蛙攜帶虹彩病毒的分子流行病學研究
2017-06-10 19:03:25邵軍輝張利峰王樹云李聰羅
湖北農業科學 2017年9期
關鍵詞:流行病學
邵軍輝++張利峰++王樹云++李聰++羅楊志++陳孝煊++袁軍法

摘要:運用多重實時PCR對多地的養殖和野生蛙類攜帶虹彩病毒的本底展開調查,并進行病毒分型。結果表明,檢測的樣品平均陽性率為8.90%,養殖蛙的平均陽性率為4.57%,野生蛙的平均陽性率為16.82%。從蛙種類來看,養殖牛蛙和黑斑蛙帶毒率較低,而養殖的棘胸蛙和野外捕捉的斑腿樹蛙帶毒率較高。不同蛙攜帶的蛙病毒的分子序列比較結果顯示,不同來源的蛙病毒高度保守,缺乏明顯的宿主與地域特異性,表明人工繁育場暴發虹彩病毒可能對其他野生蛙類資源產生了負面影響,建議在野生動物人工馴養繁殖體系中引入病害監測和疫病防控措施。
關鍵詞:蛙;虹彩病毒;多重實時PCR;流行病學
中圖分類號:Q959.5+3 文獻標識碼:A 文章編號:0439-8114(2017)09-1702-04
DOI:10.14088/j.cnki.issn0439-8114.2017.09.025
Study on Molecular Epidemiology of Iridoviruses in Rana
SHAO Jun-hui1,ZHANG Li-feng2,WANG Shu-yun2,LI Cong1,LUO Yang-zhi1,CHEN Xiao-xuan1,YUAN Jun-fa1
(1. College of Fisheries, Huazhong Agricultural University, Wuhan 430070, China;2. Inspection and Quarantine Technical Center of Beijing Entry-Exit Inspection and Quarantine Bureau, Beijing 100026, China)
Abstract: The multiplex real-time PCR assay was used to investigate the prevalence of iridoviruses in farmed or wild frog. Results indicated that the average positive rate was 8.90%,in which the average positive rate in farmed and wild frog was 4.57% and 16.82%,respectively. The most significant prevalence of iridoviruses was found among the farmed Quasipaa spinosa and Polypedates megacephalus,while the species of Rana catesbeiana and Rana nigromaculata rarely. Sequences alignement of major capsid protein suggested that there were no obvious host and geographic specificity among these detected Rana iridoviruses,it indicated that the outburst of iridoviruses in farm produced negative effects on wild frogs,disease monitoring,disease prevention and control measures were proposed to introduce into the artificial domestication propagation system of wild animal.
Key words: Rana; iridoviruses; multiplex real-time PCR; epidemiology
近50年來,全球范圍內多種兩棲動物種群顯著衰退,一些種類已經滅絕,部分物種瀕危,原因包括氣候變化、棲息地減少、環境污染以及疾病傳播等[1,2]。其中蛙壺菌(Batrachochytrium dendrobatidis)和虹彩病毒(Iridoviruses)是引起兩棲類種群數量下降的主要病原因素[3-5]。中國部分蛙種的資源量也因環境變化以及人為捕捉等因素急劇減少。如虎紋蛙(Hoplobatrachus ruguosus)、林蛙(Rana dybowskii)、棘胸蛙(Quasipaa spinosa)等種類因肉味鮮美、營養豐富遭到過度捕捉,導致這些蛙類野生資源量急劇減少[6,7]。近年來,為保護自然資源及滿足市場需求,人們開展了多個蛙種的馴養及人工增養殖[8-10]。但人工增養殖的模式對野生蛙類資源以及當地自然生態環境的影響缺乏系統的評估,尤其是人工增養殖中虹彩病毒病的暴發對野生資源的影響缺乏深入研究。
虹彩病毒無囊膜,對環境適應力較強,可在土壤和水體中存活長達幾個月而保持感染性[11]。為適應蛙的特殊生活習性,養殖場多位于較偏僻的山林地區,與周邊環境存在許多交互作用。當蛙養殖場暴發虹彩病毒病后,養殖用水的不規范處理、蛙的逃逸以及帶毒蛙的商業流動等對當地野生蛙類資源的影響亟需評估。先前,湖北省宜昌市某養殖場發生棘胸蛙的暴發性死亡,經鑒定為蛙病毒的感染所致[12],但其病毒來源以及蛙攜帶病毒的本底等基礎問題尚未知。為此,本研究利用建立的蛙病毒多重檢測方法[13],對不同來源的養殖和野生蛙類攜帶虹彩病毒進行初步調查,并進行病毒分型研究,以期為優化人工增養殖的蛙類資源保護模式提供參考。
1 材料與方法
1.1 樣品采集
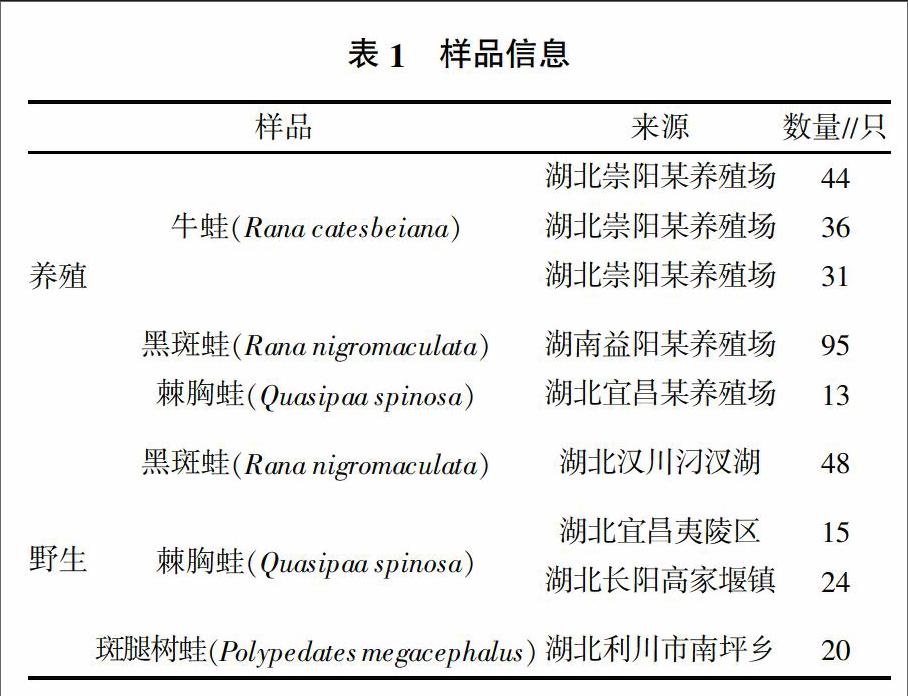
樣品詳細采樣情況見表1。采集的蛙經麻醉后取脾或腎用于后續的檢測。
1.2 樣品DNA提取與篩查
組織DNA的提取采用康為世紀通用型柱式基因組提取試劑盒,取不超過0.1 g組織,按試劑盒說明書進行,組織DNA以50 μL的三蒸餾水洗脫,-20 ℃保存。
以優化的三重熒光PCR方法篩查[13]。優化的三重熒光PCR反應體系(25 μL)如下:5 μL DNA模板,10×recreation buffer 2.5 μL,25 mmol/L MgCl2 3 μL,10 mmol/L dNTP 0.5 μL,通用正向引物(20 μmol/L)1 μL,通用反向引物(20 μmol/L)1 μL,探針1(20 μmol/L)0.5 μL,探針2(20 μmol/L)0.5 μL,探針3(20 μmol/L)0.5 μL,Taq DNA聚合酶(5 U/μL)0.5 μL,用去離子水補足至總體積25 μL。采用Roche Light Cycler 480Ⅱ熒光PCR檢測儀:第一階段:95 ℃ 3 min;第二階段:95 ℃ 15 s,54 ℃ 15 s, 60 ℃ 30 s,40個循環;60 ℃延伸時收集熒光。擴增結果以在相應的檢測通道呈典型的“S”型擴增曲線及Ct值判定。
1.3 基因擴增與克隆
以蛙病毒保守引物RGVP1/P2(5-GACTTGGC
CACTTATGAC-3/5-GTCTCTGGAGAAGAAGAA-3)擴增“1.2”中準備的部分樣品DNA,擴增循環條件為95 ℃ 30 s,50 ℃ 45 s,72 ℃ 30 s,35個循環,預期擴增片段為531 bp,預期目的條帶經膠回收后直接測序驗證。
以NCBI登陸的蛙病毒主要結構蛋白(major capsid protein,MCP)序列設計引物MCPF1/R1(5-ATGTCTTCTGTAACCGGTTCAG-3/5-TTACAAGA
TTGGGAATCCCATCG-3),用于擴增蛙病毒的MCP全長,擴增循環條件為95 ℃ 30 s,50 ℃ 45 s,72 ℃ 90 s,30個循環,預期擴增片段為1 392 bp。擴增產物經膠回收后連接至T-easy載體,每個基因選擇3個陽性克隆子測序。
1.4 序列分析
以DNAstar進行常規的序列拼接與編輯,序列比對以Clustal 2.0軟件進行,聚類分析以MEGA 6.0進行,以鄰接法構建進化樹,選擇成對缺失模式,以bootstrip值進行驗證,進化樹以Treeview進行編輯。
2 結果與分析
2.1 樣品篩查結果
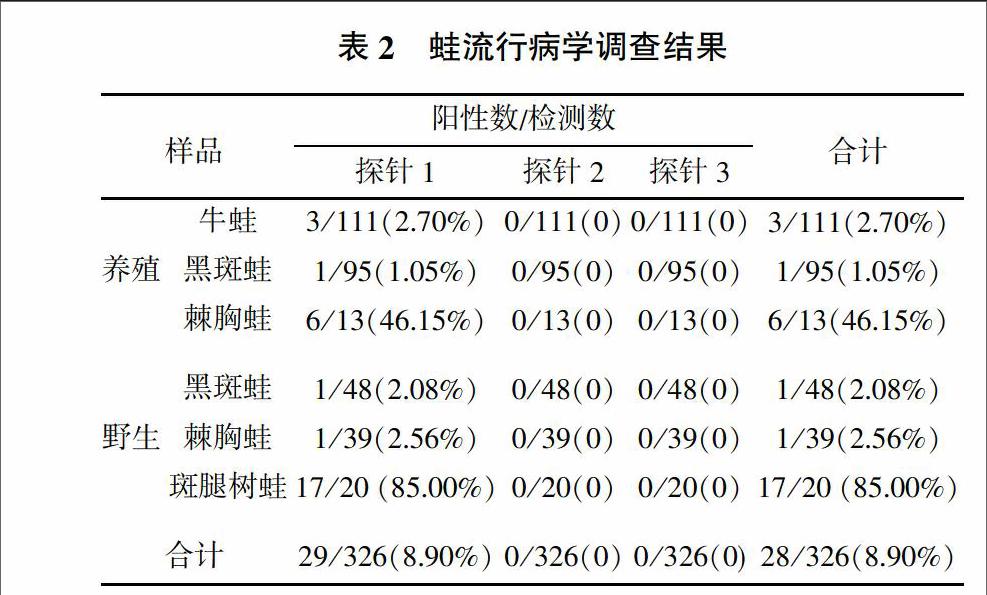
先前建立的三重熒光定量PCR可檢測不同種類的蛙病毒,其中探針1可檢測包含蛙病毒3型(Frog virus 3,FV3)、飾紋汀蛙虹彩病毒(Bohel iridovirus,BIV)、流行性造血器官壞死癥病毒(Epizootic Haematopoietic Necrosis Virus,EHNV)、歐洲鮰魚病毒(European catfish iridovirus,ECV)、歐鯰病毒(European sheetfish iridovirus,ESV)、中華鱉虹彩病毒(Soft-shelled Turtle iridovirus,STIV)、大鯢虹彩病毒(Andrias davidiamus iridovirus,ADIV)、沼澤綠牛蛙虹彩病毒(Rana grylio virus,RGV)、虎紋蛙病毒(Tiger frog virus,TFV)等的蛙病毒,探針2可檢測大口黑鱸虹彩病毒(Largemouth bass ranavirus,LMBV)、裂唇魚病毒(Doctor fish virus,DFV)和孔雀魚病毒(Guppyfish iridovirus,GV6),探針3可檢測新加坡石斑魚虹彩病毒(Singapore grouper iridovirus,SGIV)[13]。樣品篩查結果(表2)顯示,探針1檢測的陽性率為8.90%,從養殖的219份蛙樣品中檢出10份陽性,平均Ct值為26.45,最小Ct值為20.03。從野外捕捉的107份蛙樣品中檢出19份陽性,平均Ct值為28.18,最小Ct值為22.97。從蛙種類來看,牛蛙和黑斑蛙帶毒率較低,而野外捕捉的斑腿樹蛙帶毒率較高。探針2和3均未檢出陽性。
2.2 不同來源蛙攜帶病毒的比較
因探針1可檢測多種蛙病毒,為確定檢測到的蛙病毒屬于哪一種,進一步以蛙病毒MCP保守引物進行擴增,擴增片段為531 bp,并進行測序。Blast比對結果(圖1)顯示,獲得的MCP部分序列高度保守,同Rana grylio virus 9807高度保守,僅養殖來源的棘胸蛙(QSV46)同其他序列相差一個堿基,并導致一個氨基酸的差異。
為比較上述不同來源的蛙病毒的差異,設計引物擴增養殖的棘胸蛙來源(QSV46)與斑腿樹蛙(PMV69)攜帶病毒的MCP全長。如圖1所示,QSV46和PMV69的核苷酸序列僅相差4個堿基,并導致4個氨基酸的差異。同蛙病毒屬的代表種FV3編碼的MCP比較,QSV46同FV3相差5個氨基酸,PMV69同FV3僅相差2個氨基酸。值得指出的是,PMV69的序列同本實驗室先前從某養殖場獲得的QSV01 MCP全長序列完全一致[12],但兩者種類不同、來源不一致、采樣地點的距離超過400 km。同中國其他來源的蛙病毒比較顯示,不同蛙類攜帶的FV3或FV3樣病毒,如RGV、TFV同源性都非常高。
從GenBank獲得虹彩病毒科各代表種的MCP全長序列,與本研究中獲得的MCP全長進行比對,并構建進化樹(圖2),由圖2可見,QSV01、QSV46以及PMV69同FV3和STIV聚為一支,均為蛙病毒屬的成員。
以ClustalX2比對虹彩病毒科各代表種編碼的MCP氨基酸序列,以MEGA6構建進化樹。參考序列如下:STIV(EU627010)、RGV(JQ654586)、FV3(AY548484)、TFV(AF389451)、ATV(AY150217)、GSIV(AET51835.1)、ISKNV(AF371960)、RBIV(AY532606)、OSGIV(AY894343)、LCDV-C(AY380826)、LCDV-1(L63545)、IIV6(NP149737.1)、IIV3(DQ643392);QSV01、QSV46和PMV69為本研究涉及;GSIV-HN為華中農業大學水生動物病毒實驗室于2012年從河南發病大鯢分離獲得。
3 小結與討論
近年來,氣候變化與人類活動是導致蛙類種群數量下降的主要原因,但蛙壺菌和蛙病毒等病原生物的感染也是導致蛙種群數量降低的重要原因[1-5]。據報道,目前蛙病毒感染的兩棲類種類多達74種,其中主要為無尾目(Anura)蛙科(Ranidae)的成員[4]。蛙病毒可能引起蛙類的致死性感染,如Green等[14]分析了1996-2001年間美國兩棲類死亡事件,其中57%的兩棲類死亡事件與蛙病毒有關。張奇亞等[15]、王曉紅等[16]、劉曉東等[17]分別從沼澤綠牛蛙(Rana grylio)、虎紋蛙(Rana tigrina rugulosa)、牛蛙(Rana catesbeiana)等養殖蛙上分離到蛙病毒,李莉娟等[12]首次從人工養殖的患病棘胸蛙中分離到蛙病毒QSV01。蛙病毒除了致死性感染外,也能形成無臨床癥狀的攜帶狀態。如Xu等[18]對黑龍江省的東北林蛙攜帶蛙病毒的初步調查顯示東北林蛙成體的帶毒率為5%(9/180),夏季蝌蚪的平均帶毒率高達42.5%(51/120)。本研究也顯示從養殖或野外捕捉的蛙攜帶病毒的平均比例為8.90%(29/326)。野生蛙類的帶毒狀態對蛙類種群的潛在影響尚缺乏系統評估。存在因棲息地減少、食物缺乏以及環境劇變等強應激下,蛙病毒從攜帶感染(或潛伏感染)轉化為急性感染、引起蛙的死亡的潛在威脅。亟需利用生態學與流行病學等多學科的研究手段,在生態學的大尺度上評估蛙攜帶病毒對其種群的影響。
綜合已有的研究,野生蛙攜帶病毒的比率受調查種類、棲息環境以及采樣季節的影響,波動幅度非常大(0%~80%)[4]。在本研究中,不同來源與種類的野生蛙帶毒比率差別也非常大,其中斑腿樹蛙的帶毒率最高,黑斑蛙帶毒率最低,僅2%,可能與不同蛙種對蛙病毒的敏感性有差異相關。Hoverman等[19]的研究已證實不同蛙類對蛙病毒的敏感性存在差異,他們在調查北美的19個蛙種中,木蛙(Lithobates sylvaticus)和穴蛙(L. capito)的敏感性最高。
通過比較不同蛙攜帶的蛙病毒的分子序列,顯示不同來源的蛙病毒高度保守,缺乏明顯的宿主與地域特異性。如從患病沼澤綠牛蛙中分離到蛙病毒(RGV9506、RGV9807、RGV9808)、從牛蛙中分離的蛙病毒(FJ049)、從虎紋蛙中分離的蛙病毒(TFV)以及從棘胸蛙分離的蛙病毒株(QSV01、MV69)都高度相似,氨基酸一致率在99%以上,其編碼的MCP僅相差0~5個氨基酸(圖1)。值得指出的是,QSV01來源于患病棘胸蛙,PMV69來源于野外捕捉的斑腿樹蛙,兩個樣本來自不同地域,但兩者編碼的MCP完全一致,提示蛙病毒沒有嚴格的宿主特異性,很可能存在跨種間傳播的現象,為蛙類野生資源保護提出了更為嚴峻的現實難題。根據《中華人民共和國野生動物保護法》等規定,國家鼓勵馴養繁殖野生動物。但人工馴養與增養殖的野生動物保護策略,尤其是蛙類、珍稀魚類等群體生物,隨著養殖密度的提高、養殖環境的應激增強等因素極大地增加了疾病傳播的幾率。同時,這些養殖場多同自然環境交互較密切,養殖廢水的消毒不徹底、養殖對象的逃逸、帶毒動物的商業化流通甚至放流等因素很可能對野生種質資源造成負面影響。因此在野生動物人工馴養繁殖體系中建立病害監測、引入系統的疫病防控策略顯得尤為重要和迫切。
參考文獻:
[1] STUART S N,CHANSON J S,COX N A,et al. Status and trends of amphibian declines and extinctions worldwide[J].Science,2004,306:1783-1786.
[2] RACHOWICZ L J,KNAPP R A,MORGAN J A,et al. Emerging infectious disease as a proximate cause of amphibian mass mortality[J].Ecology,2006,87(7):1671-1683.
[3] CHINCHAR V G,WALTZEK T B. Ranaviruses: not just for frogs[J].PLoS Pathog,2014,10(1):e1003850.
[4] MILLER D,GRAY M,STORFER A. Ecopathology of ranaviruses infecting amphibians[J].Viruses,2011,3(11):2351-2373.
[5] STOHR A C,LOPEZ-BUENO A,BLAHAK S,et al. Phylogeny and differentiation of reptilian and amphibian ranaviruses detected in Europe[J]. PLoS One,2015,10(2):e0118633.
[6] ESKEW E A,TODD B D. Parallels in amphibian and bat declines from pathogenic fungi[J].Emerging Infectious Diseases,2013,19(3):379-385.
[7] 劉 欣,張 偉,遇寶成,等.東北三省中國林蛙資源調查研究[J].林業資源管理,2007(3):82-85.
[8] 梅祎蕓,葉容暉,宋婷婷,等.浙江省棘胸蛙養殖現狀及發展對策[J].浙江農業科學,2015,56(7):1122-1125.
[9] 俞寶根,葉容暉,鄭榮泉,等.人工環境下棘胸蛙(Paa spinosa)繁殖期的行為譜及活動節律[J].生態學報,2008,28(12):6371-6378.
[10] 王 兵. 遼寧省中國林蛙馴養繁殖現狀、問題與對策建議[J].遼寧林業科技,2012(3):52-54.
[11] NAZIR J,SPENGLER M, MARSCHANG R E. Environmental persistence of amphibian and reptilian ranaviruses[J].Dis Aquat Organ,2012,98(3):177-184.
[12] 李莉娟,羅楊志,顧澤茂,等.蛙病毒3介導棘胸蛙的暴發性死亡[A].中國水產學會,中國科學院水生生物研究所,海南大學.2013年中國水產學會魚病專業委員會學術研討會論文集[C].海口,2016.
[13] 李江宇,王樹云,王 姝,等.蛙病毒三重熒光PCR檢測方法的建立[J].高技術通訊,2015,25(7):746-752.
[14] GREEN D E,CONVERSE K A,SCHRADER A K,et al. Epizootiology of sixty-four amphibian morbidity and mortality events in the USA,1996-2001[J].Ann NY Acad Sci,2002, 969(1):323-339.
[15] 張奇亞,李正秋,江育林,等.沼澤綠牛蛙病毒的分離及其細胞感染的初步研究[J].水生生物學報,1996,20(4):390-392.
[16] 王曉紅,翁少萍,何建國.虎紋蛙病毒體外培養及其理化特性[J].水產學報,2002,26(4):363-367.
[17] 劉曉東,楊金先,龔 暉,等.一株牛蛙源虹彩病毒的分離及鑒定[J].中國動物傳染病學報,2012,20(1):16-20.
[18] XU K,ZHU DZ,WEI Y,et al. Broad distribution of Ranavirus in free-ranging Rana dybowskii in Heilongjiang,China[J].Ecohealth,2010,7(1):18-23.
[19] HOVERMAN J T,GRAY M J,HAISLIP N A,et al. Phylogeny,life history,and ecology contribute to differences in amphibian susceptibility to ranaviruses[J].Ecohealth,2011,8(3):301-319.
猜你喜歡
現代畜牧科技(2021年8期)2021-10-13 07:22:14
現代畜牧科技(2021年9期)2021-10-13 06:39:12
現代畜牧科技(2021年3期)2021-07-21 08:42:04
現代畜牧科技(2021年3期)2021-07-21 08:42:02
現代畜牧科技(2021年6期)2021-07-16 05:50:28
現代畜牧科技(2021年6期)2021-07-16 05:50:22
現代畜牧科技(2021年6期)2021-07-16 05:50:10
現代畜牧科技(2021年2期)2021-03-19 07:49:16
現代畜牧科技(2021年2期)2021-03-19 07:48:58
世界科學技術-中醫藥現代化(2020年2期)2020-07-25 02:05:36
