膜下滴灌水稻基因型耐缺鐵性評價
2017-06-13 11:05:47李言言白如霄張新疆楊玉珍黃致華侯建偉危常州
植物營養與肥料學報 2017年3期
李言言,白如霄,張新疆,楊玉珍,黃致華,侯建偉,危常州
膜下滴灌水稻基因型耐缺鐵性評價
李言言,白如霄,張新疆,楊玉珍,黃致華,侯建偉,危常州*
(石河子大學農學院,新疆石河子 832003)
【目的】新疆生產建設兵團采用膜下滴灌技術后,水稻缺鐵黃化現象較為嚴重,研究該條件下水稻對缺鐵反應的基因型差異,可為耐缺鐵性水稻基因型的篩選和分類提供可靠的理論依據。 【方法】以六個水稻基因型為研究材料,采用膜下滴灌技術管理。在水稻幼苗期、分蘗期和成熟期隨機采樣,測定不同基因型水稻的分蘗數、生物量、產量以及鐵含量,利用隸屬函數分析和聚類分析研究了水稻在全生育期對缺鐵脅迫反應的基因型差異。 【結果】水稻在幼苗期、分蘗期和成熟期對缺鐵的反應存在基因型差異。在幼苗期水稻基因型 T-04和 T-05 的葉片活性鐵含量顯著低于其他基因型;T-201 在幼苗期葉片活性鐵、地上部鐵的分配,分蘗期葉片鐵含量和地上部干物質均較高;在分蘗期 T-04 鐵的轉移能力最小,但是 T-04 的分蘗數較高;在成熟期 T-04的有效分蘗與其他基因型差異不顯著,T-04 的產量處于中等水平,但是其籽粒鐵的收獲指數低于其他基因型水稻。聚類分析顯示耐缺鐵水稻基因型 T-43 的各指標高于其他基因型。 【結論】在水稻的幼苗期和分蘗期葉片鐵的有效利用和自身鐵的轉移保證了水稻的生長和較高有效穗數,根據全生育期水稻鐵營養效率和產量的基因型差異初步確定 T-43 耐缺鐵能力較強,T-04 為對缺鐵敏感的水稻基因型。
水稻;膜下滴灌;基因型;耐缺鐵性
水稻是耗水較多的農作物之一,超過 43% 的農業灌溉用水被用于水稻生產[1],而我國又是世界上水資源最短缺的國家之一[2],因此節水灌溉是我國稻作的理性選擇,灌溉技術的發展勢在必行[3]。近年來,新疆建設兵團研發出水稻膜下滴灌技術,與水稻傳統種植方式相比,該技術節水 60% 以上[4–5]。
雖然膜下滴灌水稻種植技術是一項節水高產的水稻栽培模式,但是在幼苗期經常出現“黃化失綠”現象,嚴重時導致水稻幼苗死亡[6–7]。石灰性土壤上含較高的 HCO3–,pH 值高,氧化還原電位均是造成水稻黃化的原因[8–10]。節水旱作時,水稻缺鐵黃化現象較為普遍[11–12],鐵素營養失調已成為制約膜下滴灌水稻高產、穩產的限制因子。Brown[13]指出篩選適應土壤環境的植物較改變土壤環境去滿足植物需求更實際可行。篩選和利用鐵高效作物基因型是解決作物缺鐵黃化有效途徑之一。且篩選鐵高效的作物品種已經在許多作物上實現[14–17]。但關于膜下滴灌水稻對缺鐵脅迫的基因型差異還沒有相關研究。
目前,通過基因工程培育耐缺鐵性水稻基因型的研究已經取得成果[18–20],但是在田間耕作時,水稻耐缺鐵能力受土壤環境的綜合影響,推廣起來存在困難。田間試驗下選擇適宜的、方便可靠的鑒定指標進行耐缺鐵能力的評定依然是獲得耐缺鐵水稻基因型的主要手段[21–23]。鐵在植物體內的利用涉及多個生理生化過程,從單個生育期或者單個指標分析植物鐵營養狀況存在局限性。大多數關于水稻耐缺鐵能力的研究是在幼苗期進行。Zeng 等[24]研究發現,水稻在鹽脅迫下的分蘗數存在顯著的基因型差異,這種差異是造成水稻產量不一的重要原因。而關于缺鐵脅迫下水稻分蘗數、籽粒收獲指數和產量等農藝性狀的基因型差異研究較少。雖然缺鐵脅迫下水稻鐵營養吸收、利用和轉運能力存在顯著差異,但是其基因型差異與農業生產的實際沒有很好的結合。在缺鐵脅迫土壤上進行水稻全生育期培養,將水稻田間農藝性狀與鐵的吸收、轉移能力相結合評價水稻耐缺鐵能力的基因型十分有必要。本研究通過田間試驗,利用隸屬函數值、聚類分析方法進行聚類分析排名,對膜下滴灌模式下種植的 6 個水稻品種進行鐵營養和農藝性狀評價,篩選出適合膜下滴灌種植的抗黃化品種,從而為耐缺鐵脅迫的機理研究及鐵高效品種的選育提供基礎材料。
1 材料與方法
1.1 土壤性質和供試品種
試驗于 2015 年 4 月中旬至 9 月中旬在新疆天業農業研究所試驗田進行。土壤類型為灰漠土,土壤質地為輕壤土,歷年種植經驗確認該試驗地點水稻缺鐵黃化現象比較嚴重。土壤有機質含量 29.6 g/kg,全氮含量 2.47 g/kg,堿解氮 61.72 mg/kg,Olsen-P 為67.29 mg/kg,速效鉀 (NH4OAc-K) 282 mg/kg,有效鐵 (Fe-DTPA) 為 4.15 mg/kg,pH 為 8.43 (土水比1∶2.5),CaCO3含量 35.9 g/kg。本試驗采用的 6 個水稻基因型均取自新疆天業研究所,是從 200 多個品種 (系) 篩選的,且黃化差異較大的品種。
1.2 試驗設計與田間管理
本試驗為單因素試驗,3 次重復,隨機區組排列,每個小區面積為 17 m2,共 18 個小區。4 月 29日播種,種子播深 2~3 cm,干播濕出。每穴播種10 粒,畝穴數 1.9 萬穴。全生育期灌水量為 10500 m3/hm2,肥料分別選用尿素、磷酸二銨和硫酸鉀。各處理的氮、磷、鉀施肥量相同,分別為 N 300 kg/hm2、P2O5270 kg/hm2、K2O 240 kg/hm2。施肥方法是將肥料溶于肥料罐中,采用水—肥—水的方式通過滴灌帶將肥料施于土壤中。分別在水稻幼苗期、分蘗期和成熟期隨機采樣,測定葉片葉綠素含量,葉片活性鐵含量,全鐵含量,干物質等指標。水稻成熟期完成測產和其他采樣工作后,用谷物聯合收割機收獲。
1.3 測定項目與方法
1.3.1 干物質 在水稻幼苗期 (出苗后 28 d)、分蘗期(出苗后 53 d) 和成熟期 (出苗后 156 d) 進行取樣。在幼苗期用不銹鋼剪刀將植株分成地上部和根部,分蘗期將植株分為根、莖和葉,成熟期分為根、莖、葉和籽粒,地上部各組織用 0.1 mol/L HCl 清洗 0.5 h,再用蒸餾水清洗數次,根部用飽和 CaSO2溶液浸泡0.5 h,再用蒸餾水清洗數次后分別裝入牛皮紙袋中,于 105℃ 下殺青 30 min,75℃ 烘至恒重。測定各器官干物質后,粉碎待用。
1.3.2 葉片 SPAD 值 于出苗后第 28 天,用 SPAD-205 葉綠素儀 (日本柯尼卡美能達公司) 測定完全展開新葉 SPAD 值,每小區取連續 10 穴,計算平均值作為該試驗小區 SPAD 值。
1.3.3 植株活性鐵和全鐵含量 出苗后 28 天隨機選取具有代表性的連續 5 穴水稻,分離幼葉,將幼葉用不銹鋼剪刀剪成 2~3 mm 大小的碎片,稱取 2.0000 g碎葉放于塑料瓶中,加入 20 mL 1 mol/L 鹽酸,靜置24 h 后,過濾,濾液用 AAS 法測定葉片中活性鐵含量。另外,稱取一定量粉碎后的各組織樣品,在 550℃馬福爐干灰化 7~8 h,灰分用1∶30 HNO3溶解后,用 AAS 法測定全鐵含量。
1.3.4 考種 每個小區選取 1 m2的水稻進行測產考種,分別記錄水稻有效穗數、穗粒數、實粒數和千粒重。
1.4 數據處理
采用 SAS 8.0 統計軟件包進行數據分析,水稻基因型間差異顯著性用 LSD 法檢驗 (P < 0.05)。采用Excel 對各性狀進行隸屬函數度的計算。采用 SPSS分析程序進行 Ward 法 Squared Euclidean distance (歐氏距離平方) 聚類分析。
隸屬函數度的計算 X (u) = (X – Xmin)/(Xmax– Xmin)式中:X 為作物各基因型的某一指標測定值,Xmax為所試基因型中某一指標測定值的最大值,Xmin為該指標中最小值。
地上部鐵的分配 = (地上部鐵的含量 × 地上部生物量)/(根部鐵的含量 × 根部生物量 + 地上部鐵的含量 × 地上部生物量) × 100%。
2 結果與分析
2.1 幼苗期水稻耐缺鐵性的基因型差異
由表 1 可知,6 個基因型的葉片 SPAD 值變幅為32.98~37.32,新葉活性鐵含量范圍為 30.15~39.47 μg/g,地上部鐵的分配在 52.70%~74.61% 之間,生物量變異系數為 17.2%,水稻耐缺鐵脅迫能力存在明顯的基因型差異。
基因型 T-X 葉片 SPAD 值顯著高于其他基因型。T-05 和 T04 葉片 SPAD 值較低,活性鐵含量也最低,與其他基因型水稻差異顯著。但是 T-04 地上部鐵分配比率較 T-X 高。T-69 的生物量較小,其次為 T-04,而 T-43 的各項指標都相對較高。可見,篩選耐缺鐵性的水稻品種應該綜合考慮相關指標。
2.2 分蘗期水稻耐缺鐵性的基因型差異
分蘗期水稻葉片鐵含量、鐵的轉運和地上部干物質在基因型間存在差異顯著 (表 2)。6 個水稻品種葉片鐵含量最小值為 70.1 μg/g,最大值為 126.5 μg/g;地上部干物質變幅在 1269.1~3147.6 kg/hm2之間,地上部鐵分配比率為 48.9%~74.6%,各項指標變異系數在 13.4%~36.5% 之間。
水稻 T-201、T-X 和 T-43 地上部干物質較高,與 T-69、T-05 和 T-04 差異顯著,后三者之間無顯著差異,表明缺鐵脅迫下水稻在分蘗期的生長狀況存在差異。水稻葉片鐵含量 T-201 和 T-43 最高,顯著高于 T-X 和 T-04,T-04 葉片鐵含量較低,僅為 T-43的 60.5%。不同基因型水稻地上部鐵的分配率差異顯著,T-43 和 T-X 鐵的轉運能力顯著高于其他基因型水稻,其中 T-04 鐵的轉運能力較小。與其他基因型相比,在分蘗期 T-201 和 T-43 的耐缺鐵脅迫能力較高,而 T-05 和 T-04 的各項指標均較低,所以膜下滴灌栽培模式種植水稻時要綜合考慮。
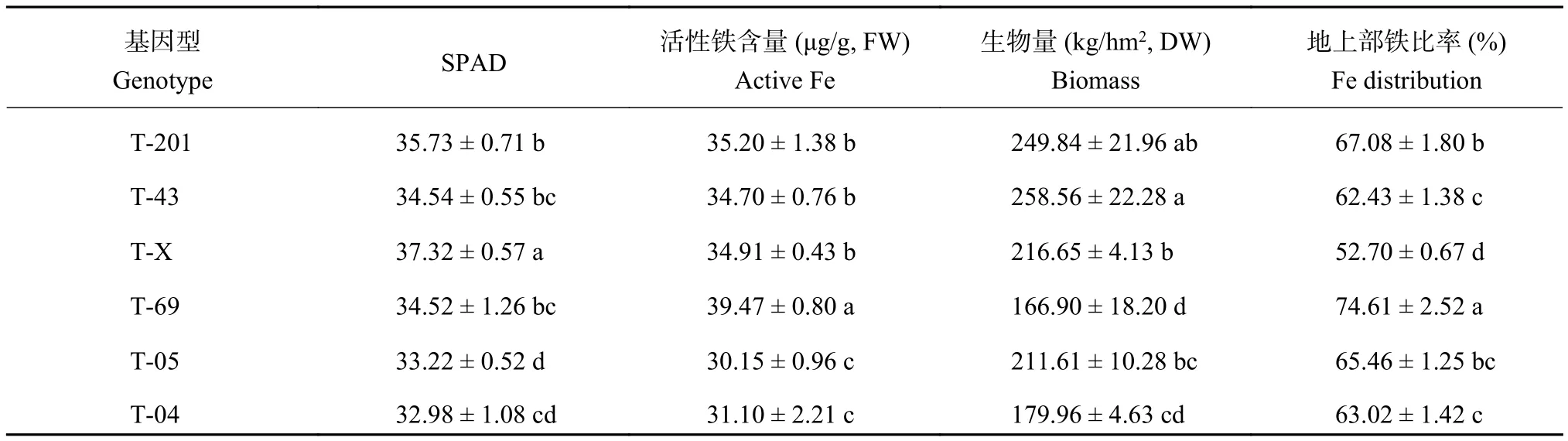
表1 水稻幼苗期葉片 SPAD 值、活性鐵含量、生物量和地上部鐵的分配Table1 SPAD value, plant leaf active iron content, dry biomass and Fe distribution in rice seedling shoot
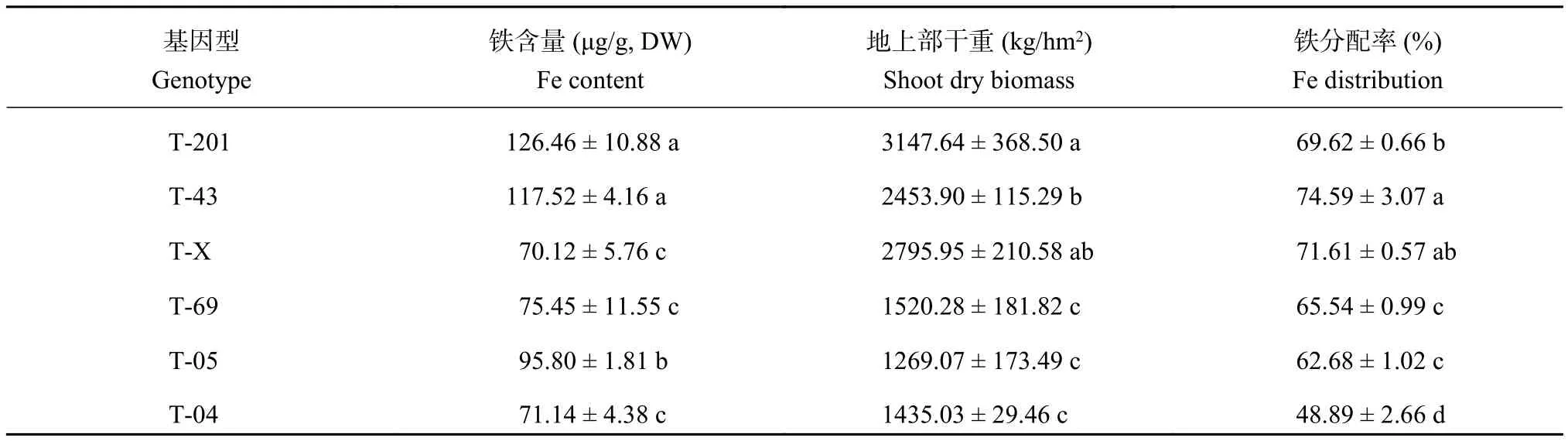
表2 水稻分蘗期葉片鐵含量、地上部干物質量、地上部鐵的分配率Table2 Leaf Fe content, shoot dry biomass, Fe distribution in shoot of selected rice plants at tillering stage
2.3 不同基因型水稻莖蘗動態差異
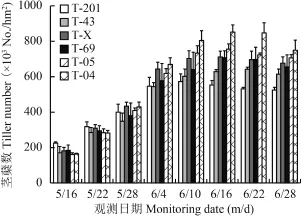
圖1 不同基因型水稻分蘗動態Fig. 1 Number of tillers of different rice genotypes at investigation date
圖1 表明,在水稻分蘗盛期 (6 月 16 日和 6 月22 日),莖蘗的變異系數分別為 14.7% 和 15.1%。由此可見水稻的分蘗數存在顯著的基因型差異,而分蘗數反映了水稻的生長狀況,且與產量密切相關,評價水稻耐缺鐵性時分蘗數被視為重要指標。
5 月 16 日 T-201 的分蘗數顯著高于其他基因型水稻,但是 6 月 4 日之后 T-201 的分蘗數均顯著低于其他基因型水稻,由此看出,T-201 的分蘗能力偏弱,T-04 的分蘗數卻明顯高于其他品種水稻。6 月28 日,各基因型水稻的分蘗數較前期均有下降,但是 T-04 的分蘗數降低了 98 × 104株/hm2,降低程度較其他基因型明顯,可見,T-04 受缺鐵脅迫的影響,其分蘗死亡率較高,耐缺鐵脅迫的能力較小。在 6 個基因型間,T-43 的分蘗數偏低,為 615 × 104株/hm2。
2.4 水稻產量和產量構成因子的基因型差異
表3 結果顯示, T-201 有效穗數、結實率和千粒重均顯著低于其他基因型水稻,其他基因型水稻的有效穗數無顯著差異,T-43 和 T-69 的結實率顯著高于其他基因型水稻,T-201 和 T-04 的穗粒數較高。T-201 的產量最低,其次是 T-X,而 T-43 的產量顯著高于其他基因型水稻的產量。T-04 的有效穗數與 T-43、T-69 差異不顯著,但是其產量卻低于二者的產量,對 T-04 而言,在育種工作中,提高籽粒的結實率是提高 T-04 產量的途徑之一。T-69 和 T-43的結實率顯著高于其他基因型,但是二者的穗粒數較低,提高二者的穗粒數是提高產量的主要途徑。在所測 6 個基因型水稻中,T-05 產量構成因子和產量屬于中等水平。缺鐵脅迫下,不同基因型水稻的產量受不同的產量因子的影響,提高產量需要結合各基因型不同的特征,其中提高 T-43 穗粒數可以進一步提高其產量。
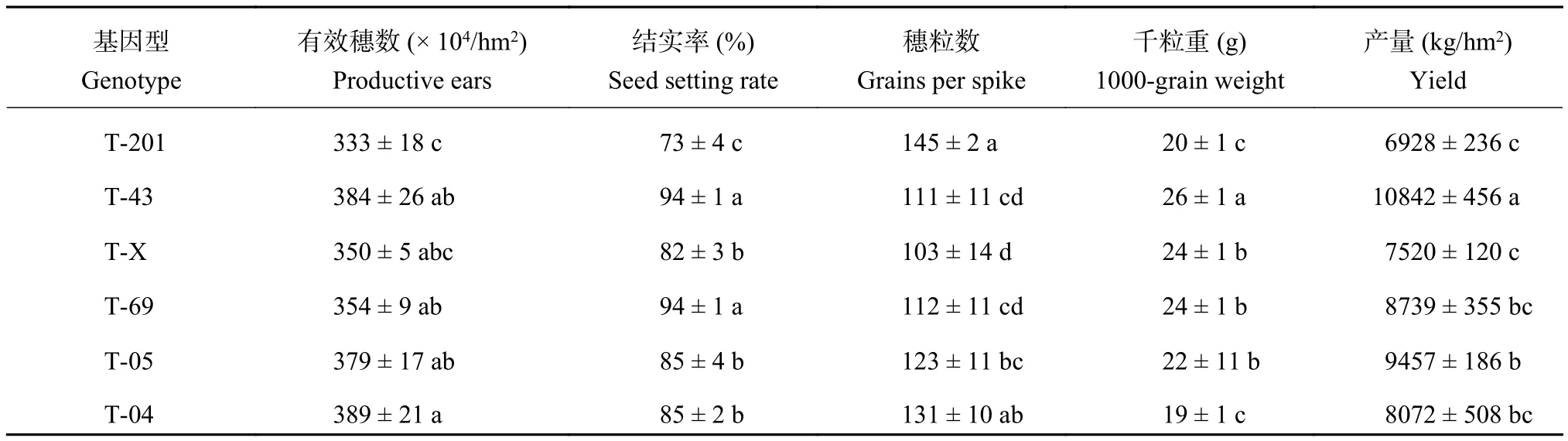
表3 各基因型水稻產量和產量構成Table3 Yields and yield components of the selected rice genotypes
除去 T-201 以外,T-04 的分蘗數顯著高于其他品種 (圖 1),但有效穗數與其他基因型差異不顯著(表 3),說明 T-04 的無效分蘗較多。T-43 的分蘗數雖較其他基因型低 (圖 1),但其有效穗數較高 (表 3),即有效分蘗率相對較高。
2.5 水稻籽粒收獲指數和鐵收獲指數的基因型差異
圖2 表明,T-05 籽粒收獲指數最高,顯著高于其他基因型水稻,其次為 T-69 和 T-43,T-201 水稻籽粒收獲指數最小,僅為 33.0%,并顯著低于其他基因型水稻。T-69 和 T-43 水稻鐵收獲指數也較高,分別為 25.7% 和 23.8%,T-04 鐵收獲指數最低,僅為12.4%。由此可見,成熟期水稻組織器官的鐵向籽粒的轉運和干物質在籽粒的分配均存在顯著的基因型差異。
2.6 不同基因型水稻各指標隸屬函數值
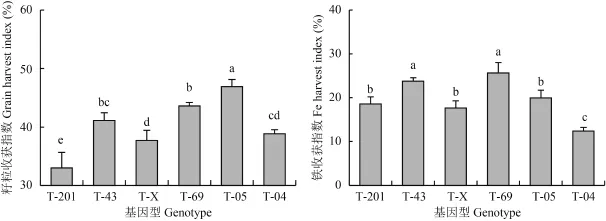
圖2 不同基因型水稻籽粒收獲指數和鐵鐵收獲指數Fig. 2 Grain harvest indices and Fe harvest indices of different rice genotypes[注(Notes):籽粒收獲指數 = 籽粒產量/(籽粒產量 + 秸稈干物質) × 100%;鐵收獲指數 = 籽粒鐵吸收量/(籽粒鐵吸收量 + 秸稈鐵吸收量) × 100%. Grain harvest index = grain yield/(grain yield + straw dry weight) × 100%;Fe harvest index = grain Fe uptake/(grain Fe uptake + straw Fe uptake) × 100%.]
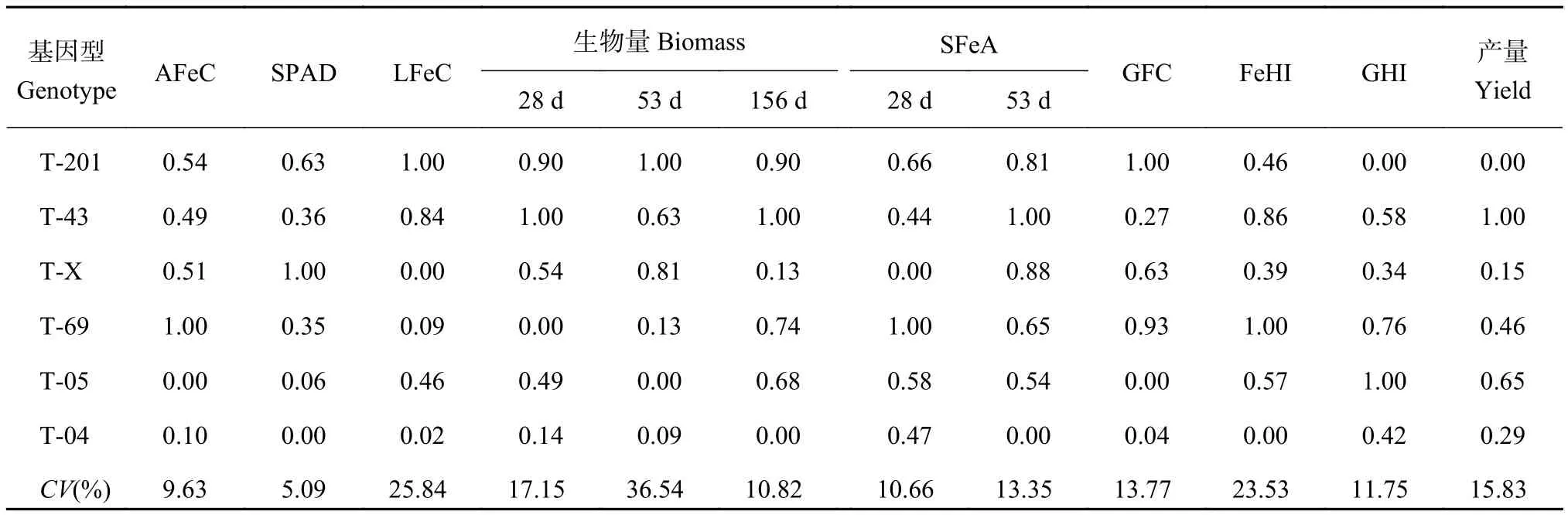
表4 不同基因型水稻各指標的隸屬函數值Table4 Subordinate function values of tolerating iron deficiency indexes in rice
由表 4 可知,對于同一指標如葉片活性鐵含量,T-69 的隸屬函數值最大,為 1,表明 T-69 在這一指標上表現為耐缺鐵脅迫的能力最強;而 T-05 最小,為 0,說明 T-05 表現為耐缺鐵性小。其中 T-05 和T-04 的多個耐缺鐵指標的隸屬函數值如活性鐵、SPAD 值、156 d 生物量、籽粒中鐵的含量和鐵的收獲指數等均最小,表現為耐缺鐵脅迫能力低,而 T-43在 28 d 和 156 d 生物量、53 d 時鐵的分配和產量的隸屬函數值均最大,說明在這些指標上 T-43 耐缺鐵能力較強。
各項指標變異系數在 5.1% 和 36.5% 之間波動,水稻 53 d 的生物量、葉片鐵含量和鐵的收獲指數變異系數較大,分別為 36.5%、25.8% 和 23.5%,說明這三個指標對缺鐵脅迫反應較為敏感。
根據全生育期各指標的隸屬函數值之和進行系統聚類分析 (表 5),比如生物量是以 28 d、56 d 和156 d 三個取樣時期的隸屬函數值之和進行聚類分析排名,結果發現 T-04 的生物量聚類排名最靠后,在生物量上表現為缺鐵敏感型水稻,T-201 和 T-43 的聚類分析排名靠前。以幼苗期葉片活性鐵含量、葉片 SPAD 值和分蘗期 (53 天) 葉片鐵含量隸屬函數值之和進行聚類分析排名發現,T-04 和 T-05 聚類分析排名得分最靠后。同樣 T-04 鐵的轉運能力也低于其他基因型水稻。T-69 鐵的收獲指數的聚類排名得分明顯高于其他基因型水稻,T-43、T-69 和 T-05 的產量和籽粒收獲指數也明顯高于其他基因型水稻。
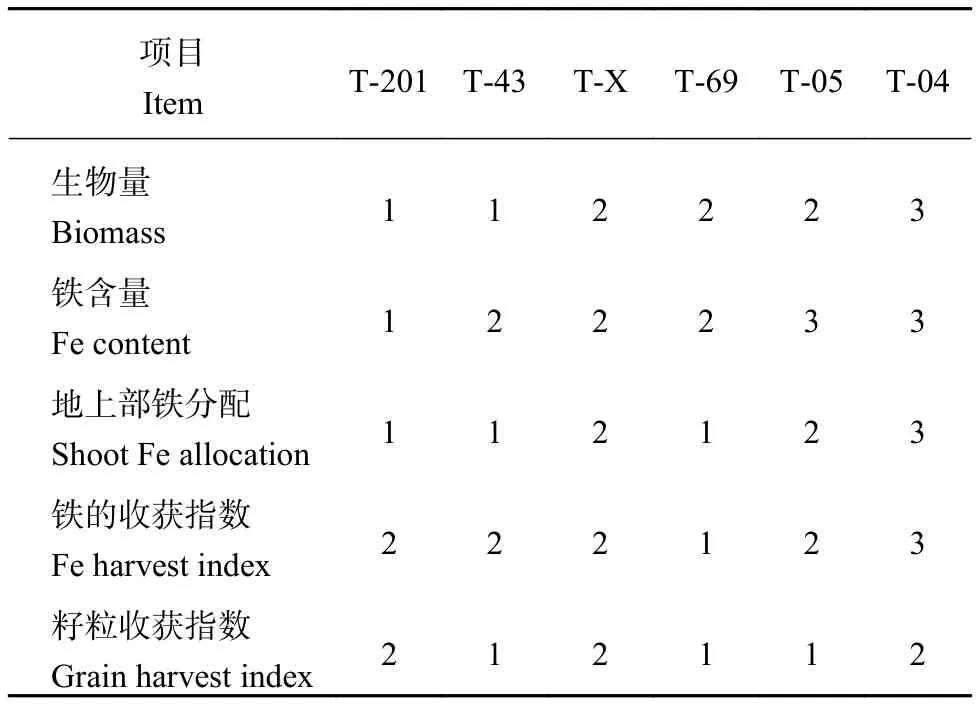
表5 不同基因型水稻全生育期各指標聚類分析排名Table5 Ranking of selected rice genotypes for the tested items
3 討論與結論
試驗結果表明,在相同的缺鐵土壤環境和田間管理下,膜下滴灌水稻在不同生育期對缺鐵的反應存在基因型差異,表現在葉片活性鐵含量、鐵的轉運、分蘗數和產量等指標上,這與不同種、屬的植物以及同一種不同品種對缺鐵的敏感程度和適應能力存在很大差異[25–27]一致。高麗等[28]研究結果也顯示在低鐵脅迫下花生葉綠素在品種間的變異系數顯著高于供鐵處理。基因型變異程度越大,說明該性狀指標越有利于基因型差異的顯示[29]。本研究中水稻分蘗期葉片鐵含量、53 d 生物量、籽粒鐵收獲指數和產量的變異系數都超過 20%,說明本實驗水稻的性狀指標反映了水稻耐缺鐵能力的基因型差異,可用于此缺鐵脅迫下水稻耐缺鐵能力的評價。
選擇適宜的鑒定指標和適宜的生育期是評價植物耐缺鐵性的重要基礎,如花生開花期葉片活性鐵含量和葉綠素含量與產量高度相關[30];結莢期葉綠素含量與花生單株果重呈二次拋物線關系[31],韓振海等[32]研究指出離子吸收動力學參數可作為鐵高效蘋果基因型植物的篩選指標;鄒春琴等[33]的研究表明活性鐵含量是診斷植物鐵營養狀況的可行指標。本試驗結果表明耐缺鐵水稻 T-43 幼苗期葉片活性鐵含量、分蘗期地上部鐵的分配和收獲期有效穗數均較高,同時在聚類分析中耐缺鐵水稻 T-43 全生育期地上部鐵的分配,生物量和籽粒收獲指數的排名均為 1,說明水稻根系中的鐵向地上部的及時供應,保證了水稻的正常生長和成熟期籽粒形成。
充足的有效穗數、穗粒數和穗型是水稻高產的特征[34]。朱青等[35]研究發現適量的亞鐵可提高水稻分蘗期的株高和分蘗數,李麗等[36]研究發現膜下滴灌水稻在分蘗期根系活力存在基因型差異,而根系活力是評價作物耐缺鐵能力的指標之一[17]。本研究發現,缺鐵脅迫下水稻的分蘗數存在顯著的基因型差異,缺鐵敏感型水稻 T-04 的分蘗數高于 T-43,但是其有效分蘗數與 T-43 無顯著差異,同時缺鐵敏感型水稻T-04 幼苗期地上部鐵含量和分蘗期葉片鐵含量也較低,這說明水稻自身鐵的有效利用可能影響著水稻分蘗的形成,而水稻的分蘗數通常與產量呈正相關,在水稻的分蘗期評價水稻的鐵營養狀況是必不可少的。在幼苗期和分蘗期,T-201 的多個指標,如葉片活性鐵含量、地上部鐵含量,地上部鐵的分配均較高,但是其產量卻較低,T-201 幼苗期和分蘗期的鐵營養效率與其豐產性沒有表現一致,說明 T-201僅可以作為耐缺鐵遺傳材料,而適宜種植的水稻基因型應兼有耐缺鐵、豐產等性狀。夏友霖等[31]的研究也出現類似的花生品種,由此體現在缺鐵脅迫土壤上進行作物全生育期培養,結合農藝性狀和鐵營養效率評價水稻耐缺鐵能力的必要性。膜下滴灌種植模式下水稻籽粒中鐵含量存在顯著的基因型差異,Gregorio 等[37]通過對不同來源的 1138 份水稻種質研究發現,稻米鐵含量為 6.3~24.4 mg/kg,其中 IR68144水稻子粒的鐵濃度較高,其遺傳性狀穩定,不受土壤水分、酸堿度和氮肥使用量的影響,Fan 等研究發現旱作水稻籽粒鐵的收獲指數受缺鐵脅迫的影響[12],水稻籽粒鐵生物有效性主要受基因型控制[38-39],籽粒鐵含量與人們的鐵營養密切相關,籽粒鐵含量也應該作為培育耐缺鐵水稻基因型的衡量指標之一,本試驗發現耐缺鐵性水稻籽粒鐵收獲指數高于缺鐵敏感型水稻。
本實驗結果表明,膜下滴灌節水栽培模式下,在不同的生育期結合水稻的農藝性狀和鐵吸收、轉移和利用評價水稻耐缺鐵能力是可行的。不同鐵敏感型水稻在不同生育期對缺鐵的反應存在顯著差異,初步確定 T-04 屬于對缺鐵敏感的水稻基因型,T-43 是耐缺鐵的基因型,其他品種的水稻屬于中等耐性的基因型。在幼苗期和分蘗期缺鐵脅迫下鐵敏感型水稻的生長、鐵的吸收和轉移能力弱,在生理成熟期其無效分蘗增加,培育耐缺鐵性水稻是提高水稻產量的經濟措施,但是膜下滴灌水稻缺鐵脅迫機理還需要深入研究。
[1]Bouman BA M, Humphreys E, Tuong TP and Barker R. Rice and water[J]. Advances in Agronomy, 2007, 92(4): 187–237.
[2]張正斌, 徐萍. 中國水資源和糧食安全問題探討[J]. 中國生態農業學報, 2008, 16(5): 1305–1310. Zhang ZB, Xu P. Water and food security in China[J]. Chinese Journal of Eco-Agriculture, 2008, 16(5): 1305–1310.
[3]薛全義, 荊宇, 華玉凡. 略論我國旱稻的生產及發展[J]. 中國稻米, 2002, 4(4): 5–7. Xue QY, Jing Y, Hua YF. Reflection on studies of upland rice production and development in China[J]. China Rice, 2002, 4(4): 5–7.
[4]Lu XH, Wu LH, Pang LJ, et al. Effects of plastic film mulching cultivation under non-flooded condition on rice quality[J]. Journal of the Science of Food & Agriculture, 2007, 87(2): 334–339.
[5]郭慶人, 陳林. 水稻膜下滴灌栽培技術在我國發展的優勢及前景分析[J]. 中國稻米, 2012, 18(4): 36–39. Guo QR, Chen L. The advantages and prospects of rice cultivation techniques under-mulch-drip irrigation[J]. China Rice, 2012, 18(4): 36–39.
[6]Zhang XJ, Zhang SJ, Zhao HH, et al. Effect of HCO3–on rice growth and iron uptake under flood irrigation and drip irrigation with plastic film mulch[J]. Journal of Plant Nutrition and Soil Science, 2015, 178(6): 944–952.
[7]張書捷, 張新疆, 王娟, 等. 施用土壤酸化劑和調整播期防治膜下滴灌水稻苗期缺鐵黃化的效果研究[J]. 中國水稻科學, 2015, 29(5): 519–527. Zhang SJ, Zhang XJ, Wang J, et al. Applying soil acidification agents and adjusting sowing date to deal with rice chlorosis in seedling stage caused by ion deficiency under drip irrigation with plastic film mulching[J]. China Rice Science, 2015, 29(5): 519–527.
[8]Ishimaru Y, Kim S, Tsukamoto T, et al. Mutational reconstructed ferric chelate reductase confers enhanced tolerance in rice to iron deficiency in calcareous soil[J]. Proceedings of the National Academy of Sciences of the United States of America, 2007, 104(18): 7373–7378.
[9]Cao X, Chen Y, Wang X, Deng X. Effects of redox potential and pH value on the release of rare earth elements from soil[J]. Chemosphere, 2001, 44(4): 655–661.
[10]Hasegawa H, Rahman MM, Kadohashi K, et al. Significance of the concentration of chelating ligands on Fe3+-solubility, bioavailability, and uptake in rice plant[J]. Plant Physiology and Biochemistry, 2012, 58: 205–211.
[11]Matsuo N, Mochizuki T. Growth and yield of six rice cultivars under three water-saving cultivations[J]. Plant Production Science, 2009, 12(4): 514–525.
[12]Fan X, Karim MR, Chen X, et al. Growth and iron uptake of lowland and aerobic rice genotypes under flooded and aerobic cultivation[J]. Communications in Soil Science and Plant Analysis, 2012, 43(13): 1811–1822.
[13]Brown JC, Ambler JE, Chaney RL, Foy CD. Differential responses of plant genotypes to micronutrients [A]. Mortvedt J, Giordano PM, Lindsay WL. Micronutrients in agriculture [M]. Madison, Wisconsin: Soil Science Society of America. 389–418.
[14]Gao L, Shi Y. Genetic differences in resistance to iron deficiency chlorosis in peanut[J]. Journal of Plant Nutrition, 2007, 30(1): 37–52.
[15]Van Egmond F, Akta? M. Iron-nutritional aspects of the ionic balance of plants[J]. Plant and Soil, 1977, 48(3): 685–703.
[16]張福鎖, 劉書娟, 毛達如, 韓振海. 蘋果抗缺鐵基因型差異的生理生化指標研究[J]. 園藝學報, 1995, (1): 1–6. Zhang FS, Liu SJ, Mao DR, Han ZH. Genotypic differences in iron-deficiency stress respones in Malu[J]. Acta Horticulturae Sinica, 1995, (1): 1–6.
[17]王永章, 張玉娜, 徐坤. 供鐵水平對不同蘋果基因型生長及根系特性的影響[J]. 萊陽農學院學報, 2000, 17(3): 161–165. Wang YZ, Zhang YN, Xu K. Effects of iron levels on seedling growth and root characteristics of iron-efficient or-inefficient species in Malus[J]. Journal of Laiyang Agricultural College, 2000, 17(3): 161–165.
[18]Itai RN, Ogo Y, Kobayashi T, et al. Rice genes involved in phytosiderophore biosynthesis are synchronously regulated during the early stages of iron deficiency in roots[J]. Rice, 2013, 6(1): 16.
[19]Bashir K, Hanada K, Shimizu M, et al. Transcriptomic analysis of rice in response to iron deficiency and excess[J]. Rice, 2014, 7(1): 18.
[20]Kobayashi T, Itai RN, Nishizawa NK. Iron deficiency responses in rice roots[J]. Rice, 2014, 7(1): 27.
[21]Mori S, Nishizawa N, Hayashi H, et al. Why are young rice plants highly susceptible to iron deficiency?[J]. Plant and Soil, 1991, 130(1): 143–156.
[22]Dos Santos LS, De Oliveira AC. Rice iron metabolism: from source to solution[J]. Journal of Crop Science and Biotechnology, 2007, 10(2): 64–72.
[23]Bashir K, Ishimaru Y, Nishizawa NK. Iron uptake and loading into rice grains[J]. Rice, 2010, 3(2–3): 122–130.
[24]Zeng L, Shannon MC, Grieve CM. Evaluation of salt tolerance in rice genotypes by multiple agronomic parameters[J]. Euphytica, 2002, 127(2): 235–245.
[25]Kannan S. Cultivar differences for tolerance to Fe and Zn deficiency: A comparison of two maize hybrids and their parents[J]. Journal of Plant Nutrition, 1983, 6(4): 323–337.
[26]石榮麗, 張福鎖, 鄒春琴. 不同基因型小麥鐵營養效率差異及其可能機制[J]. 植物營養與肥料學報, 2010, 16(6): 1306–1311. Shi RL, Zhang FS, Zou CQ. Iron efficiency of different wheat genotypes and its main contributed factors[J]. Plant Nutrition and Fertilizer science, 2010, 16(6): 1306–1311.
[27]郝紅梅, 鄒春琴, 張福鎖. 不同旱稻品種(系)間鐵營養狀況的比較[J]. 中國農業大學學報, 2003, 8(5): 79–82. Hao HM, Zou CQ, Zhang FS. Difference in iron nutritional status of different aerobic rice varieties (lines)[J]. Journal of China Agricultural University, 2003, 8(5): 79–82.
[28]高麗, 周健民, 史衍璽. 石灰性土壤上花生耐低鐵品種的篩選[J].中國油料作物學報, 2008, 30(2): 168–173. Gao L, Zhou JM, Shi YX. Tolerance screening of peanut varieties to iron deficiency chlorosis on calcareous soil[J]. Chinese Journal of Oil Crop Sciences, 2008, 30(2): 168–173.
[29]韓璐, 張薇. 棉花苗期氮營養高效品種篩選[J]. 中國農學通報, 2011, 27(1): 84–88. Han L, Zhang W. Screening of cotton varieties with high nitrogen efficiency at seedling stage[J]. Chinese Agricultural Science Bulletin, 2011, 27(1): 84–88.
[30]高麗, 史衍璽, 周健民. 花生缺鐵黃化的敏感時期及耐低鐵品種的篩選指標[J]. 植物營養與肥學報, 2009, 15(4): 917–922. Gao L, Shi YX, Zhou JM. Study on the sensitive period and screening index for iron deficiency chlorosis in peanut[J]. Plant Nutrition and Fertilizer Science, 2009, 15(4): 917–922.
[31]夏友霖, 廖伯壽, 毛金雄, 等. 四川丘陵紫色土花生品種耐缺鐵性鑒定與評價[J]. 中國油料作物學報, 2013, 35(3): 326–330. Xia YL, Liao BS, Mao JX, et al. Evaluation of lime induced irondeficiency chlorosis tolerance of peanut on calcareous and purplish soil in Sichuan[J]. Chinese Journal of Oil Crop Sciences, 2013, 35(3): 326–330.
[32]韓振海, 王永章, 孫文彬. 鐵高效及低效蘋果基因型的鐵離子吸收動力學研究[J]. 園藝學報, 1995, 22(4): 313–317. Han ZH, Wang YZ, Sun WB. Iron absorption kinetics for Feefficient vs-ineiffeient species in Malus[J]. Acta Horticulturae Sinica, 1995, 22(4): 313–317.
[33]鄒春琴, 陳新平, 張福鎖, 毛達如. 活性鐵作為植物鐵營養狀況診斷指標的相關研究[J]. 植物營養與肥料學報, 1998, 4(4): 399–406. Zou CQ, Chen XP, Zhang FS, Mao DR. Study on the correlation between the active Fe and Fe nutritional status of plants[J]. Plant Nutrition and Fertilizer Science, 1998, 4(4): 399–406.
[34]趙雙玲, 李向春, 陳林, 等. 膜下滴灌水稻品種產量、品質和農藝性狀的相關性分析[J]. 北方水稻, 2015, 3(3): 6–9, 13. Zhao SL, Li XC, Chen L, et al. Correlation analysis among yield, quality and agronomic trait in rice with drip irrigation under mulch film[J]. Journal of North Rice, 2015, 3(3): 6–9, 13.
[35]朱青, 盧松, 陳正剛, 等. 亞鐵對水稻分蘗期生長發育的影響[J]. 耕作與栽培, 2012, (6): 1–2. Zhu Q, Lu S, Chen ZG, et al. The effect of ferrous on rice growth at tillering stage[J]. Tillage and Cultivation, 2012, (6): 1–2.
[36]李麗, 陳林, 張婷婷, 等. 膜下滴灌對水稻根系形態及生理性狀的影響[J]. 排灌機械工程學報, 2015, 33(6): 536–540. Li L, Chen L, Zhang TT, et al. Effects of mulch drip irrigation on the form of rice root and physiological characteristics[J]. Journal of Drainage and Irrigation Machinery Engineering, 2015, 33(6): 536–540.
[37]Gregorio GB, Senadhira D, Htut T, et al. Improving iron and zinc value of rice for human nutrition[J]. Agriculture et Development, 1999, 23(9): 77–81.
[38]孫明茂, 洪夏鐵, 李圭星, 等. 水稻籽粒微量元素含量的遺傳研究進展[J]. 中國農業科學, 2006, 39(10): 1947–1955. Sun MM, Hong XT, Li GX, et al. Progress of genetic research on trace minerals content in rice seed[J]. Scientia Agricultura Sinica, 2006, 39(10): 1947–1955.
[39]何萬領. 水稻品種籽粒中鐵生物有效性差異及機制[D]. 杭州: 浙江大學博士學位論文, 2009. He WL. Difference and mechanism of iron bioavailability in grain of different rice genotypes[D]. Hangzhou: PhD Dissertation of Zhejiang University, 2009.
Evaluating of rice genotypes tolerant to Fe deficiency with drip irrigation under the film mulch
LI Yan-yan, BAI Ru-xiao, ZHANG Xin-jiang, YANG Yu-zhen, HUANG Zhi-hua, HOU Jian-wei, WEI Chang-zhou*
( College of Agronomy, Shihezi University, Shihezi, Xinjiang 832003, China )
【Objectives】Rice is grown on calcareous soil and managed using drip irrigation under the film mulch technology in the Xinjiang Uyghur Autonomous Region, where rice often suffers chlorosis caused by Fe deficiency. Evaluating the Fe-stress tolerance of different rice genotypes is of great importance for screening suitable rice cultivar. 【Methods】Six rice cultivars were selected and managed with drip irrigation under the film mulch method in this research. At seedling, tillering and mature stages of rice, the biomass, tillers, yield and Fe content were analyzed. Cluster analysis and subordinate function values analysis were used to identify the multiple parameters associated with iron deficiency tolerance. 【Results】The 6 tested rice cultivars showed significant differences in resistance to iron deficiency at the seedling, tillering and mature stages. The leaf active iron contents of T-04 and T-05 were significantly lower than those of the other genotypes at seedling stage. T-201 had higher leaf active iron and Fe distribution in shoot at the seedling stage and leaf iron content and dry weight in shoot at tillering stage, T-04 had the lowest distribution of Fe in shoot at the tillering stage, while T-04 had the biggest tiller number. There were no significant differences in effective tiller number between T-04 and other genotypes, and yield of T-04 was the medium level. Cluster analysis showed that T-43 had higher indexes than other genotypes. 【Conclusions】The effective use of leaf iron and the ability of iron transfer ensure rice growth and higher effective panicle number, comprehensively considering iron nutrition efficiency and yield at all growthstages with drip irrigation under the film mulch, the rice cultivar T-43 has the strongest tolerance to iron deficiency, and T-04 shows the lowest tolerance at all growth stages.
rice; drip irrigation under film mulch; genotype; iron deficiency tolerance
2016–08–02 接受日期:2016–11–07
國家 863 計劃資助項目(2011AA100508);國家自然科學基金資助項目(31471947)資助。
李言言(1989—),女,河南商丘人,碩士研究生,主要從事膜下滴灌水稻苗期缺鐵黃化機理研究。E-mail:lyy9085@126.com * 通信作者 E-mail:czwei@shzu.edu.cn
猜你喜歡
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
音樂探索(2022年2期)2022-05-30 21:01:37
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
