺進行。供試雜交桑組合8036×農14由山東省蠶業研究所選育,桂優 12號和桂優 62 號由廣西壯族自治區蠶業技術推廣總站提供。
1.2 方法
1.2.1 種子萌發鹽堿脅迫處理方法 用1/4 Hoagland營養液配制的中性鹽和堿性鹽溶液處理桑種子,混合中性鹽(NaCl∶Na2SO4=1∶1)、混合堿性鹽(NaHCO3∶Na2CO3=1∶1)溶液終濃度均分別為50、100、150、200 mmol/L,以1/4 Hoagland營養液為對照(CK)。以上溶液用高壓蒸汽滅菌(121℃,20 min)。隨機抽取各供試桑品種的種子,室溫下經雙蒸水浸泡24 h 后,轉入 75% 乙醇中浸泡 20 s,蒸餾水沖洗 3次。無菌培養皿中放入 3 層濾紙,加入上述各種濃度溶液至濾紙飽和并稍有余液滲出。每個平皿內分散放入60粒桑種子,種粒之間保持一定距離。每個處理組3個重復,標記并記錄各平皿質量。之后每日補水至首次記錄的質量,以保持各平皿中鹽濃度不變。平皿置于人工智能培養箱中培養,培養條件:25℃、10 h光照,20℃、14 h黑暗;相對濕度75%;光照強度≥3 000 lx[15]。
1.2.2 種子萌發調查與測定方法 以胚根長度超過種子長度的1/2作為發芽標準。播種次日起每天調查統計,計算種子發芽率、發芽勢、發芽指數、鹽害指數,發芽后測萌芽種子的子葉長出情況,并對不同品種進行耐鹽性分析[16,17]。
各項性狀指標的計算公式如下:
發芽勢(%)=4 d正常發芽種子數/供試種子數× 100;
發芽率(%)=20 d正常發芽種子數/供試種子數×100;
發芽指數(Gi) =∑(Gt/Dt),式中 Gt為發芽種子粒數,Dt為相應的發芽日數;
相對發芽勢(%)=處理組種子發芽勢/對照組種子發芽勢× 100;
相對發芽率 (%)=處理組種子發芽率/對照組種子發芽率×100;
子葉生長率(%)=20 d子葉長出數/供試種子數× 100;
相對鹽害率(%)=(對照發芽率-鹽處理發芽率)/對照發芽率×100。
2 結果與分析
2.1 不同濃度鹽堿脅迫對3個品種桑樹種子萌發性狀的影響
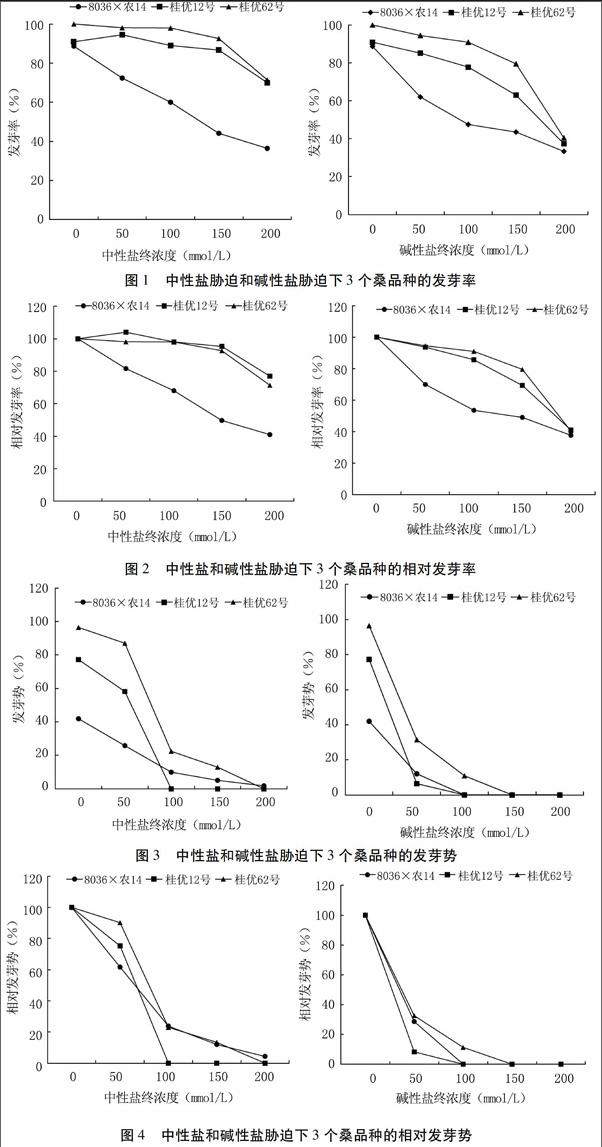
由圖1可以看出,不同濃度鹽堿脅迫對不同桑品種種子萌發的影響不同。中性鹽脅迫下,隨著鹽濃度增加,8036×農14和桂優62號的發芽率有不同程度下降,而桂優12號的發芽率則呈現先升高后降低的趨勢。8036×農14的發芽率隨著中性鹽濃度的增加而顯著降低。桂優62號的發芽率在中性鹽濃度≤150 mmol/L時下降并不明顯,在200 mmol/L時才有明顯下降,比對照低23%。桂優12號的發芽率在中性鹽濃度50 mmol/L時小幅升高,在100~150 mmol/L時下降并不明顯,在200 mmol/L時明顯下降,發芽率比對照低28.6%。
在堿性鹽脅迫下,3個桑品種的發芽率均不斷下降。8036×農14在堿性鹽終濃度為50 mmol/L時就有顯著降低,而桂優12號和桂優62號在堿性鹽終濃度≤150 mmol/L時下降較緩,在200 mmol/L時明顯下降。
由圖2可知,中性鹽處理下,桂優12號的相對發芽率在50 mmol/L時有一個小幅增高,在終濃度≥100 mmol/L時桂優12號和桂優62號的相對發芽率相差不大,在終濃度≤150 mmol/L時,桂優12號和桂優62號的相對發芽率均達到90%以上,在終濃度為200 mmol/L時相對發芽率均在70%以上,推測這兩個品種對中性鹽脅迫不敏感。而8036×農14的相對發芽率則隨鹽濃度增加而顯著下降,因此推斷8036×農14對中性鹽脅迫耐受性較差。
在堿性鹽處理下,桂優62號的相對發芽率在終濃度為100~150 mmol/L高于桂優12號,其它幾個濃度下二者基本相同,且在終濃度200 mmol/L時兩者均有大幅度下降。8036×農14的相對發芽率則在堿性鹽終濃度≥50 mmol/L時就有顯著下降,且隨鹽濃度升高逐漸下降。由此可見桂優12號和桂優62號對低濃度堿性鹽有一定的耐受性,8036×農14則對堿性鹽脅迫較敏感。
發芽勢反映種子發芽的快慢和整齊度,是鑒別種子發芽整齊度的主要指標,在發芽率相同時,發芽勢高的種子生命力強[18]。由圖3可看出,8036×農14發芽勢較低,隨著中性鹽濃度增加逐漸下降。桂優12號和桂優62號發芽勢在中性鹽濃度為50 mmol/L時分別下降24.7%和9.8%,但是在中性鹽濃度≥100 mmol/L時發芽勢銳減,說明高濃度的中性鹽抑制了種子的萌發。
相對于中性鹽,堿性鹽加劇了對桑樹種子萌發的抑制。在堿性鹽終濃度為50 mmol/L時3個桑品種的發芽勢就有顯著下降,隨鹽濃度增加不斷下降;在終濃度為100 mmol/L時,桂優12號和8036×農14的發芽勢為零,桂優62號的發芽勢在堿性鹽終濃度為150 mmol/L時為零,即無發芽種子。
3個桑品種種子在遭受中性鹽脅迫時相對發芽勢均有不同程度降低(圖4),其中桂優12號下降最快,桂優62號下降最慢。堿性鹽脅迫下,相對發芽勢下降趨勢加快,在堿性鹽濃度≤150 mmol/L時桂優62號的相對發芽勢高于桂優12號和8036×農14。推測桂優62號耐堿性鹽能力最強。
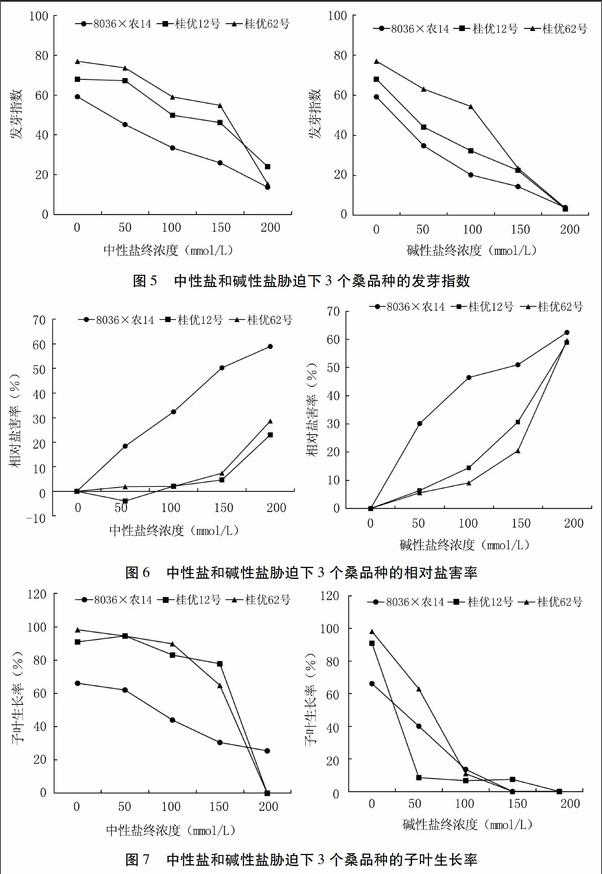
發芽指數是衡量種子在鹽堿脅迫下不同天數發芽情況的重要指標,發芽指數能很好地反映種子萌發的速度和整齊程度[19],可以反映植物芽期耐鹽性的強弱,發芽指數越大,表明植物的耐鹽性越強[20]。由圖5可知,3個桑品種的發芽指數與鹽堿脅迫呈負相關。中性鹽處理下,濃度≤150 mmol/L時發芽指數由高到低依次為桂優62號、桂優12號、8036×農14,當濃度≥200 mmol/L時發芽指數由高到低依次為桂優12號、桂優62號、8036×農14。堿性鹽處理下,濃度≤150 mmol/L時發芽指數由高到低依次為桂優62號、桂優12號、8036×農14,當濃度≥200 mmol/L時發芽指數均為零。
2.2 不同濃度鹽堿脅迫下3個桑樹品種相對鹽害率的分析
由圖6可知,50 mmol/L中性鹽有利于桂優12號的萌發。在中性鹽脅迫下,鹽害率最高的是8036×農14,最低的是桂優12號。在堿性鹽脅迫下,鹽害率最高的是8036×農14,最低的是桂優62號。從這一結果看,桂優12號耐中性鹽脅迫能力最強,桂優62號次之,8036×農14最差。而堿性鹽脅迫抗性最強的是桂優62號,桂優12號次之,8036×農14最差。
2.3 不同濃度鹽堿脅迫對3個桑樹品種子葉生長率的影響
鹽堿脅迫不僅抑制種子的萌發,同樣也抑制子葉生長。播種20 d后子葉生長情況如圖7所示。中性鹽脅迫下,除桂優12號在50 mmol/L時子葉生長率稍有增加外,3個品種的子葉生長率均呈下降趨勢。8036×農14的子葉生長率一直最低,這可能與種子活力不高有關。在中性鹽濃度為200 mmol/L時,桂優12號和桂優62號的子葉生長率銳減至零,但8036×農14卻仍有25.4%子葉生長率。
桂優12號子葉生長率在堿性鹽脅迫下下降很快,50 mmol/L時即降為較低水平。桂優62號降低最慢。≥100 mmol/L時,3個品種的子葉生長率均較低。由此可見堿性鹽脅迫對子葉生長的抑制性比中性鹽脅迫更強。
3 討論與結論
通過調查3個不同品種桑樹種子在不同濃度鹽堿脅迫下的萌發狀況,發現鹽堿脅迫對桑種子萌發有明顯影響,并且隨著鹽分脅迫濃度的增加,桑種子的發芽勢、發芽率、發芽指數、子葉生長率等均呈明顯下降趨勢,不同品種間萌發指標下降程度不同。
種子萌發是植物生命活動的起始階段,也是植物對外界逆境脅迫最敏感的時期。本試驗結果顯示,低濃度的中性鹽脅迫對桂優12號種子的萌發有促進作用,而高濃度則轉為抑制作用,這種規律也在水稻等作物種子中得到了驗證[17,21-25]。其原因可能是低濃度Na+激活了代謝過程中的某些酶[26],使發芽所需物質合成更加充分,從而使其發芽更加迅速;或者低濃度鹽能促進細胞膜的滲透調節,有利于從逆境中吸水和對抗不良環境[27]。而高濃度的鹽離子毒害作用可能使一些膜蛋白的生理功能發生改變,改變了細胞膜的通透性,最終鹽離子積累抑制了種子的萌發[28]。
與中性鹽相比,堿性鹽脅迫加劇了桑種子的萌發抑制。桂優12號種子在50 mmol/L中性鹽下發芽率高于對照,但在50 mmol/L堿性鹽下發芽率低于對照,桂優62號和8036×農14在堿性鹽下發芽率和發芽勢均低于相同濃度的中性鹽。說明相同鹽濃度處理下,堿性鹽對桑樹種子發芽的抑制作用大于中性鹽。推測在高濃度鹽脅迫下,中性鹽對桑樹種子脅迫主要由Na+造成,而堿性鹽對桑樹種子脅迫除Na+作用外又增加了高pH值的作用,因此堿性鹽對桑樹有更高的脅迫作用。
子葉生長率可以在一定程度上反映植物幼苗生長情況。本研究發現相比中性鹽脅迫,堿性鹽脅迫加劇了對桑幼苗生長的影響。在高濃度鹽堿脅迫下,即使部分幼苗能夠形成葉片,但幼苗的根變短、褐化且末端萎蔫,最終也不能成苗。這可能與鹽分脅迫環境中較高的Na+濃度可能會導致桑種子萌發幼苗體內的活性氧代謝系統的平衡受到影響,清除活性氧的酶促和非酶促系統防御能力下降,并隨著活性氧含量的增大,誘發膜質過氧化作用,引起膜損傷和膜透性加大進而導致一系列生理生化代謝紊亂,使幼苗受到的傷害加重甚至死亡[29]。
結合種子的萌發性狀和子葉生長情況,本研究中的3個桑品種中性鹽抗性由高到低依次為桂優12號、桂優62號、8036×農14,堿性鹽脅迫抗性由高到低依次為桂優62號、桂優12號、8036×農14。
在今后工作中有必要進一步調查更多桑品種的幼齡桑樹、成齡桑樹等不同時期受復合鹽堿脅迫后生長發育的變化情況,為開展不同桑種質資源的耐鹽堿性評價以及耐鹽堿桑樹新品種選育提供理論依據。
參 考 文 獻:
[1]Deinlein U,Stephan A B,Horie T,et al. Plant salt-tolerance mechanisms[J]. Trends Plant Sci.,2014,19(6):371-379.
[2]趙東曉,杜建勛,陳傳杰,等.桑樹鹽堿脅迫研究進展[J]. 山東農業科學, 2015,47(5):132-135.
[3]Shi D C,Yin S J,Yang G H,et al.Citric acid accumulation in an alkali tolerant plant Puccinellia tenuiflora under alkaline stress[J]. Acta Botanica Sinica,2002,44(5):537-540.
[4]張華瑜,包奇軍,柳小寧,等. NaHCO3+Na2CO3對不同啤酒大麥品種萌發期的脅迫效應[J].浙江農業科學,2013(12):1560-1563.
[5]藺吉祥,李曉宇,唐佳紅,等. 鹽堿脅迫對小麥種子萌發、早期幼苗生長及Na+、K+代謝的影響[J].麥類作物學報,2011,31(6):1148-1152.
[6]劉巖, 張薇, 計東風, 等. NaCl脅迫對桑樹種子萌發和 Na+/H+逆向轉運蛋白基因表達的影響[J]. 蠶業科學,2013,39(5):851-857.
[7]于翠,胡興明,鄧文,等. 桑樹耐鹽性研究進展[J]. 蠶桑通報,2012,43(2):6-9,30.
[8]劉雪琴,丁天龍,魏從進,等. 13份桑樹雜交組合F1代的耐鹽性和耐旱性鑒定[J]. 蠶業科學,2014,40(5):764-773.
[9]王玉芳,張蘭.鹽脅迫對糯玉米種子萌發的影響[J]. 山東農業科學,2014,46(3):37-39.
[10]張舟,鄔忠康,陳志成,等. 鹽脅迫對4個品種豇豆種子萌發的影響[J]. 種子,2014,33(3):19-23.
[11]譚衛軍,李其勇,沈雪峰,等. 鹽脅迫對雜草稻和水稻種子萌發特性的影響[J]. 廣東農業科學,2014(3):5-8.
[12]張秀玲. 鹽脅迫下野大豆種子萌發特性研究[J]. 中國野生植物資源,2013,32(6): 21-24.
[13]姜云天,張麗娜,顧地周,等. 鹽脅迫對茶花鳳仙種子萌發的影響[J].東北林業大學學報,2014,42(3):37-41.
[14]卞付萍. 鹽脅迫對水榆花楸種子萌發的影響[J]. 現代園藝,2013(14):12-13.
[15]Wang Z L,Wang G,Liu X M. Germination strategy of the temperate sandy desert annual chenopod Aegriophyllum squarrosum[J]. J. Arid Environ.,1998,40(1):69-76.
[16]緱鋒利.NaCl脅迫下8個苜蓿品種萌發期耐鹽性比較研究[J]. 陜西農業科學,2011(3):78-82,120.
[17]苗昊翠,李利民,宋彬,等.NaCl脅迫對兩種錦雞兒種子萌發的影響[J].新疆農業科學,2011,48(3):498-503.
[18]胡生榮,高永,武飛,等. 鹽脅迫對兩種無芒雀麥種子萌發的影響[J].植物生態學報,2007,31(3):513-520.
[19]楊帆,丁菲,杜天真. 鹽脅迫對構樹種子萌發及幼苗生理特性的影響[J]. 種子,2008,27(8):68-71.
[20]張瑞富,王云,喬宏偉,等. 鹽脅迫對不同品種小麥發芽的影響[J]. 內蒙古民族大學學報,2007,22(3):297-301.
[21]斯琴巴特爾,吳紅英. 鹽脅迫對玉米種子萌發及幼苗生長的影響[J].干旱區資源與環境,2000,14(4):76-80.
[22]楊秀玲,郁繼華,李雅佳,等. NaCl脅迫對黃瓜種子萌發及幼苗生長的影響[J]. 甘肅農業大學學報,2004,39(1):6-9,17.
[23]時麗冉,崔興國,劉志華,等. 混合鹽堿脅迫對旱稻種子萌發的影響[J].種子,2006,25(2):25-27,31.
[24]徐珊珊,葉景學,張廣臣.鹽堿脅迫對辣椒種子萌發的影響[J]. 種子,2011,30(3):85-87,90.
[25]張學云,趙丹,袁慶華. 多花胡枝子野生種質材料芽期耐鹽性評價[J]. 種子,2011,30(9):73-78.
[26]王慶亞,劉敏,張守棟,等. 鹽脅迫對鹽角草種子萌發與幼苗生長效應的研究[J]. 江蘇農業科學,2002(2):69-71.
[27]劉祖祺,張石城.植物抗性生理學[M]. 北京:中國農業出版社,1994.
[28]祝娟娟,丁天龍,魏從進,等. 鹽脅迫下不同桑品種種子萌發特性研究[J]. 蠶學通訊,2013,33(1):1-6.
[29]喬慧萍,李建設,雍立華,等. 植物鹽脅迫生理及其適應性調控機制的研究進展[J]. 寧夏農林科技,2007(3):34-36,24.
