新德里金屬-β-內酰胺酶-1的結構、催化機理及其抑制劑研究進展
2017-07-06 10:29:05胡建平
成都大學學報(自然科學版) 2017年2期
關鍵詞:結構
胡建平, 左 柯, 萬 華
(1.成都大學 藥學與生物工程學院, 四川 成都 610106;2.成都大學 四川抗菌素工業研究所, 四川 成都 610052;3.華南農業大學 數學與信息學院, 廣東 廣州 510642)
?
新德里金屬-β-內酰胺酶-1的結構、催化機理及其抑制劑研究進展
胡建平1,2, 左 柯1,2, 萬 華3
(1.成都大學 藥學與生物工程學院, 四川 成都 610106;2.成都大學 四川抗菌素工業研究所, 四川 成都 610052;3.華南農業大學 數學與信息學院, 廣東 廣州 510642)
β-內酰胺類抗生素,作為一線抗生素被廣泛用于各類細菌感染的臨床治療.由位于質粒的基因blaNDM-1編碼的新德里金屬-β-內酰胺酶-1可高效水解幾乎所有β-內酰胺類抗生素,對β-內酰胺類抗生素的應用造成了極大的威脅,開發有效的抑制劑新德里金屬-β-內酰胺酶-1抑制劑迫在眉睫.而抑制劑分子的設計改造工作的開展有賴于對新德里金屬-β-內酰胺酶-1結構和催化功能的研究,本綜述旨在總結目前有關新德里金屬-β-內酰胺酶-1結構、催化機理及其抑制劑的相關研究進展,為基于新德里金屬-β-內酰胺酶-1結構的藥物分子設計、改造等研究提供幫助.
細菌耐藥;NDM-1;催化機理;抑制劑;藥物設計
0 引 言
自1929年,人類發現第一種抗生素——青霉素以來,抗生素已挽救了無數的生命.目前,以青霉素和頭孢菌素為代表的β-內酰胺類抗生素,是治療細菌感染的常用藥物,該類抗生素的化學結構中均含有一個發揮藥效必需的四元β-內酰胺環,其化學結構如圖1所示.當其作用于細菌時,β-內酰胺類抗生素的β-內酰胺環開環,與細菌細胞壁合成途徑中的羧肽酶和轉肽酶發生酰化作用,抑制該酶的生物活性而使合成的細胞壁出現缺陷,最終導致菌體裂解死亡[1-2].
隨著抗生素的廣泛應用,細菌耐藥問題也日益凸顯[3],同時攜帶多種耐藥基因的“超級細菌",如泛耐藥鮑曼不動桿菌、耐碳青霉烯肺炎克雷伯菌、銅綠假單胞菌等也頻頻被報道[4-6].在細菌耐藥機制中,產生一種或多種水解酶或鈍化酶滅活抗菌藥物最為常見,而β-內酰胺酶是一種可催化β-內酰胺環水解開環的水解酶.基于氨基酸序列的同源性[7],β-內酰胺酶可分為A、B、C和D 4類.其中,A、C和D類酶的活性位點為絲氨酸殘基,故又統稱為絲氨酸-β-內酰胺酶(serine-β-lactamase,SBL);而B類酶則依靠1個或2個金屬離子行使催化功能,因此又稱為金屬-β-內酰胺酶(metallo-β-lactamase,MBL),具體可細分為B1、B2和B3 3個亞類.
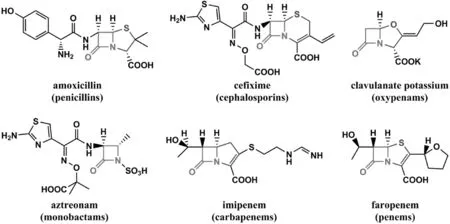
圖1 β-內酰胺類抗生素的化學結構
新德里金屬-β-內酰胺酶-1(New Delhi metallo-β-lactamase-1,NDM-1)是一種廣譜高效的B1 MBL,于2009年首次被報道[8-9].目前,研究人員已發現同類的另外14個變種[10],依次命名為NDM-2~NDM-15.NDM-1由位于質粒的blaNDM-1基因編碼,可水解除氨曲南和美西林外幾乎所有的β-內酰胺類抗生素[8,10-11].研究發現,blaNDM-1基因并不僅限于在醫院獲得性菌種間傳遞,同時也可在社區高度流行的菌種間傳播[12-13],現已發現近10種細菌表達NDM-1[14].這些表達NDM-1的細菌可引起包括尿道和肺部感染、腹膜炎、軟組織感染及敗血癥等疾病,為相關病癥的臨床治療造成了嚴重威脅.
隨著檢測技術的發展,比如,改良Hodge試驗[15-16]和基于顯色培養基篩選法[17-19]等表型方法,基于PCR的體外蛋白表達技術[20]、基質輔助激光解吸電離飛行時間質譜法[21-22]等基因型方法,以及分子方法均可有效檢測出產NDM-1的菌株.同時,在治療方面,雖然有報道聯用多種抗菌藥物并口服磷霉素成功治愈產NDM-1肺炎克雷伯菌引發的尿道感染的個案[23],但目前仍缺乏對產NDM-1菌株特效的治療藥物.
考慮到β-內酰胺類抗生素良好的藥效和較低的毒副作用,開發有效的NDM-1抑制劑并與β-內酰胺類抗生素配伍使用是目前主要的研發思路.而掌握NDM-1的化學結構、催化機理等生物化學信息,可為其抑制劑的設計、改造等提供有力的技術支撐.
1 NDM-1的分子結構
獨特的分子結構是NDM-1發揮水解功能的基礎,同時也是篩選、設計抑制劑的重要依據.截至2017年3月,RCSB蛋白質數據庫(http://www.rcsb.org/)共收錄了30個NDM-1晶體結構,除3S0Z(PDB ID)為大腸桿菌NDM-1外,其余均來自肺炎克雷伯菌.NDM-1為雙金屬MBL[24-27],在30個NDM-1晶體結構中,活性口袋中含有金屬離子的結構有23個(3SFD為單鋅離子結構),同時結合β-內酰胺類抗生素的結構縮為14個,其中結合抑制劑的結構有3個(5A5Z、4U4L、4EXS).表1列出了NDM-1-底物/抑制劑復合物晶體結構的具體信息.

表1 NDM-1-底物/抑制劑復合物的晶體結構信息
1.1 NDM-1的整體結構
NDM-1由單多肽鏈構成,共含有270個氨基酸殘基,呈典型的αβ/βα夾層結構[28-30],具體如圖2所示.
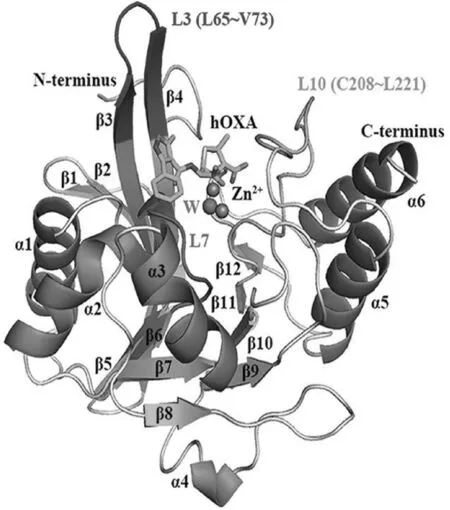
圖2 NDM-1的整體結構
從圖2中可以看出,分子暴露在溶劑中的兩側分別為2段α螺旋,從N端往C端依次為α1、α2、α5和α6;2個β折疊通過一段α-β-α的結構相連接位于分子的內層.其中,靠近N端的β折疊由7股平行或反平行的β折疊片構成,而靠近C端的β折疊則由4股組成[26].3段loop區L3(L65-V73)、L7(T119~M126)和L10(C208~L221)組成了活性口袋的主體部分,具有催化活性的2個Zn2+(Zn1與Zn2)則位于口袋底部[31].
此外,研究還發現,在細菌體內,NDM-1以單體形式發揮催化活性[31],而目前解析得到的NDM-1晶體結構則多為二聚體,其在膜結合與純化狀態下能通過疏水與范德華相互作用進行二聚被推測與該酶獨特的耐藥機制有關[28].
1.2 NDM-1的活性位點
圖3給出了NDM-1活性位點的結構:位于活性中心的Zn1與H120、H122、H189及水分子/OH-(與晶體生長時的pH有關[27,35])結合構成四面體構型,即組氨酸位點;Zn2與D124、C208和H250結合構成三角錐形結構形成半胱氨酸位點;Zn1與Zn2通過D124的側鏈相聯系[26,30-31].
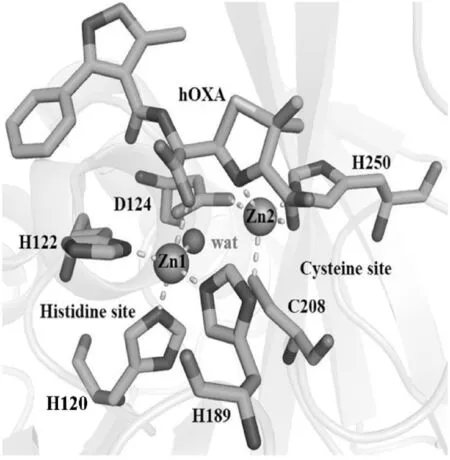
圖3 NDM-1活性位點的局部放大圖
從圖3可以看出,Zn1與水解后的苯唑西林(oxacillin,OXA)的β-羧基間存在相互作用,Zn2還與噻唑環上的N和羧基配位結合,提示Zn2+在NDM-1與底物的結合和催化水解反應的過程中均具有重要作用.Zn2+與底物間的這種相互作用,使β-內酰胺環維持在易被進攻的空間取向狀態,從而使得水解反應得以高效進行[36].晶體學實驗也發現,Zn2+可以被其他金屬離子,如Cd2+、Mn2+、Co2+等,替換[30].
Yang等[37]測定了野生型NDM-1和Zn/Co、Co/Co和Co/Cd 3種金屬離子替換后的NDM-1水解chromacef的動力學常數,以及紫外—可見光光譜(UV-vis)、核磁共振氫譜(1H NMR)、電子順磁共振譜(EPR)、X射線吸收精細結構譜(EXAFS)等數據,結果顯示:金屬替換后,NDM-1的酶促動力學常數出現明顯變化,具有順磁性的金屬取代NDM-1,比如Co/Cd和Zn/Co,與底物反應時有顯著的結構變化;與其他B1 MBLs相比,NDM-1的活性位點擁有更大的表面積,這顯然與構成活性位點的L3和L10均距離鋅離子中心較遠有關[28].Yuan等[38]通過分子對接預測了160種β-內酰胺類抗生素在NDM-1活性位點處的構象,發現NDM-1在結合底物后,其活性位點構象并未出現較為顯著的變化.這為NDM-1水解底物的廣譜性提供了重要的結構基礎.
此外,科研人員對活性位點的結構分析還發現一些關鍵識別殘基,例如:K125通過與周圍殘基形成的氫鍵網絡穩定L7的構象,同時對與Zn1結合的殘基有一定的空間位阻排斥效應[27];Y229對穩定活性位點的構象同樣具有重要意義[27,31];定點突變L65A導致氨芐西林和一些頭孢菌素的最低抑菌濃度下降,而定點突變N220A則導致NDM-1對氨芐西林、頭孢吡肟、美羅培南和亞胺培南的水解能力大大降低[39].Zhu等[40]對NDM-1與氨芐西林、頭孢硝噻吩和美羅培南形成的3個復合物體系進行了30 ns的分子動力學模擬發現,位于L3上的L65、M67和F70均與氨芐西林和頭孢硝噻吩的側鏈形成了較強的疏水作用,此對于維持復合物的穩定性具有重要意義,并給出了L3在整個模擬過程中具有3簇主要構象,并呈現出較大的片段柔性.相關研究表明:Q123與D124側鏈的氧原子可與臨近的β內酰胺類抗生素側鏈形成氫鍵[27];K211、G219、N220與H189在IMP-1、CcrA等同類MBLs中高度保守,K211除了參與底物的識別與水解反應外,還可通過與底物形成鹽鍵維持其有利于水解反應的正確取向;N220可與底物的羰基氧原子形成氫鍵,還可與Zn1形成相互作用,后者可能與維持水解反應中的四面體中間產物的穩定有關[26-28,39,41].
2 NDM-1的催化機理
作為B1類MBL成員之一,NDM-1催化底物水解的機理與同類MBLs類似[27],主要包括,β-內酰胺羰基與Zn1結合,OH-的親核進攻,質子向N原子的轉移和C-N鍵的斷裂,具體如圖4所示.但其水解機理中的具體細節尚未被全部闡明,還有待進一步的探究.
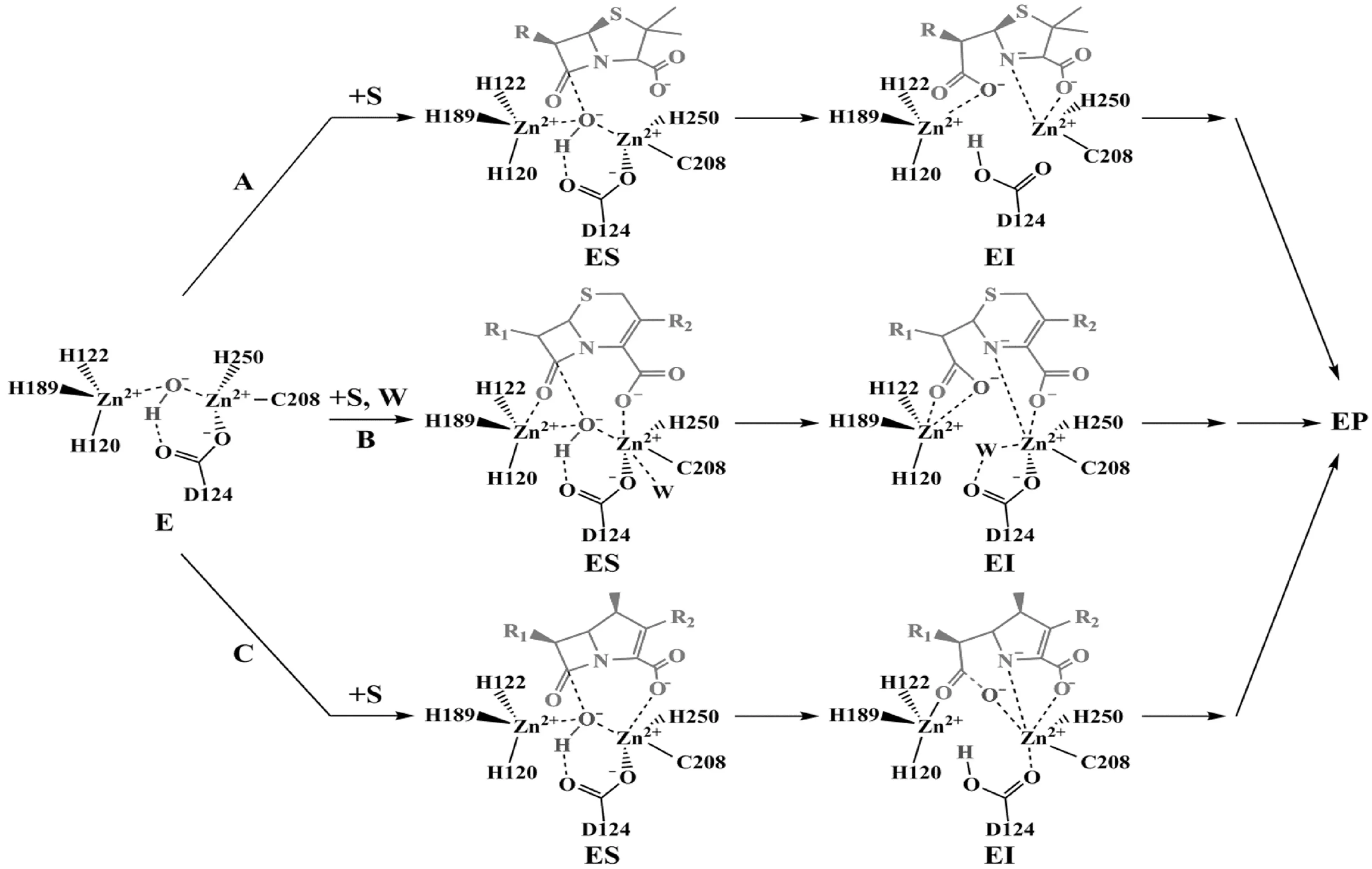
A表示青霉素類β-內酰胺類抗生素時可能的水解途徑;B表示頭孢菌素類β-內酰胺類抗生素時可能的水解途徑;C表示碳青霉烯類β-內酰胺類抗生素時可能的水解途徑.E表示NDM-1,S表示底物,I表示中間體,P表示產物,W表示水.機理的繪制參考文獻[37,42,45].
圖4 NDM-1可能的催化機理
關于反應中的親核試劑和廣義堿,有學者認為,溶劑環境中的水分子在水解反應中作為親核試劑,而位于活性位點中的OH-作為廣義堿,溶劑環境中的水分子向OH-給出質子后生成新的OH-,然后進攻β-內酰胺環的羰基碳原子,該反應的發生需要越過約80 kJ/mol的能壘[27,35];也有學者認為,親核試劑可能是位于活性位點的水分子,而D124或環境中的水分子作為廣義堿接受質子,該情況下的反應能壘大于或等于200 kJ/mol[42].
有學者認為,按照其他MBLs的反應機理,位于Zn1和Zn2間的OH-作為親核試劑進攻β-內酰胺環后,C-N鍵可立即斷裂形成氮負離子中間體[43].但Zhang等[42]提出,β-內酰胺環受到OH-進攻后,C-N鍵的斷裂可能與負離子的質子化反應協同進行,并不產生氮負離子中間體.但無論是上述哪種情況,氮原子上的質子均來自親核進攻的OH-.而科研人員在NDM-1Δ6、NDM-1Δ21和NDM-1Δ36(分別表示移除分子的前6、21和36個氨基酸)的酶促動力學研究過程中發現,NDM-1的催化機理中可能存在不只一個中間體[44].在探究NDM-1對頭孢菌素類β-內酰胺類抗生素的水解機制中,Feng等[25]通過1H NMR監測到了二氫噻嗪環中雙鍵轉移的情況,最終質子并不是轉移至N原子,而是根據底物的結構遷移至不同的部位,并得到了反應中間體的晶體結構.
此外,研究人員在對金屬取代NDM-1催化機理的研究中還發現:金屬離子的種類并不能顯著影響反應的能壘;產物的釋放速率與半胱氨酸位點的金屬路易斯酸性大小有關(Zn>Co>Cd),但同時也受到組氨酸位點處金屬種類的影響;當用Co2+離子取代Zn1時,生成產物的速率比未取代時快2~3倍,這可能與Co2+離子改變了與底物反應時的電子結構,降低了電子結構對稱性有關;當用Cd2+離子取代Zn2時,中間體的生成速率有明顯影響,提示Zn2在親核進攻和穩定中間體的過程中具有重要意義[37].
目前,分子動力學模擬和混合量子力學/分子力學模擬等理論研究方法在酶催化反應的機理研究中應用廣泛.Zheng等[42]使用SCC-DFTB/CHARMM混合力場模擬了NDM-1與底物氨芐西林和抑制劑L-卡托普利的結合模式,并根據密度泛函理論對氨芐西林的開環過程進行了理論計算,模擬結果顯示:OH-位于兩個Zn2+的中間位置,并通過氫鍵與D124相連;氨芐西林的羧基并未與Zn2相連,而是通過氫鍵與K211和N220連接;后續OH-親核進攻、C-N鍵斷裂、形成氮負離子中間體以及開環后羧基與Zn2配位鍵的形成幾乎同時進行(見圖4).Zhu等[45]使用ONIOM算法對NDM-1催化美羅培南的水解機理研究發現,離子化的D124是C-N鍵斷裂的有利因素;在沒有水分子與Zn2結合時,反應過程中更傾向于形成氮負離子中間體,且K211在底物結合和催化反應的過程中均具有重要意義.由于NDM-1高效的水解速率,目前很難通過實驗對理論計算得出的過渡態進行驗證.相關研究也表明,針對不同類型的底物NDM-1的催化機理各不相同.為了指導設計開發具有臨床應用價值的抑制劑,NDM-1催化下的水解機理還有待在分子層面做進一步探究.
3 NDM-1抑制劑的研究現狀
目前,科研人員為應對NDM-1帶來的耐藥問題采取的主要策略為:在了解NDM-1結構及可能水解機制的情況下,不斷開發或尋找不易被NDM-1水解的新抗生素;篩選設計NDM-1抑制劑,并與傳統抗生素制成合劑.
3.1 含硫類抑制劑
硫元素是生命活動中的重要元素之一,也常出現在各類藥物分子中.含硫類抑制劑的結構式及IC50值如圖5所示.
血管緊張素轉化酶抑制劑卡托普利中含有巰基結構,是經典的抗高血壓藥物.研究發現,D-卡托普利1對NDM-1表現出較好的抑制活性(IC50=7.9 μM),而L-卡托普利的抑制活性相對較弱,其IC50為202.0 μM[31].Li等[46]為進一步探究NDM-1抑制劑的藥效基團,設計合成并測試了系列D-卡托普利類似物2-6的抑制活性,發現R構型的芐胺類似物6a表現出了較高的抑制活性,其IC50值為1.5 μM,遠高于S型旋光異構體.Shen等[47]發現噻吩羧酸類衍生物7-9對NDM-1具有一定的抑制作用,與美羅培南聯合給藥可提高美羅培南的抗菌活性.此外,Wang等[48]通過多步虛擬篩選從含有280萬個類藥分子的ZINC化合物庫中篩出3個磺胺類NDM-1抑制劑10-12,其分子動力學模擬顯示,分子中的磺酰胺基直接與Zn1結合,從而阻礙水解反應的發生,提示磺酰胺基可能是開發NDM-1抑制劑式可選的優勢骨架結構.
3.2 苯并吡喃類抑制劑
苯并吡喃類NDM-1抑制劑的結構如圖6所示.
利用分子對接方法,Proschak等[49]將SPECS數據庫中的分子對接至NDM-1等4種MBLs晶體結構中.分子對接同時使用MOE、GOLD和PLANTS 3款軟件進行,以3款軟件預測結合模式的均方根偏差小于或等于0.2 nm為判據進行分子片段篩選;在篩得的分子片段中,僅保留MBLs中結合模式均方根偏差值小于或等于0.2 nm的片段進行Fluorocillin試驗評價活性,由此得到抑制活性最好的苯并吡喃類化合物13(IC50=50±9 μM)[50].此外,Gan等[51]還從青霉菌株(Penicillium sp. 109F 484)中分離得到2種具有微弱NDM-1抑制活性的聚酮化合物14和15,二者的IC50分別為94.9 μM和87.9 μM.抗菌實驗顯示,化合物14和15對產NDM-1肺炎克雷伯菌均無抑制活性,與美羅培南聯合使用時,也無法提高美羅培南的抗菌活性.
圖5 含硫類NDM-1抑制劑的結構
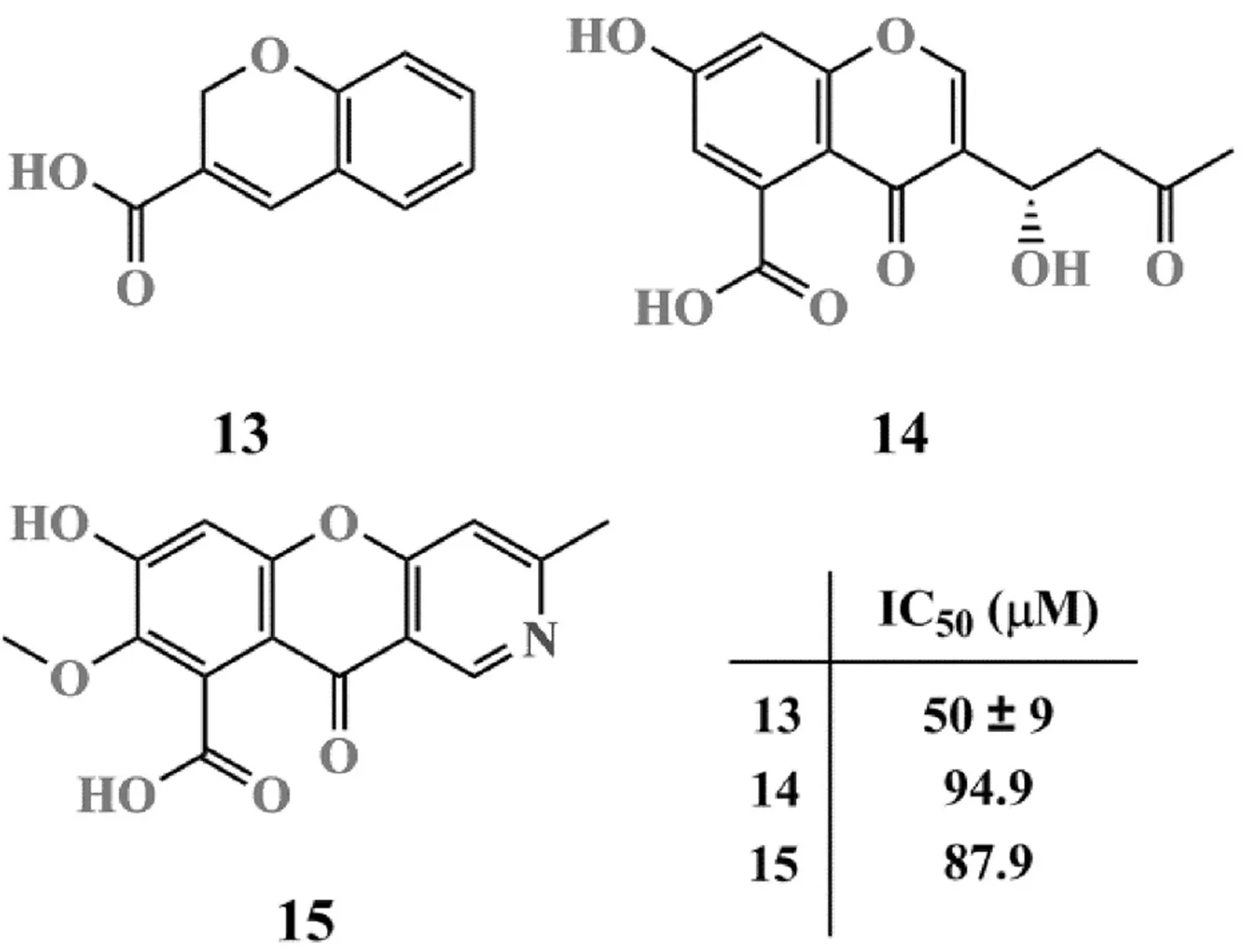
圖6 苯并吡喃類NDM-1抑制劑的結構
3.3 共價結合型抑制劑
共價結合型NDM-1抑制劑的結構如圖7所示.
研究發現,通過氫鍵、范德華等非鍵相互作用與靶點可逆性結合是大多數藥物在發揮藥效時與靶點的結合特點,但也不乏通過形成共價鍵與靶點不可逆結合的例子,如細胞毒類抗腫瘤藥物.在NDM-1共價結合型抑制劑的開發過程中,Christopeit等[52]報道了2種可逆共價結合型非β-內酰胺類抑制劑16(Kd=181 μM)和17(Ki=580 nM),其電噴霧離子化質譜和氨基酸定點突變實驗顯示,化合物16和17通過單鍵與NDM-1活性位點的K224結合;通過共價分子對接還發現,化合物17的酮羰基與Zn2之間存在極性相互作用,并與W87、I35和V67間形成輸水相互作用.Thomas等[53]通過高通量篩選也得到了2種共價結合型NDM-1抑制劑,對氯汞苯甲酸18和硝普鈉19,兩者的抑制活性(IC50)分別為2.3 μM和9.0 μM,通過質譜聯用技術檢測和C208D突變實驗發現,化合物18可以選擇性共價修飾NDM-1活性位點結合Zn2的C208,從而發揮抑制作用.另外,Thomas等[54]還報道了頭孢克洛20通過K211介導的多步反應,不可逆地共價抑制NDM-1的活性(Ki=2.3±0.1 mM),提示K211可能是開發共價結合型抑制劑的關鍵氨基酸殘基.依布硒啉21是一種新型非甾體抗炎藥,臨床上主要用于缺血和中風的治療,研究發現:依布硒啉可與鍵合Zn2的C208形成S-Se鍵,從而影響NDM-1的催化功能;以(1.3~1.4)∶1的比例聯合使用依布硒啉和氨芐西林或美羅培南,可使得β-內酰胺類抗生素的MICs分別下降4和35倍[55].
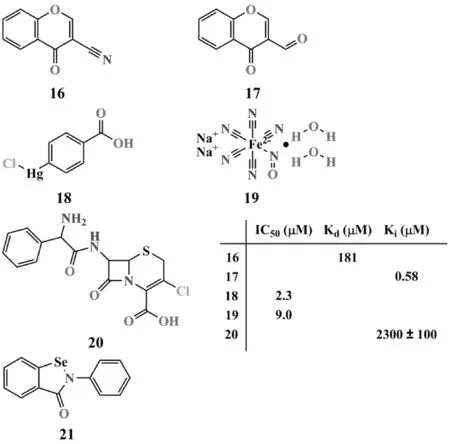
圖7 共價結合型NDM-1抑制劑的結構
3.4 金屬螯合劑
金屬離子作為NDM-1發揮催化功能的關鍵輔助因子,若使用金屬螯合劑封閉NDM-1活性位點的Zn2+,理論上可有效抑制其催化活性.典型NDM-1金屬螯合劑的結構如圖8所示.
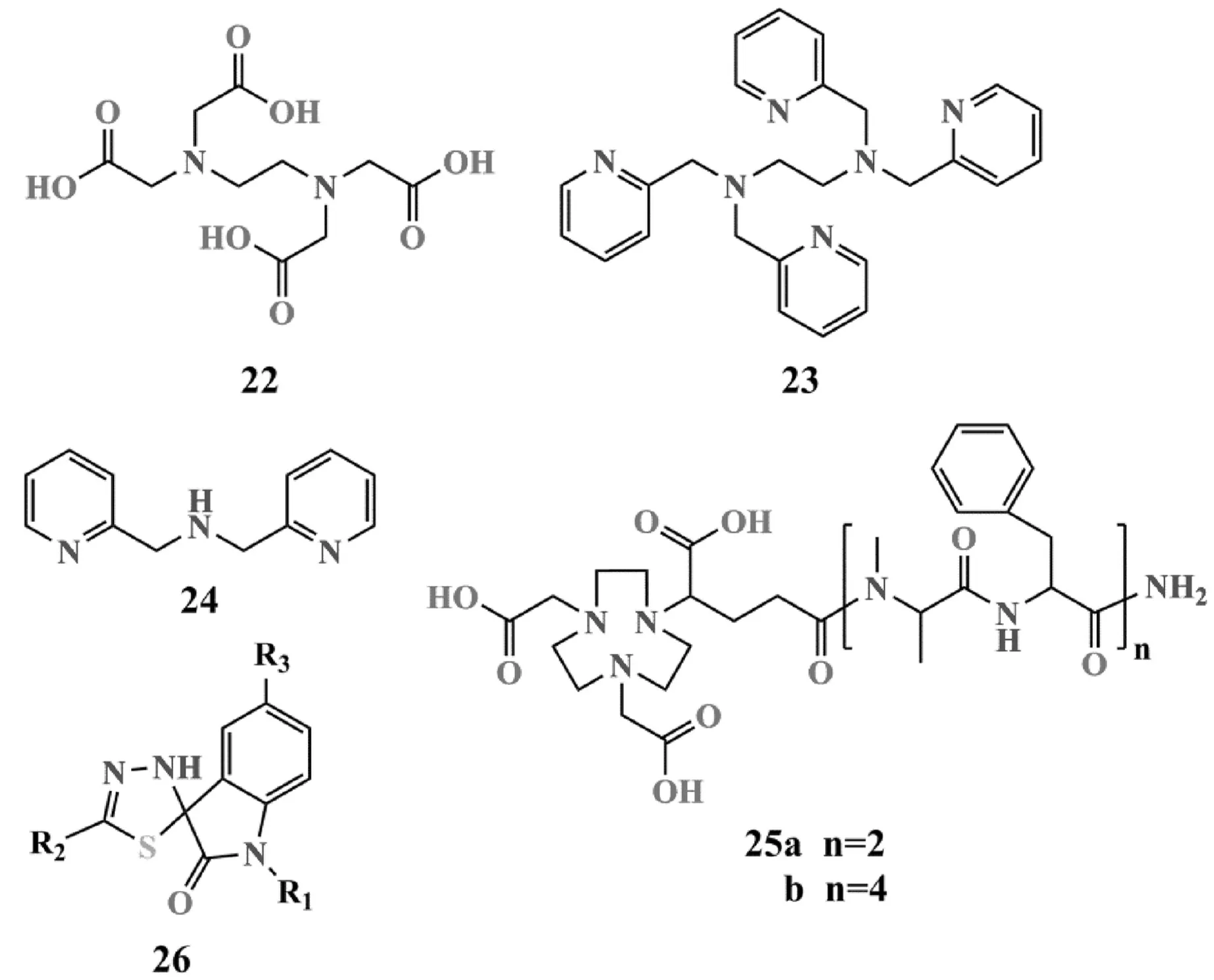
圖8 金屬螯合劑的結構
研究發現,乙二胺四乙酸22是常見的金屬螯合劑,可有效抑制NDM-1(IC50=1.6 μM),但由于其具有較強的毒性,并不適合直接用于臨床治療[56].Azumah等[57]評價了3種金屬螯合劑23~25在與美羅培南聯合使用時,NDM-1對美羅培南的耐藥情況:當化合物23用量在4和8 mg/L時活性最佳,可使美羅培南的MICs分別降低至0.5和0.06 mg/L;化合物24在8和16 mg/L的劑量時,同樣也可將美羅培南的MICs分別降至1和0.125 mg/L;但化合物25對NDM-1的抑制效果較差,需要將用量提至32 mg/L才可將美羅培南的MICs降低至0.06 mg/L.此外,Falconer等[58]還報道了一類二氫吲哚螺噻二唑類金屬螯合劑26,并通過產NDM-1肺炎克雷伯菌小鼠感染模型證明了該類化合物對NDM-1的抑制活性,其單晶結構顯示,原化合物26的S-Cspiro鍵斷裂,噻二唑環開環形成亞胺結構,暴露的S與亞胺N構成二齒配體與Zn2+進行螯合,從而發揮抑制作用.
3.5 其 他
其他NDM-1抑制劑分子的結構如圖9所示.
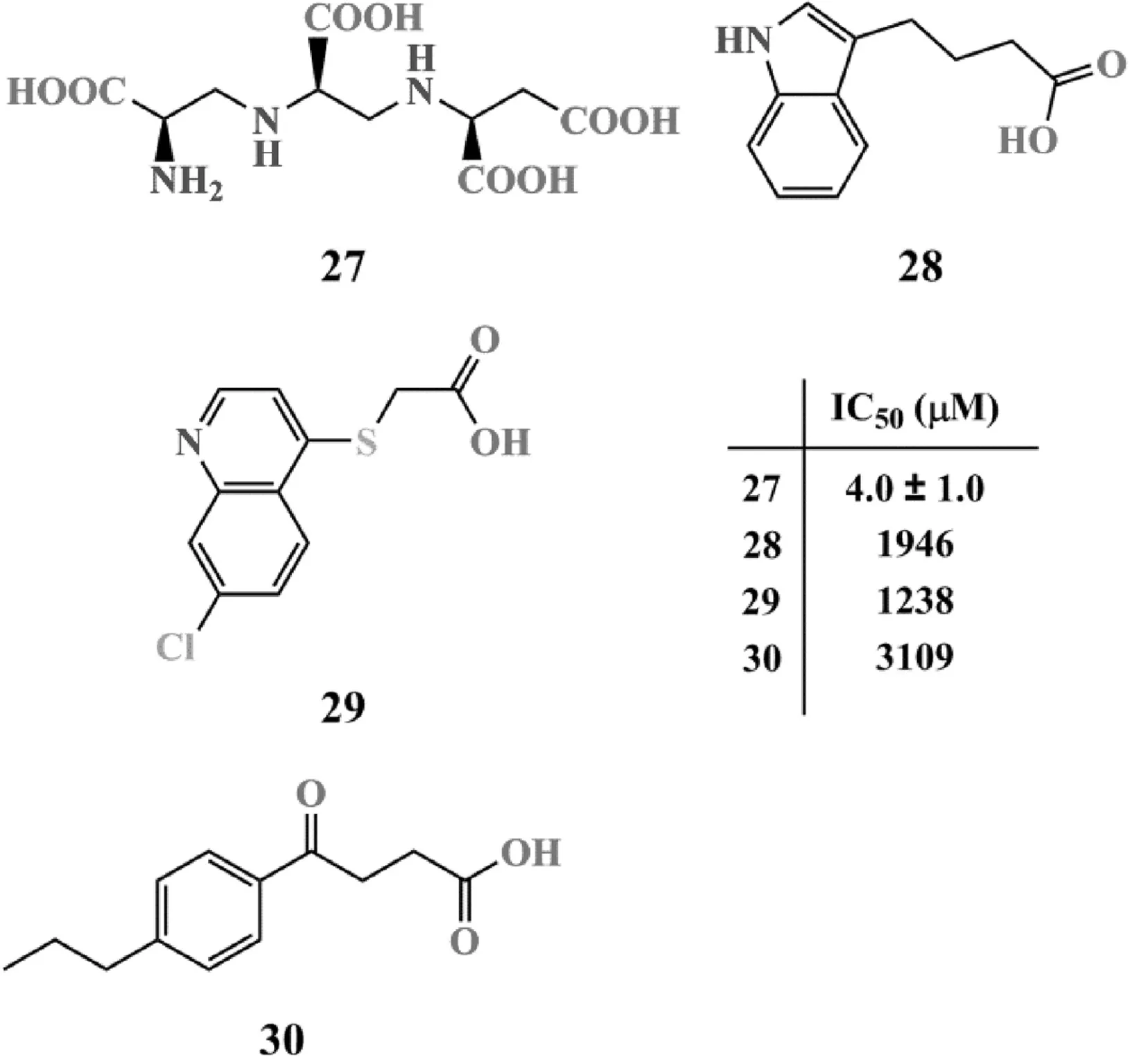
圖9 其他NDM-1抑制劑分子的結構
曲霉明A 27是一種從雜色曲霉菌株中分離得到的一種天然產物.研究發現,曲霉明對血管緊張素轉化酶和內皮素轉化酶均有良好的抑制作用,其IC50值分別為1.2 μM和3.4 μM[59-61].進一步研究發現,曲霉明對NDM-1的抑制作用同樣出色(IC50=4.0±1.0 μM)[62].研究人員利用電感耦合質譜證實,曲霉明對NDM-1的抑制主要通過耗盡NDM-1活性位點中的Zn2+實現,屬于一種不可逆抑制作用,但這種抑制作用造成的失活狀態可通過加入過量的ZnSO4實現完全復活[62].雖然曲霉明在抑制活性和毒性等方面的表現令人滿意,但由于其較高的親水性,其藥動學性質還有待進一步改善[62].Chrestopeit等[63]使用正交篩選聯合表面等離子共振技術得到了另外3種對NDM-1有一定抑制作用的片段分子28~30,其QM-polarized分子對接結果顯示:28~30的羧基可與Zn2結合,并與K224和N233形成氫鍵;28和30與W87、L59和V67間還存在一定的疏水作用.這些結合力可能是維持這些片段分子發揮抑制活性的關鍵因素.
4 展 望
產NDM-1的耐藥菌感染對使用β-內酰胺類抗生素的臨床治療造成了嚴重威脅.自2009年首次報道NDM-1至今,各國學者已在NDM-1的結構、催化機理及其抑制劑等領域開展了一系列研究,并基于不同底物提出了NDM-1多種可能的催化機理,但對于催化機理中的堿、親核試劑及中間體質子化過程等具體細節仍在探討之中.就目前開發的抑制劑而言,抑制活性普遍維持在微摩爾級,雖在體外或體內均取得了較為滿意的結果,但又由于分子固有的毒性或較差的藥代動力學性質等,尚未能成功開發為藥物上市銷售.
隨著各學科的交叉融合及大數據時代的來臨,對NDM-1研究而言,根據量子或分子力學等理論,通過計算機模擬NDM-1的水解機理或其與抑制劑分子間的結合模式,并通過數學建模定量探究分子結構與活性之間的關系,為NDM-1抑制劑分子的設計與改造提供理論指導等已取得了一定的成效.隨著計算科學的飛速發展,模擬精度的逐步提高,藥學科研工作中計算機輔助的應用將會越來越多,相信在不久的將來,基于理論的計算可望成為應對諸如細菌耐藥問題等各類醫、藥學難題時不可缺少的主要工具之一.
[1]Wilke M S,Lovering A L,Strynadka N C.Beta-lactamantibioticresistance:acurrentstructuralperspective[J].Curr Opin Microbiol,2005,8(5):525-533.
[2]Rawat D,Nair D.Extended-spectrumβ-lactamasesingramnegativebacteria[J].J Glob Infect Dis,2010,2(3):263-274.
[3]World Health Organization.Antimicrobialresistance:globalreportonsurveillance2014[R].Geneva,Switzerland:World Health Organization,2014.
[4]Protic D,Pejovic A,Andjelkovic D,et al.Nosocomialinfectionscausedbyacinetobacterbaumannii:arewelosingthebattle?[J].Surg Infect,2016,17(2):1-7.
[5]Stoesser N,Mathers A J,Moore C E,et al.Colistinresistancegenemcr-1andpHNSHP45plasmidinhumanisolatesofescherichiacoliandklebsiellapneumoniae[J].Lancet Infect Dis,2016,16(3):285-286.
[6]Labarca J A,Salles M J C,Seas C,et al.Carbapenemresistanceinpseudomonasaeruginosaandacinetobacterbaumanniiinthenosocomialsettinginlatinamerica[J].Crit Rev Microbiol,2016,42(2):276-292.
[7]Bush K,Jacoby G A.Updatedfunctionalclassificationofbeta-lactamase[J].Antimicrob Agents Chemother,2010,54(3):969-976.
[8]Yong D,Toleman M A,Giske C G,et al.Characterizationofanewmetallo-beta-lactamasegene,bla(NDM-1),andanovelerythromycinesterasegenecarriedonauniquegeneticstructureinKlebsiellapneumoniaesequencetype14fromIndia[J].Antimicrob Agents Chemother,2009,53(12):5046-5054.
[9]Moellering R C J.NDM-1—acauseforworldwideconcern[J].New Engl J Med,2010,363(25):2377-2379.
[10]Groundwater P W,Xu S,Lai F,et al.Newdelhimetallo-β-lactamase-1:structure,inhibitorsanddetectionofproducers[J].Future Med Chem,2016,8(9):993-1012.
[11]Marrs E C,Day K M,Perry J D.InvitroactivityofmecillinamagainstenterobacteriaceaewithNDM-1carbapenemase[J].J Antimicrob Chemother,2014,69(10):2873-2875.
[12]Denis C,Poirel L,Carricajo A,et al.NosocomialtransmissionofNDM-1-producingescherichiacoliwithinanon-endemicareainFrance[J].Clin Microbiol Infec,2012,18(5):E128-E130.
[13]Walsh T R,Weeks J,Livermore D M,et al.DisseminationofNDM-1positivebacteriainthenewdelhienvironmentanditsimplicationsforhumanhealth:anenvironmentalpointprevalencestudy[J].Lancet Infect Dis,2011,11(5):355-362.
[14]Berrazeg M,Diene S,Medjahed L,et al.Newdelhimetallo-beta-lactamasearoundtheworld:aneReviewusinggooglemaps[J].Euro Surveil,2014,19(20):2-15.
[15]Girlich D,Poirel L,Nordmann P.Valueofthemodifiedhodgetestfordetectionofemergingcarbapenemasesinenterobacteriaceae[J].J Clin Microbiol,2012,50(2):477-479.
[16]Abidin N Z Z,Sulong A,Alfizah H,et al.Screeningfornewdelhimetallo-β-lactamase-1inenterobacteriaceae:istherearoleforthemodifiedhodgetest?[J].Pak J Med Sci,2015,31(6):1340-1343.
[17]Perry J D,Naqvi S H,Mirza I A,et al.PrevalenceoffaecalcarriageofenterobacteriaceaewithNDM-1carbapenemaseatmilitaryhospitalsinpakistan,andevaluationoftwochromogenicmedia[J].J Antimicrob Chemother,2011,66(10):2288-2294.
[18]Nordmann P,Poirel L,Walsh T R,et al.TheemergingNDMcarbapenemases[J].Trends Microbiol,2011,19(12):588-595.
[19]Nordmann P,Poirel L,Carrr A,et al.Howtodetectndm-1producers[J].J Clin Microbiol,2011,49(2):718-721.
[20]Huang L,Hu X,Zhou M,et al.RapiddetectionofNewdelhimetallo-β-lactamasegeneandvariantscodingforcarbapenemaseswithdifferentactivitiesbyuseofaPCR-basedinvitroproteinexpressionmethod[J].J Clin Microbiol,2014,52(6):1947-1953.
[21]Hrabák J,Studentová V,Walková R,et al.DetectionofNDM-1,VIM-1,KPC,OXA-48,andOXA-162carbapenemasesbymatrix-assistedlaserdesorptionionization-timeofflightmassspectrometry[J].J Clin Microbiol,2012,50(7):2441-2443.
[22]Hrabák J,Chudácková E,Walková R.Matrix-assistedlaserdesorptionionization-timeofflight(maldi-tof)massspectrometryfordetectionofantibioticresistancemechanisms:fromresearchtoroutinediagnosis[J].Clin Microbiol Rev,2013,26(1):103-114.
[23]Wilkowski P,Ciszek M,Dobrzaniecka K,et al.SuccessfultreatmentofurinarytractinfectioninkidneytransplantrecipientscausedbymultiresistantklebsiellapneumoniaeproducingnewDelhimetallo-beta-lactamase(NDM-1)withstrainsgenotyping[J].Transplant Proc,2016,48(5):1576-1579.
[24]Klingler F,Wichelhaus T A,Frank D,et al.Approveddrugscontainingthiolsasinhibitorsofmetallo-β-lactamases:strategytocombatmultidrug-resistantbacteria[J].J Med Chem,2015,58(8):3626-3630.
[25]Feng H,Ding J,Zhu D,et al.StructuralandmechanisticinsightsintoNDM-1catalyzedhydrolysisofcephalosporins[J].J Am Chem Soc,2014,136(42):14694-14697.
[26]King D T,Worrall L J,Gruninger R,et al.NewDelhimetallo-β-lactamase:structuralinsightsintoβ-lactamrecognitionandinhibition[J].J Am Chem Soc,2012,134(28):11362-11365.
[27]Zhang H,Hao Q.CrystalstructureofNDM-1revealsacommonβ-lactamhydrolysismechanism[J].FASEB J,2011,25(8):2574-2582.
[28]King D,Strynadka N.Crystalstructureofnewdelhimetallo-β-lactamaserevealsmolecularbasisforantibioticresistance[J].Protein Sci,2011,20(9):1484-1491.
[29]Kim Y,Tesar C,Mire J,et al.Structureofapo-andmonometalatedformsofNDM-1—ahighlypotentcarbapenem-hydrolyzingmetallo-β-lactamase[J].Plos One,2011,6(9):e24621.
[30]Green V L,Verma A,Owens R J,et al.Structureofnewdelhimetallo-β-lactamase1(NDM-1)[J].Acta Crystallogr Sect F Struct Biol Cryst Commun,2011,67(10):1160-1164.
[31]Yu G,Jing W,Niu G,et al.AstructuralviewoftheantibioticdegradationenzymeNDM-1fromasuperbug[J].Protein Cell,2011,2(5):384-394.
[32]Aitha M,Moller A J,Sahu I D,et al.InvestigatingthepositionofthehairpinloopinNewDelhimetallo-β-lactamase,NDM-1,duringcatalysisandinhibitorbinding[J].J Inorg Biochem,2015,156(7):35-39.
[33]Rehman M T,Khan A U.Roleofnon-activesiteresidueTrp-93inthefunctionandstabilityofNewDelhiMetallo-β-Lactamase-1(NDM-1)[J].Antimicrob Agents Chemother,2015,60(1):356-360.
[34]Marcoccia F,Bottoni C,Sabatini A,et al.KineticstudyoflaboratorymutantsofNDM-1metallo-β-lactamase:theimportanceofanisoleucineatposition35[J].Antimicrob Agents Chemother,2016,60(4):2366-2372.
[35]Kim Y,Cunningham M A,Mire J,et al.NDM-1,theultimatepromiscuousenzyme:substraterecognitionandcatalyticmechanism[J].FASEB J,2013,27(5):1917-1927.
[36]Wang Z.Metallo-beta-lactamase:structureandmechanism[J].Curr Opin Chem Biol,1999,3(5):614-622.
[37]Yang H,Aitha M,Marts A R,et al.Spectroscopicandmechanisticstudiesofheterodimetallicformsofmetallo-β-lactamaseNDM-1[J].J Am Chem Soc,2014,136(20):7273-7285.
[38]Yuan Q,He L,Ke H.Apotentialsubstratebindingconformationofβ-lactamsandinsightintothebroadspectrumofNDM-1activity[J].Antimicrob Agents Chemother,2012,56(10):5157-5163.
[39]Chiou J,Leung T Y,Chen S.MolecularmechanismsofsubstraterecognitionandspecificityofNewDelhimetallo-β-lactamase[J].Antimicrob Agents Chemother,2014,58(9):5372-5378.
[40]Zhu K,Lu J,Ye F,et al.Structure-basedcomputationalstudyofthehydrolysisofNewDelhimetallo-β-lactmase-1[J].Biochem Biophys Res Commun,2013,431(1):2-7.
[41]Liang Z,Li L,Wang Y,et al.MolecularBasisofNDM-1,anewantibioticresistancedeterminant[J].PloS One,2010,6(8):e23606.
[42]Zheng M,Xu D.NewDelhimetallo-β-lactamaseI:substratebindingandcatalyticmechanism[J].J Phys Chem B,2013,117(39):11596-11607.
[43]Wang Z,Walter Fast A,Benkovic S J.Onthemechanismofthemetallo-β-lactamasefromBacteroidesfragilisn[J].Biochemistry,1999,38(31):10013-10023.
[44]Yang H,Aitha M,Hetrick A M,et al.Mechanisticandspectroscopicstudiesofmetallo-β-lactamaseNDM-1[J].Biochemistry,2012,51(18):3839-3847.
[45]Zhu K,Lu J,Liang Z,et al.Aquantummechanics/molecularmechanicsstudyonthehydrolysismechanismofNewDelhimetallo-β-lactamase-1[J].J Comput Aid Mol Des,2013,27(3):247-256.
[46]Li N,Xu Y,Xia Q,et al.SimplifiedcaptoprilanaloguesasNDM-1inhibitors[J].Bioorg Med Chem Lett,2014,24(1):386-389.
[47]Shen B,Yu Y,Chen H,et al.Inhibitordiscoveryoffull-lengthnewdelhimetallo-β-lactamase-1(NDM-1)[J].PloS One,2013,8(5):e62955.
[48]Wang X,Lu M,Shi Y,et al.DiscoveryofnovelNewDelhimetallo-β-lactamases-1inhibitorsbymultistepvirtualscreening[J].Plos One,2015,10(3):e0118290.
[49]Rydzik A M,Brem J,van Berkel S S,et al.MonitoringconformationalchangesintheNDM-1metallo-beta-lactamaseby19FNMRspectroscopy[J].Angew Chem Int Edit,2014,53(12):3129-3133.
[50]Klingler F M,Moser D,Büttner D,et al.Probingmetallo-β-lactamaseswithmolecularfragmentsidentifiedbyconsensusdocking[J].Bioorg Med Chem Lett,2015,25(22):5243-5246.
[51]Gan M,Liu Y,Bai Y,et al.Polyketideswithnewdelhimetallo-β-lactamase1inhibitoryactivityfromPenicilliumsp[J].J Nat Prod,2013,76(9):1535-1540.
[52]Christopeit T,Albert A,Leiros H K.Discoveryofanovelcovalentnon-β-lactaminhibitorofthemetallo-β-lactamaseNDM-1[J].Bioorg Med Chem,2016,24(13):2947-2953.
[53]Thomas P W,Spicer T,Cammarata M,et al.Analteredzinc-bindingsiteconfersresistancetoacovalentinactivatorofNewDelhimetallo-beta-lactamase-1(NDM-1)discoveredbyhigh-throughputscreening[J].Bioorg Med Chem,2013,21(11):3138-3146.
[54]Thomas P W,Cammarata M,Brodbelt J S,et al.Covalentinhibitionofnewdelhimetallo-β-lactamase-1(NDM-1)bycefaclor[J].ChemBioChem,2014,15(17):2541-2548.
[55]Chiou J,Wan S,Chan K F,et al.Ebselenasapotentcovalentinhibitorofnewdelhimetallo-β-lactamase(NDM-1)[J].Chem Commun,2015,51(46):9543-9546.
[56]Ma J,McLeod S,MacCormack K,et al.Real-timemonitoringofnewdelhimetallo-β-lactamaseactivityinlivingbacterialcellsby1HNMRspectroscopy[J].Angew Chem Int Edit,2011,126(8):2162-2165.
[57]Azumah R,Dutta J,Somboro A M,et al.Invitroevaluationofmetalchelatorsaspotentialmetallo-β-lactamaseinhibitors[J].J Appl Microbiol,2016,120(4):860-867.
[58]Falconer S B,Reid-Yu S A,King A M,et al.Zincchelationbyasmall-moleculeadjuvantpotentiatesmeropenemactivityinvivoagainstNDM-1-producingklebsiellapneumoniae[J].Acs Infect Dis,2015,1(11):533-543.
[59]Mikami Y,Suzuki T.Novelmicrobialinhibitorsofangiotensin-convertingenzyme,aspergillomarasminesAandB[J].Agric Biol Chem,1983,47(11):2693-2695.
[60]Arai K,Ashikawa N,Nakakita Y,et al.AspergillomarasmineAandB,potentmicrobialinhibitorsofendothelin-convertingenzyme[J].Biosci Biotechnol Biochem,2014,57(11):1944-1945.
[61]Matsuura A,Okumura H,Asakura R,et al.Pharmacologicalprofilesofaspergillomarasminesasendothelinconvertingenzymeinhibitors[J].Jpn J Pharmacol,1993,63(2):187-193.
[62]King A M,Reidyu S A,Wang W,et al.AspergillomarasmineAovercomesmetallo-β-lactamaseantibioticresistance[J].Nature,2014,510(7506):503-506.
[63]Christopeit T,Leiros H S.Fragment-baseddiscoveryofinhibitorscaffoldstargetingthemetallo-β-lactamasesNDM-1andVIM-2[J].Bioorg Med Chem Lett,2016,26(8):1973-1977.
Advance Research on NDM-1:Structure,Catalytic Mechanism and Inhibitors
HUJianping1,2,ZUOKe1,2,WANHua3
(1.School of Pharmacy and Bioengineering,Chengdu University, Chengdu 610106, China;2.Sichuan Industrial Institute of Antibiotics,Chengdu University, Chengdu 610052, China;3.College of Mathematics and Informatics, South China Agricultural University, Guangzhou 510642, China)
As the first-line antibiotics,β-lactam antibiotics are widely used in the clinical treatment of various infections.New Delhi metallo-β-lactamase-1(NDM-1) encoded by gene blaNDM-1in plasmid DNA can hydrolyze nearly all kinds of β-lactam antibiotics,posing a critical threat to the application of β-lactamase.It is of great urgency to design novel NDM-1 inhibitors.The design and modification of inhibitor molecules are closely related to the research of both structural information and catalytic mechanism of NDM-1.In this review,structure,catalytic mechanism and inhibitors of NDM-1 all are summarized,which will be helpful for the design and modification of drug molecules targeting NDM-1.
antimicrobial resistance;NDM-1;catalytic mechanism;inhibitor;drug design
1004-5422(2017)02-0115-10
2017-03-02.
國家自然科學基金(31600591、 11247018)資助項目.
胡建平(1978 — ), 男, 博士, 教授, 從事計算機輔助藥物設計研究.
Q55;R915
A
猜你喜歡
小獼猴智力畫刊(2023年4期)2023-04-23 08:49:58
哲學評論(2021年2期)2021-08-22 01:53:34
中華詩詞(2019年7期)2019-11-25 01:43:04
模具制造(2019年3期)2019-06-06 02:10:54
中學生數理化·高一版(2018年1期)2018-02-10 05:20:03
影視與戲劇評論(2016年0期)2016-11-23 05:26:01
七彩語文·寫字與書法(2016年7期)2016-07-28 21:40:22
七彩語文·寫字與書法(2016年6期)2016-07-15 19:36:34
人間(2015年21期)2015-03-11 15:23:21
現代企業(2015年9期)2015-02-28 18:56:50
