穿山龍抗急性痛風性關節炎的肝臟代謝組學研究
2017-06-19 14:50:43劉樹民張寧于棟華王宇周琦盧芳
中國中藥雜志 2017年10期
劉樹民+張寧+于棟華+王宇+周琦+盧芳

[摘要] 該研究通過肝臟代謝組學研究從肝論治痛風的科學性以及穿山龍提取物對急性痛風性關節炎模型大鼠的防治作用,并找到其相關的潛在生物標志物和相關代謝通路。課題組采用尿酸鈉(MSU)誘導的急性痛風性關節炎模型并運用UPLC-TOF-MS結合模式識別方法探討穿山龍提取物干預急性痛風性關節炎的潛在生物標志物及其相關代謝通路。鑒別出了11個共同的潛在生物標志物,其中穿山龍對正常大鼠的潛在干預靶點中,4個上調,4個下調;穿山龍對急性痛風性關節炎模型大鼠的潛在干預靶點中,5個代謝物在MSU誘導下被上調,5個代謝物被下調,而穿山龍提取物表現出糾正磷酸腺苷、5-甲基四氫葉酸、氧化型谷胱甘肽、次黃嘌呤、二十二碳六烯酸、谷胱甘肽、尿苷二磷酸葡萄糖、肌苷這8個代謝物異常表達的趨勢,其相關性最強的代謝通路是谷胱甘肽代謝、淀粉與蔗糖代謝和嘌呤代謝。故推測穿山龍可以通過干預急性痛風性關節炎狀態下和生理狀態下肝臟中內源性代謝產物變化,實現防治急性痛風性關節炎作用。
[關鍵詞] 急性痛風性關節炎; 穿山龍; 尿酸鈉; 肝臟代謝組學
[Abstract] To explore the prevention and protection effect of Diosocorea nipponica (DNM)) on acute gouty arthritis (AGA) rats based on liver metabonomics, and find potential biomarkers and related pathways. AGA model rats were induced by monosodium urate crystal suspension. UPLC-TOF-MS coupled with pattern recognition technique was employed to find out the potential biomarkers and related metabolic pathways. Eleven common potential biomarkers were identified. Among the potential intervention targets in normal rats given by DNM, 4 biomarkers were up-regulated, and the other 4 targets were down regulated. Among the potential intervention targets in AGA rats given by DNM, 5 metabolites were up-regulated by MSU and 5 metabolites were down regulated. The abnormal expression levels of adenosine monophosphate, 5-methyltetrahydrofolic acid, oxidized glutathione, hypoxanthine, docosahexaenoic acid, glutathione, uridine diphosphate glucose and inosine could be corrected by DNM extract. Three pathways were founded with greatest correlation, including purine metabolism, starch and sucrose metabolism and glutathione metabolism. Therefore, it could be inferred that D. nipponica has the effect for anti-acute gouty arthritis by intervening endogenous metabolites from the liver under physiological condition and acute gouty arthritis condition.
[Key words] acute gouty arthritis; Diosocorea nipponica; monosodium urate; liver metabonomics
《醫學傳燈·痛風》記載:“痛風者,遍身疼痛,晝減夜甚,痛徹筋骨…皆由肝經血少火盛,熱極生風,非是外來風邪…”[1],而現代醫學認為痛風的原因是尿酸含量過高,其根源與肝臟代謝異常有關[2],因此讓痛風從肝臟得到根治是個亟待解決的問題,但從肝論治痛風是否具有科學性仍需要臨床和實驗進一步的檢驗。穿山龍有祛風除濕、舒筋通絡、活血止痛之功,用于各種關節炎[3]。在東北物產豐富,加大對穿山龍的開發,為解除患者疼痛、擴大社會生產力、北藥開發戰略的實施有極其深遠的意義。前期實驗表明穿山龍對高尿酸血癥及痛風性關節炎有良好的治療作用[4-9],基于此運用肝臟代謝組學,預期從整體觀探討穿山龍對生理及急性痛風性關節炎狀態下大鼠的影響,尋找穿山龍作用于肝臟中的潛在靶點,驗證從肝論治痛風的科學性,模擬出穿山龍抗痛風性關節炎的作用通路及其作用機制。
1 材料
QI軟件(Waters 3.0);UPLC-TOF-MS超高液相色譜-飛行時間-質譜儀(美國Waters公司);KDC-160HR高速冷凍離心機(科大創新有限公司中佳分公司);MassLynx V4.1工作站(美國Waters公司)。
乙腈(色譜級,美國Thermo Fisher 科技公司);甲酸(色譜級,美國Dikma 科技公司);亮氨酸腦啡肽(美國Sigma-Aldrich公司);尿酸鹽(美國Sigma-Aldrich公司);蒸餾水(屈臣氏)。黑龍江中醫藥大學中藥毒理學實驗室按本課題組的前期研究方法制得穿山龍提取物,其中偽原薯蕷皂苷、原薯蕷皂苷、甲基原薯蕷皂苷的總含量達到 60%[10]。
SD大鼠,雄性,SPF級,體重(230~280) g,購自黑龍江中醫藥大學實驗動物中心,許可證號SCXK(黑)2013-004,飼養于黑龍江中醫藥大學藥物實驗動物中心屏障系統實驗室。飼養條件:12 h光照,12 h避光循環飼養,室內溫度(22±1) ℃,相對濕度40%~50%,飼喂標準飼料、飲用水。
2 方法
2.1 實驗動物分組及給藥
將SD大鼠40只,隨機分為對照組、穿山龍組、MSU模型組和穿山龍干預組,每組10只。穿山龍組和穿山龍干預組灌胃給予穿山龍提取物,劑量為480 mg·kg-1[11],每天1次,連續5 d。對照組和MSU模型組給予等體積的生理鹽水,每天1次,連續5 d。
用滅菌蒸餾水配成0.025 g·mL-1的MSU混懸液,取0.2 mL一次性踝關節腔注射。第3天灌胃1 h后,將MSU模型組和穿山龍干預組大鼠仰位固定,后肢膝關節周圍剃毛,然后用75%醫用酒精清洗消毒,膝關節輕度彎曲,用6號注射針在大鼠膝關節上方髕上韌帶進針,注射0.2 mL MSU混懸液到關節腔,以關節囊對側鼓起為注入指標,造成實驗性急性痛風關節炎模型。對照組和穿山龍組按上述方法注射等體積生理鹽水。
2.2 肝臟樣本的收集與制備
末次給藥24 h后,將大鼠處死并取出肝臟組織,放入冰冷的生理鹽水中洗去浮血,濾紙吸干后,在冰浴上剪成碎片,稱取適量組織,加入10倍體積冰冷的甲醇進行勻漿,隨后勻漿液于13 000 r·min-1,4 ℃離心15 min,上清液轉移至另一離心管后再重復該條件離心1次,最后的上清液儲存于-80 ℃中。
2.3 代謝組學樣本分析方法的建立
2.3.1 UPLC條件 ACQUITY UPLC BEH C18色譜柱(2.1 mm× 50 mm,1.7 μm);進樣量3 μL;流動相0.1%甲酸乙腈(A)-0.1%甲酸水溶液(B),洗脫梯度,0~2.5 min,1%~10% A,2.5~3.5 min,10%~40% A,3.5~11 min,40%~70% A,11~12 min,70%~80% A,12~13 min,80%~100% A,流速0.4 mL·min-1;柱溫40 ℃。
2.3.2 MS條件 電噴霧離子源(ESI),采用正、負離子W模式檢測,鎖定質量溶液,采用美國Waters公司LockSprayTM校正系統進行在線質量校正。亮氨酸腦啡肽準分子離子峰m/z 556.277 1[M+H]+;m/z 554.261 5[M-H]-,鎖定質量濃度為1 μg·L-1,流速為30 μL·min-1。掃描方式為全掃描,質量掃描范圍為質荷比m/z 100~1 500。其質譜條件見表1。
2.4 多元數據分析及潛在干預靶點的鑒定
UPLC-TOF-MS系統為不同離子所對應的質譜峰提供了精確的相對分子質量數據,這些數據被首先用于幫助確定代謝物的相對分子質量。對肝臟樣品進行檢測,將數據導入QI 軟件[12],聯合MarkerLynx XS軟件中Ezinfo 2.0軟件對大鼠肝臟中代謝物組進行主成分分析(principal components analysis,PCA),繪制反映組間離散度的Scores plot,觀察肝臟代謝物軌跡是否在各自組內相似度較好,在同一判別標準下不同的組別是否呈現各自聚類現象。結合各組間各離子點量的變化趨勢,在S-plot圖中選擇潛在的生物標志物,以變量投射重要性(VIP)>1的離子作為生物標志物,根據鑒定結果聯合HMDB(人類代謝組數據庫)及KEGG(Kyoto encyclopedia of genes and genomes,京都基因與基因組百科全書)等數據庫及大量文獻對生物標志物進行生物學意義的解釋,最終實現對數據的分析確定和代謝通路的分析。
3 結果
3.1 大鼠肝臟代謝數據分析
為了獲得大鼠肝臟樣本代謝輪廓的概況,3D-PCA被運用于隨后的UPLC-TOF-MS數據分析中。結果顯示空白組和穿山龍組明顯分離,說明空白組和穿山龍組出現了明顯的化學擾動,見圖1。MSU模型組的代謝輪廓遠離了空白組,表明MSU誘導后大鼠肝臟內發生了顯著地生物學變化,見圖2,3。穿山龍提取物干預組的代謝輪廓遠離于MSU模型組,有向空白組回調趨勢,說明由MSU誘導產生的偏離得到了明顯改善。
在S-plots圖中,絕大多數離子聚集于原點附近,只有少數離子偏離原點,即S-plots圖兩端的離子造成了2組組間的差異,見圖4。并選定VIP>1的離子預選為潛在的生物標志物,見圖5。
3.2 穿山龍對正常大鼠和急性痛風性關節炎模型大鼠潛在生物標志物
不同生理病理狀態可能存在相同的代謝產物譜,這可能是機體應激出現的結果,也可能是藥物作用的潛在靶點,因此通過分析發生變化的共同代謝產物有助于推測疾病的發病機制以及藥物對疾病的預防和治療作用。穿山龍對正常和急性痛風性關節炎模型大鼠的代謝變化比較,得到11個相同的生物標志物,見表2。
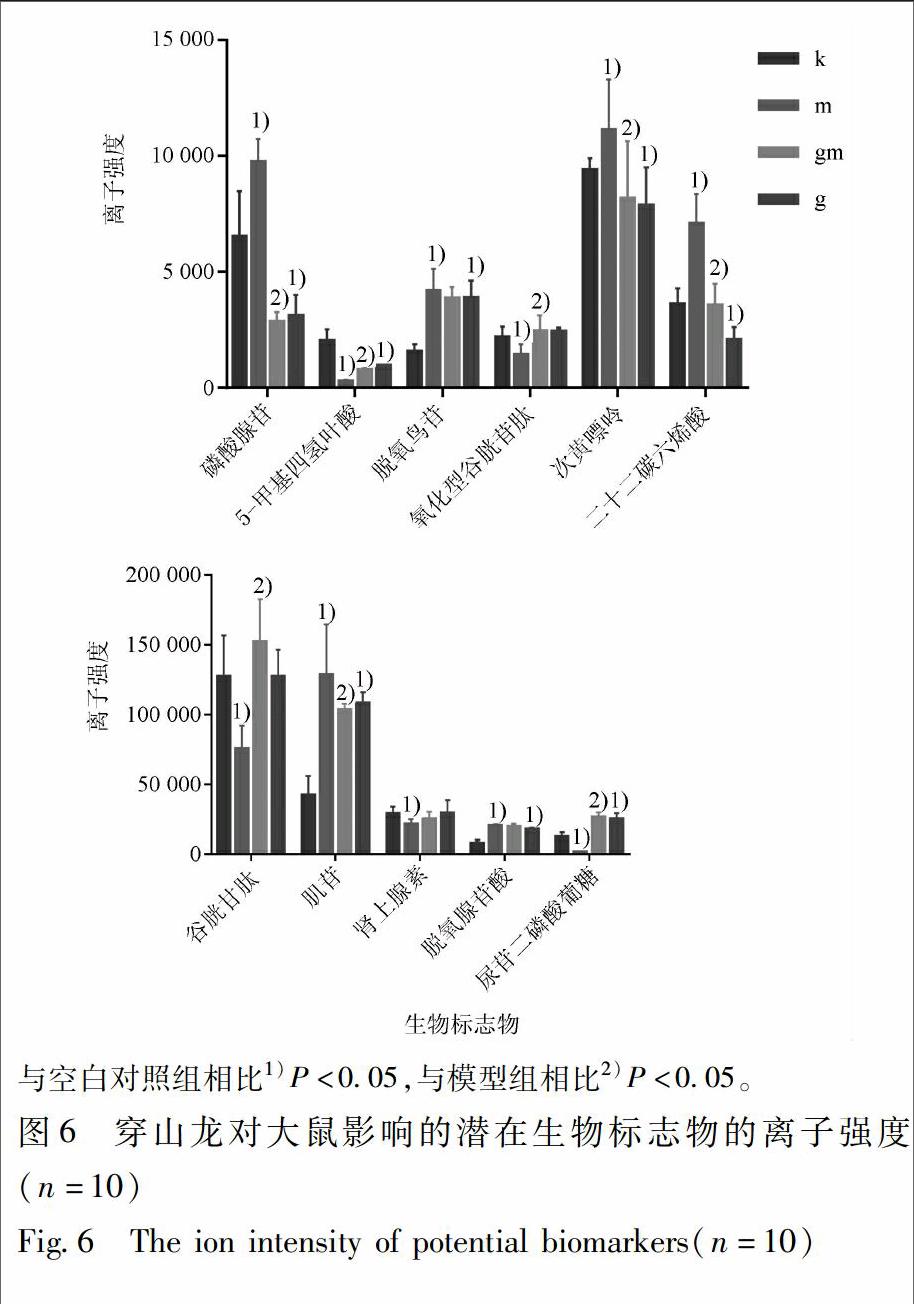
通過比對這些潛在生物標志物的離子強度,其中穿山龍對正常大鼠的潛在干預靶點中,4個上調,4個下調;穿山龍對急性痛風性關節炎模型大鼠的潛在干預靶點中,5個代謝物在MSU誘導下被上調,5個代謝物被下調,而穿山龍提取物表現出糾正磷酸腺苷、5-甲基四氫葉酸、氧化型谷胱甘肽、次黃嘌呤、二十二碳六烯酸、谷胱甘肽、尿苷二磷酸葡萄糖、肌苷這8個代謝物異常表達的趨勢,見圖6。
3.3 肝臟中潛在生物標志物的分類
使用HMDB對潛在生物標志物進行分類,11個潛在靶點中,大約9%屬于脂類,46%為核苷、核苷酸及它們的類似物;18%為有機酸及其衍生物,9%為兒茶酚胺類及其衍生物,18%為雜環化合物。在生物功能分布中,所調節的潛在靶點中主要參與的生物過程有嘌呤代謝(9%)、谷氨酸代謝(4%)、蛋氨酸代謝組分(5%)、谷胱甘肽代謝組分(7%)、廢物(11%)和其他(64%)。另外,這些代謝物主要存在于細胞外(29%)、細胞質(21%)、溶酶體(14%)和線粒體(14%)等部位。
3.4 肝臟中潛在生物標志物的通路分析
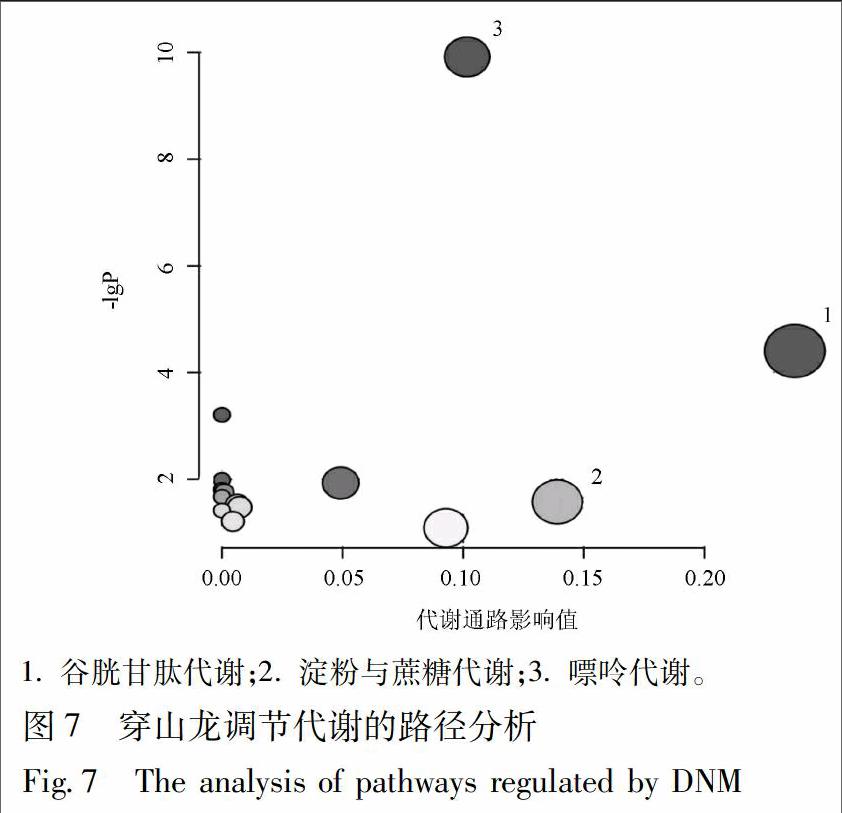
將11個共同的潛在生物標志物作為穿山龍抗急性痛風性關節炎的肝臟潛在生物標志物,并進行MetaPA識別顯著富集通路分析,確定與所分析代謝物組相關性最強的代謝通路。代謝通路影響值的臨界值設置為0.10,大于這個值,將選擇作為潛在關鍵代謝通路,見圖7,表3。主要富集于谷胱甘肽代謝、淀粉與蔗糖代謝和嘌呤代謝。在穿山龍干預后,代謝物的含量得到回調,代謝通路的紊亂得到改善,在代謝途徑層面驗證了穿山龍對由MSU誘導的急性痛風性關節炎模型有良好的預防和保護作用。
4 結論與討論
4.1 谷胱甘肽代謝
谷胱甘肽(GSH)和氧化型谷胱甘肽(GSSG)參與了谷胱甘肽代謝。在活細胞內,2分子GSH可脫氫以二硫鍵相連形成GSSG。GSSG在NADPH+和H+存在下,由谷胱甘肽還原酶(glutathione reductase,GR)催化,又可還原成GSH。還參與機體三羧酸循環及糖代謝,使機體獲得高能量,能激活各種酶,從而促進糖、脂肪、蛋白質代謝,也能影響細胞的代謝過程,改善中毒性肝炎、感染性肝炎的癥狀[13],炎癥免疫與急性痛風性關節炎密切相關[14],且前期研究證實穿山龍總皂苷抑制急性痛風性關節炎模型大鼠產生IL-1β 和 TNF-α 這類致炎因子,阻斷其所介導和激發的炎癥反應過程[11] 。此外,也能抑制脂肪肝的形成[15],脂肪肝與血脂和血尿酸的關系是并存的,且互為影響,脂肪肝形成的過程中尿酸的合成增加,使患者亦并發痛風[16]。在此研究中,MSU模型組中GSH和GSSG的濃度是明顯減少的(P<0.05),且GSH的降低幅度高于GSSG,可以推測出MSU主要抑制了谷胱甘肽還原酶,致GSH的濃度在肝臟中的含量減少,而穿山龍提取物的干預有利于肝臟中GSH和GSSG轉化的進行。穿山龍提取物的防治作用可能是通過調節谷胱甘肽代謝中谷胱甘肽還原酶的活性,進而調節GSH和GSSG的水平來實現的,見圖8。
4.2 淀粉與蔗糖代謝
尿苷二磷酸葡萄糖參與了淀粉與蔗糖代謝,為生物體合成雙糖及多糖的中間物[17]。肝臟中存在的動物性多糖為肝糖原,其在機體內的終產物為葡萄糖,為體內代謝供能。此研究中,MSU模型中尿苷二磷酸葡萄糖的含量是明顯減少的(P<0.05),這表明MSU可以間接的減少機體內肝糖原的含量,而穿山龍提取物可顯著增加尿苷二磷酸葡萄糖的含量(P<0.05),間接的增加機體內肝糖原的貯備量,提高機體的運動耐力[18]。因此穿山龍提取物可以通過間接增加機體內肝糖原的貯備量來改善MSU模型急性痛風性關節炎患者運動不便的困境。
4.3 嘌呤代謝
脫氧鳥苷(dG)、肌苷、次黃嘌呤、脫氧腺苷酸(dAMP)和磷酸腺苷(AMP)分別參與了嘌呤代謝。肌苷酸和腺苷是肌苷的直接前體。dG是鳥苷一磷酸的直接前體,可轉化成肌苷酸,而肌苷是其部分水解的產物;腺苷是由dAMP間接轉換而來,肌苷由AMP在腺苷脫氨酶催化下生成,在有關酶作用下可形成次黃嘌呤,它是核酸中嘌呤堿基的代謝中間物。經黃嘌呤氧化酶催化作用可形成黃嘌呤,進一步代謝形成尿酸[19]。痛風患者一般都有尿酸代謝紊亂,出現尿酸高的現象[20],在此研究中,MSU模型中的dG,肌苷,次黃嘌呤,dAMP和AMP的濃度均明顯增加(P<0.05),可以推測MSU促進并誘導了機體內尿酸的合成,而穿山龍提取物的干預均抑制尿酸的合成,推測穿山龍提取物的防治作用可能是通過調節嘌呤代謝中dG,肌苷,次黃嘌呤,dAMP和AMP與尿酸的水平來實現的。
本研究共發現了11個共同的潛在生物標志物,分別為磷酸腺苷、腎上腺素、次黃嘌呤、5-甲基四氫葉酸、谷胱甘肽、脫氧鳥苷、氧化型谷胱甘肽、肌苷、二十二碳六烯酸、脫氧腺苷酸、尿苷二磷酸葡糖,相關性最強的通路有谷胱甘肽代謝、淀粉與蔗糖代謝和嘌呤代謝。值得注意的是穿山龍提取物對dG、肌苷、次黃嘌呤、dAMP和AMP的下調和尿苷二磷酸葡萄糖、GSH和GSSG的上調均具有顯著的逆轉趨勢,能夠使得它們的表達恢復平衡。這些結果可能表明了穿山龍提取物對MSU誘導的急性痛風性關節炎的防治作用,可以通過多個代謝物靶點的干預來實現。MSU介導的急性痛風性關節炎模型大鼠以及生理狀態下穿山龍組大鼠體內發生了內源性代謝物的擾動。鑒別的潛在生物標志物參與了急性痛風性關節炎相關的代謝通路,而穿山龍提取物對相關靶點具有干預作用,從而找到穿山龍防治急性痛風性關節炎的作用機制。
課題組的創新之處在于通過肝臟代謝組學間接驗證了從肝論治痛風的科學性,并探討了由MSU誘導的急性痛風性關節炎模型和生理狀態給予穿山龍后共同的潛在生物標志物及其相關性最強的代謝通路,力求尋找穿山龍可干預的與生理病理(急性痛風性關節炎)變化的小分子代謝物,同時監測其整個代謝網絡的變化,有助于表征急性痛風性關節炎的病理狀態,并為穿山龍的預防和治療急性痛風性關節炎提供診斷信息。同時,為疾病和藥物的生物化學作用機制提供新的見解。肝臟代謝組學在中醫藥研究中有利于理解急性痛風性關節炎這種疾病的本質及評估穿山龍的防治作用。
[參考文獻]
[1] 朱婉華, 張愛紅, 顧冬梅, 等. 痛風性關節炎中醫證候分布規律探討[J]. 中醫雜志,2012,53(19):1667.
[2] Irondi E A, Oboh G, Agboola S O, et al. Phenolics extract of Tetrapleura tetraptera fruit inhibits xanthine oxidase and Fe2+-induced lipid peroxidation in the kidney, liver, and lungs tissues of rats in vitro[J]. Food Sci Hum Well, 2016, 5(1): 17.
[3] 張寧, 于棟華, 周琦, 等. 穿山龍藥理作用的研究進展[J]. 中國藥房, 2015,26(4):547.
[4] 周琦, 張翀, 于棟華, 等. 穿山龍總皂苷對高尿酸血癥的降尿酸及細胞抗炎作用研究[J]. 中華中醫藥雜志, 2013, 28(5):1444.
[5] 姚麗, 劉樹民. 穿山龍治療急性痛風性關節炎有效部位的實驗研究[J]. 中華中醫藥學刊, 2010, 28(8):1724.
[6] 周琦, 張寧, 盧芳, 等. 穿山龍總皂苷對痛風性關節炎大鼠關節炎滑膜IL-1β及其信號轉導通路的影響[J]. 中藥藥理與臨床, 2013, 29(6):52.
[7] 周琦, 張寧, 盧芳, 等. 穿山龍總皂苷對白介素-1β誘導的成纖維樣滑膜細胞基質細胞衍生因子-1及IκB激酶表達的影響[J]. 中國中西醫結合雜志, 2015, 35(2):234.
[8] 盧芳, 周琦, 張穎, 等. 基于痛風性關節炎ABCG2尿酸轉運靶點的穿山龍調控機制研究[J]. 中華中醫藥學刊, 2016, 34(5):1057.
[9] Zhou Q, Liu S M, Yu D H, et al. Therapeutic effect of total saponins from Dioscorea nipponica Makino on gouty arthritis based on the NF-κB signal pathway: an in vitro study[J]. Pharmacogn Mag, 2016, 12(47):235.
[10] 劉穎,劉樹民,于棟華, 等. 大孔樹脂純化穿山龍總皂苷的工藝優化[J]. 醫藥導報,2011,30(5):635.
[11] 劉樹民,張寧,周琦, 等. 藥理效應法測定穿山龍總皂苷的藥動學參數[J]. 中國實驗方劑學雜志,2016,22(16):75.
[12] Zhang J, Yang W, Li S, et al. An intelligentized strategy for endogenous small molecules characterization and quality evaluation of earthworm from two geographic origins by ultra-high performance HILIC/QTOF MS(E) and Progenesis QI[J]. Anal Bioanal Chem, 2016, 408(14): 3881.
[13] Ibáez-Samaniego L, Salcedo M, Vaquero J, et al. De novo autoimmune hepatitis after liver transplantation: a focus on glutathione S-transferase theta 1[J]. Liver Transpl, 2017, 23(1): 75.
[14] Zhang N, Gong X, Lu M, et al. Crystal structures of Apo and GMP bound hypoxanthine-guanine phosphoribosyltransferase from Legionella pneumophila and the implications in gouty arthritis[J]. J Struct Biol, 2016, 194(3): 311.
[15] Misako T, Nobuko M, Chizuru I, et al. Elevation of glucose 6-phosphate dehydrogenase activity induced by amplified insulin response in low glutathione levels in rat liver[J]. Scientific World J, 2016, 2016:1.
[16] Johnson R J. Why focus on uric acid[J]. Curr Med Res Opin, 2015, 31(sup2):3.
[17] Schmlzer, Katharina, Lemmerer, et al. Integrated process design for biocatalytic synthesis by a Leloir glycosyltransferase: UDP-glucose production with sucrose synthase[J]. Biotechnol Bioeng, 2016,doi: 10.1002/bit.26204.
[18] Adeva-Andany M M, Pérez-Felpete N, Fernández-Fernández C, et al. Liver glucose metabolism in humans[J]. Bioscience Rep, 2016, 36(6): e00416.
[19] Maiuolo J, Oppedisano F, Gratteri S, et al. Regulation of uric acid metabolism and excretion[J]. Int J Cardiol, 2016, 213: 8.
[20] Saigal R, Agrawal A. Pathogenesis and clinical management of gouty arthritis[J]. J Assoc Physicians India, 2015, 63(12):56.
[責任編輯 曹陽陽]
