大米蛋白的BN大鼠致敏動物模型建立
2017-06-01 12:21:12王一俠蔡木易谷瑞增
食品工業科技 2017年5期
王一俠,尹 曼,馬 濤,魯 軍,蔡木易,谷瑞增
(中國食品發酵工業研究院,北京市蛋白功能肽工程技術研究中心,北京 100015)
大米蛋白的BN大鼠致敏動物模型建立
王一俠,尹 曼,馬 濤,魯 軍,蔡木易,谷瑞增*
(中國食品發酵工業研究院,北京市蛋白功能肽工程技術研究中心,北京 100015)
以大米蛋白為過敏原,腹腔注射致敏雌性棕色挪威大鼠(BN大鼠),建立大米蛋白過敏BN大鼠動物模型。觀察BN大鼠全身主動過敏反應及體重、體溫、免疫臟器指數變化,檢測其血清特異性IgG、總IgE和特異性Ig E水平,并測定血漿組胺釋放水平及各組大鼠的血象變化。結果顯示,實驗第42 d大米蛋白激發實驗后,實驗組大鼠全部呈過敏反應陽性,且體溫降低,脾臟指數顯著增大(p<0.01),體內白細胞和嗜酸粒細胞數目升高(p<0.05);實驗組血清特異性IgG顯著升高(p<0.01),總IgE以及血漿組胺釋放量均高于對照組(p<0.05),實驗第14 d和42 d血清特異性IgE高于對照組,第28 d無明顯變化(p<0.05)。本實驗結果證明BN大鼠是評價大米蛋白致敏性較為理想的動物模型。
大米蛋白,食物過敏原,動物模型,抗體,組胺
近年來,隨著食物過敏發生率的不斷增加,食物過敏已成為一個食品安全和公共衛生問題。大米是是重要的糧食作物之一,全球半數以上的人口以大米為主食[1]。大米蛋白含有豐富的必需氨基酸,其氨基酸組成模式與WHO/FAO推薦的模式接近,易于被人體消化吸收,且生物效價及蛋白質利用率均較高。
最新研究表明,大米中含有一些過敏蛋白,包括14~16、26、33、56、60 ku等不同分子質量大小的蛋白,其中研究較普遍的是14~16 ku的α-淀粉酶/胰蛋白酶抑制劑和33 ku具有乙二醛酶Ⅰ活性的蛋白[2]。國內外的研究大多集中在過敏原提取純化方面,只有很少研究關注建立大米過敏動物模型,如XiaoWei Chen[3]等利用BALB/c小鼠灌胃大米蛋白致敏,以期建立大米蛋白過敏動物模型。但同時有研究證明BALB/c小鼠并不適合作為食物過敏的動物模型,且灌胃易產生口服耐受[4]。BN大鼠是具有高IgE反應性的種系動物,有過敏反應傾向,根據部分研究及國家標準顯示,雌性BN大鼠較雄性BN大鼠更易致敏[5-6]。因此,

表1 過敏反應級數
本研究以提取純化并經SDS-PAGE蛋白凝膠電泳和基質輔助激光解吸附飛行時間質譜(MALDI-TOF-MS)共同驗證的相對分子質量為14.4~18.4 ku的大米過敏原球蛋白為致敏原,對雌性棕色挪威大鼠(BN大鼠)進行腹腔注射致敏處理,觀察其全身主動過敏反應,并檢測特異性抗體、組胺、血象變化等相應過敏指標,初步建立大米蛋白致敏動物模型,為下一步建立大米蛋白致敏細胞模型和深入了解大米過敏引發的免疫反應機制提供基礎。
1 材料與方法
1.1 材料與儀器
雌性棕色挪威大鼠(BN大鼠),4~6周齡,12只,體重(40~60 g),由維通利華動物公司提供,許可證號為SCXK(京)2012-0001;大米 超市。
大鼠IgE酶聯免疫試劑盒和大鼠組胺試劑盒 上海酶聯免疫生物公司;辣根過氧化物酶標記的山羊抗大鼠IgG 北京中杉金橋生物有限公司;四甲基聯苯胺(TMB) Sigma公司;弗氏完全佐劑和不完全佐劑 Sigma公司,Tris Amresco公司;氯化鈉、固體硫酸銨、鹽酸、磷酸氫二鈉、磷酸二氫鈉試劑均為分析純 北京化工廠。
1.2 實驗方法
1.2.1 大米球蛋白的制備 干燥大米粉碎得到大米粉末,以濃度為1.0 mol/L,料液比1∶5(g/mL)的NaCl溶液鹽提,室溫震蕩提取16 h,8000 r/min離心20 min,上清液加入固體硫酸銨至溶液飽和度為50%,4 ℃冰箱鹽析,離心,沉淀以50 mmol/L Tris-HCl(pH9.0)緩沖液復溶,離心,上清調pH至4.02,4 ℃靜置過夜,離心,上清以20 mmol/L PBS(pH7.4,含0.15 mol/L NaCl)緩沖液透析并凍干[7]。
1.2.2 動物分組及處理 12只雌性BN大鼠隨機分為對照組和實驗組,每組6只[8-10]。實驗前采集BN大鼠血清作為對照。采取腹腔注射方式致敏,分別于第1、5、10 d分別腹腔注射1 mL 濃度為1 mg/mL大米過敏蛋白(含弗氏佐劑,大米過敏蛋白和弗氏佐劑體積比為1∶1;第一次致敏為弗氏完全佐劑,第二次和第三次致敏為弗氏不完全佐劑),第42 d注射大米過敏蛋白進行激發,對照組注射同等劑量的PBS緩沖液(pH7.4)。分別于第14、28、42 d內眥靜脈取血,離心分離血清,用于特異性抗體IgG、IgE和總IgE測定;于第14、21、35 d天內眥靜脈取血,加入抗凝劑EDTA-2K,離心分離血漿,用于測定組胺。
1.2.3 BN大鼠全身主動過敏反應評價 在第42 d對BN大鼠腹腔注射1 mL濃度為4 mg/mL的大米過敏蛋白進行激發,在激發實驗前后40 min分別測量各組BN大鼠體溫,計算BN大鼠在激發前后體溫變化;腹腔激發后,觀察各BN大鼠反應癥狀并參考何仁靜等[11]評分系統按照表1進行評分:
若平均反應達到2級即認為發生了過敏反應[12],其中平均過敏級數及過敏率按下列公式計算:
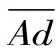
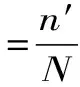
其中:n′:發生過敏的動物總數;RA:過敏率。
1.2.4 BN大鼠體重、免疫臟器指數測定 飼喂期間,每周稱體重兩次,計算平均體重,激發取血后處死BN大鼠,無菌取出胸腺和脾臟,計算免疫臟器指數[13]
1.2.5 BN大鼠免疫過程血液生化指標檢測 實驗第14、28、42 d抗凝管取血后,血漿上血球分析儀測定。
1.2.6 血清總IgE、特異性IgE、特異性IgG、血漿組胺含量測定 血清特異性IgG(IgE)測定:采用間接ELISA法進行檢測,用濃度為50 μg/mL的大米蛋白進行包被,4 ℃過夜;0.05% PBST洗滌三次,0.5%明膠37 ℃封閉1 h;洗滌三次,每孔加100 μL按1∶100稀釋的待測血清,37 ℃孵育1 h;洗滌三次,每孔加100 μL按1∶20000稀釋的HRP標記的山羊抗大鼠IgG或HRP標記山羊抗大鼠IgE),37 ℃保濕1 h;洗滌三次,每孔加100 μL底物顯色液,37 ℃反應15 min,每孔加50 μL l 2 mol/L H2SO4終止反應,用酶標儀在450 nm測OD值[14]。
血清總IgE測定和血漿組胺測定均按照ELISA試劑盒操作進行。
1.2.7 統計學分析 用Mircrosoft Excel 2007及SPSS20.0進行數據統計分析,組間檢驗采用t檢驗,p<0.01具有極顯著差異;p<0.05具有顯著差異。
2 結果與分析
2.1 BN大鼠全身主動過敏反應結果及體溫變化


表2 過敏反應結果與評分結果
注:對照組BN大鼠在第35 d取血后中因體弱死亡一只;實驗數據除第42 d對照組為5只動物數據外,其余均為6只動物實驗數據。
表5 不同時期BN大鼠血象水平變化(±s,N1=6,n1=5,n2=6)
注:**:與對照組比,p<0.01,差異極顯著;*:p<0.05,差異顯著,表6、表7同;對照組大鼠只數:第14、28 d N1=6,第42 d n1=5;實驗組大鼠只數:第14、28、42 d n2=6,表6、圖2、圖3同。由表3可知,激發前兩組BN大鼠的體溫沒有明顯差異,對照組體溫在激發前后也沒有明顯變化,而實驗組BN大鼠與激發前相比體溫卻明顯下降(p<0.01),由此說明大米蛋白致敏BN大鼠時會引起其體溫迅速下降。
表3 激發前后BN大鼠體溫變化(±s,n1=5,n2=6)
注:**:p<0.01時,顯著極差異;n1:對照組動物數;n2:實驗組動物數,圖1、表4同。
2.2 BN大鼠體重和免疫臟器指數變化
2.2.1 BN大鼠體重變化 實驗期間,每周稱量BN大鼠體重兩次取平均值,結果如圖1所示,兩組BN大鼠體重均較為平衡的增長,組間沒有顯著差異(p>0.05),可見大米過敏蛋白致敏BN大鼠對其體重沒有明顯的影響。
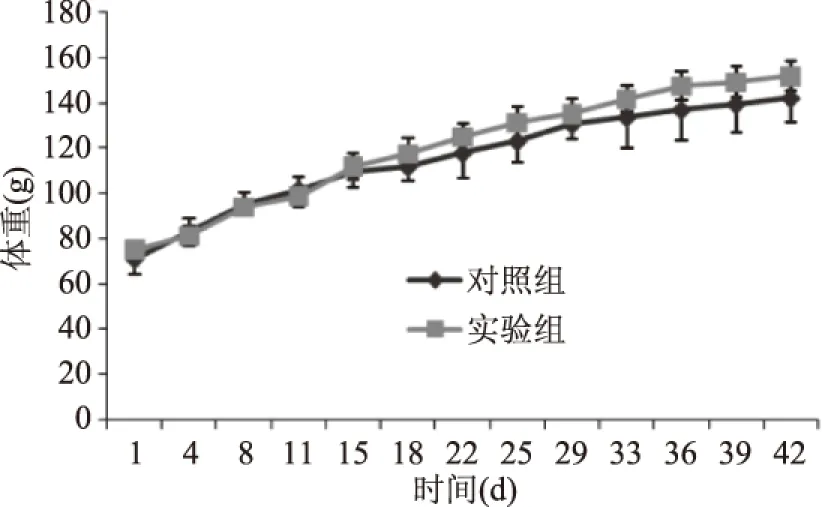
圖1 實驗期間各組BN大鼠體重變化(±s,n1=5,n2=6)Fig.1 Body weight changes of BN rats during the experiment(±s,n1=5,n2=6)
2.2.2 BN大鼠免疫臟器指數變化 如表4所示,兩組BN大鼠的胸腺指數無差異,而實驗組大鼠的脾臟指數卻有顯著升高(p<0.01)。過敏反應是機體對抗原強烈的免疫應答反應,免疫防御功能過高,易于發生過敏反應,大米過敏蛋白激發后BN大鼠的脾臟指數明顯增大說明大米蛋白已引發BN大鼠發生過敏反應[15]。
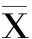
表4 大米過敏蛋白對BN大鼠脾臟指數
2.3 BN大鼠血象指標變化
過敏反應發生時,機體細胞會發生一系列應急反應,這些刺激會引起機體各類細胞、血小板等數目和種類發生變化。白細胞是機體抵御異物入侵的主要防線,由粒細胞、淋巴細胞和單核細胞組成。由表5可知,免疫第14 d,中性粒細胞數目極顯著增多,(p<0.01)單核細胞數目顯著增多(p<0.05),淋巴細胞數目變化幅度不大;第42 d,白細胞數目迅速上升(p<0.01),說明BN大鼠正處于過敏階段;食物過敏能引起嗜酸粒細胞增多,嗜酸粒細胞能夠拮抗和調節速發型超敏反應,具有吞噬能力,且能釋放組胺酶滅活組胺,抑制細胞脫顆粒[16]。但本實驗第14、28 d時嗜酸粒細胞并沒有明顯變化,只在第42 d嗜酸粒細胞數目有明顯升高。有研究表明,過敏反應期間存在不同的凝血因子紊亂,其中包括血小板的募集及活化,血小板可能作為抗原直接活化的效應細胞參與不同的超敏反應[17],在本實驗中,實驗第14 d,實驗組的血小板數量相比于對照組明顯升高(p<0.05),而第28 d與第42 d血小板的數量明顯下降,具體原因還有待進一步研究。
2.4 BN大鼠血清特異性IgG、總IgE和特異性IgE水平變化
表6 實驗期間BN大鼠血清總IgE變化(μg/mL)(±s,N1=6,n1=5,n2=6)
2.4.1 BN大鼠血清特異性IgG測定結果 不同時期大鼠血清特異性IgG水平變化如圖2所示,對照組在致敏不同時期血清特異性IgG無明顯變化;與對照組相比,實驗組BN大鼠在第14、28、42 d的血清特異性IgG抗體水平顯著升高(p<0.01)。
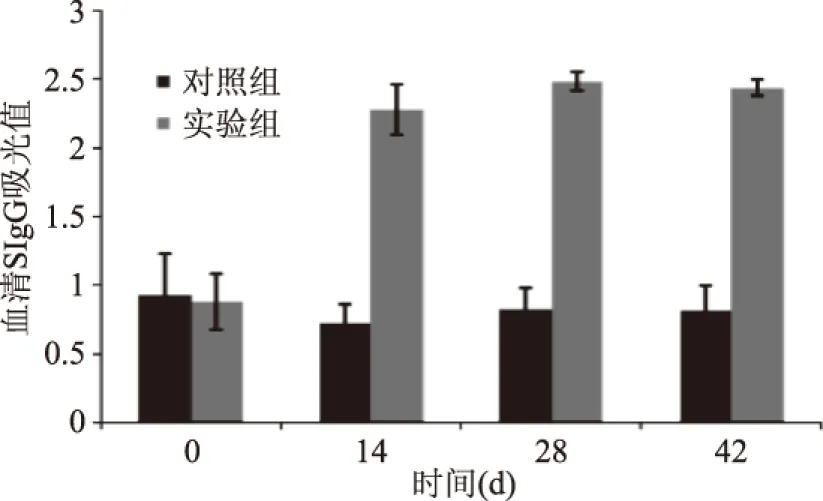
圖2 實驗期間BN大鼠血清特異性 IgG變化(±s,N1=6,n1=5,n2=6)Fig.2 Changes in Serum specific IgG of BN rats during the experiment(±s,N1=6,n1=5,n2=6)
2.4.2 BN大鼠血清總IgE測定結果 不同時期BN大鼠血清總IgE水平變化如表6所示,實驗第14、28 d實驗組BN大鼠血清總IgE抗體濃度與對照組相比顯著提高(p<0.05),42 d激發實驗后實驗組血清總IgE有極為顯著的提高(p<0.01),平均可達到4.032 μg/mL。IgE是介導Ⅰ型超敏反應(過敏反應)的主要抗體,包括總IgE和特異性IgE,總IgE水平的測定可用來鑒別超敏反應與非超敏反應[18]。實驗不同時期,實驗組與對照組相比均有顯著差異,說明BN大鼠發生了過敏反應。
2.4.3 血清特異性IgE測定結果 不同時期BN大鼠血清特異性IgE水平變化如圖3所示,血清特異性IgE抗體的測定,可以明確過敏反應是由哪種致敏原引起的(18(,實驗第14 d和42 d實驗組BN大鼠血清特異性IgE抗體濃度與對照組相比顯著提高(p<0.05),說明大鼠發生的過敏反應是由大米蛋白引起的,而實驗第28 d兩組BN大鼠血清特異性IgE抗體濃度無顯著差異,可能是由于過敏反會隨時間延長而逐漸弱化,導致血清特異性IgE抗體濃度降低;第42 d進行高濃度蛋白激發后,實驗組大鼠過敏反應劇烈,與對照組相比,血清特異性IgE抗體濃度顯著升高(p<0.05)。
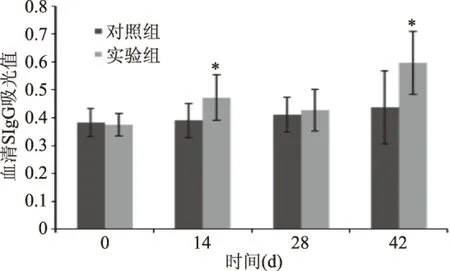
圖3 實驗期間BN大鼠血清特異性 IgE變化(±s,N1=6,n1=5,n2=6)Fig.3 Changes in Serum specific IgG of BN rats during the experiment(±s,N1=6,n1=5,n2=6)
2.5 組胺測定結果
由表7可知,在致敏第14、21、35 d,實驗組BN大鼠血漿組胺釋放量均顯著高于對照組(p<0.05)。
表7 實驗期間BN大鼠組胺變化(μg/L)(±s,n=6)
3 結論與討論
動物模型是食物致敏原檢測最直接的辦法,迄今為止報道過的用于食物過敏的動物有狗、幼豬、豚鼠、Balb/c小鼠、C3H/HeJ小鼠、BN大鼠等[19],常用的致敏方式有灌胃、腹腔注射和皮下注射,這三者均有優缺點。目前國際上尚未建立標準的食物過敏動物模型。理想的過敏動物模型,不僅僅是動物暴露于食物致敏原后產生過敏反應,在暴露于無致敏史食物時也應不產生過敏反應,且過敏癥狀應與人類相似[20]。經研究發現,BN大鼠在致敏原刺激下,能產生較高水平的特異性IgE、IgG抗體,具有較強的致敏免疫反應[21],且相比于灌胃,腹腔注射能夠克服口服耐受的缺點,是評價食物致敏原合適的動物模型。Nakase[22]、Hidehiko[23]、Yumiko[24]等對大米過敏蛋白進行過研究,結果表明相對分子質量為14~16 ku和 33 ku的大米蛋白為致敏蛋白,可引起IgE介導的過敏反應。本實驗以提取的大米過敏蛋白為致敏原致敏大鼠,通過全身主動過敏反應判斷過敏反應發生率為100%,且脾臟指數明顯增大,嗜酸粒細胞數目明顯升高。綜合以上指標初步確定大米過敏動物模型建立成功。
動物體內的特異性抗體IgG、總IgE和特異性IgE水平與致敏原敏感性相關。在食物過敏中,特異性抗體IgG的出現,往往提示該蛋白質具有免疫原性[25]。在本實驗中對sIgG抗體檢測,實驗組抗體水平明顯高于對照組,驗證了大米蛋白首先具有免疫原性,而血清總IgE、sIgE抗體和組胺水平的檢測證明該大米蛋白不僅具有致敏性,且能引起BN大鼠發生過敏反應,這也與陳如程等[14]的研究相一致。然而,同一物種不同個體對過敏原的過敏性存在差異,不同物種間差異更大,僅用動物模型來評價大米蛋白的過敏性仍存在一定的局限,因此在今后的研究中將利用大鼠RBL-2H3細胞等多種模型進一步評價大米蛋白的過敏性。
[1]李 明,李赟高,高紅艷. 大米蛋白研究進展[J]. 糧食與油脂,2006,(8):3-5.
[2]李海飛,高金燕,袁娟麗,等. 大米過敏蛋白的研究進展[J]. 食品科學,2014,35(23):308-312.
[3]CHEN XiaoWei,Ken Wan-Keung Lau,YANG fan,et al. An adjuvant free mouse model of oral allergenic sensitization to rice seeds protein[J]. BMC Gastroenterology,2011,11:62.
[4]黃建芳,王彩霞,向軍儉. 食物過敏動物模型的研究進展[J]. 食品科學,2014,35(3):280-284.
[5]孫拿拿. 轉基因食品致敏性評價嚙齒類動物模型(BN大鼠和C3H/HeJ小鼠)研究[D]. 北京:中國農業科學院,2010.
[6]農業部1782號公告-13-2012. 轉基因生物及其產品食用安全檢測挪威棕色大鼠致敏實驗方法[S].
[7]Tsukasa Matsuda,Makoto Sugiyama,etc. Purification and Properties of an Allergenic Protein in Rice Grain[J]. Agric. Food Chem.,1988,52(6):1465-1470.
[8]Tsukasa matsuda Rieko nomura,Makoto Sugiyama,et al. Immunochemical Studies on Rice Allergenic Proteins[J]. Agricultural and Biological Chemistry,1991,55(2):S9-S13.
[9]H. A. C. Atkinson,I. T. Johnsonf,J. M. Geet,F. Griroriadou,et al. Brown Norway Rat Model of food allergy:Effect of Plant components on the Development of Oral Sensitization[J]. Food & Chemical Toxicology,1999,34(1):27-32.
[10]Hiroshi Akiyama,Reiko Teshima,Jun-ichiro Sakushima,et al. Examination of oral sensitization with ovalbumin in Brown Norway rats and three strains of mice[J]. Immunology Letters,2001,78:1-5.
[11]何靜仁. 銀杏酸的變應原性及致過敏作用機制研究[D]. 武漢:華中農業大學,2003
[12]楊劍婷. 白果過敏蛋白及其致敏機理的研究[D]. 南京:南京林業大學,2003.
[13]王思蘆,汪開毓,趙玲,等. 雞樅菌多糖對免疫抑制小鼠免疫功能的影響[J]. 中國藥理學通報,2013,29(1):59-63.
[14]陳如程,李娜,樊柏林,等. BN大鼠致敏動物模型研究[M]. 中國食品衛生雜志,2014,26(1):14-17.
[15]夏春麗,舒剛,李艷華. 幾種補益方劑對雛雞生長性能和免疫臟器指數的影響(A]. 安全優質的家禽生產—第十五次全國家禽學術討論會論文集[C].廣東:華南理工大學出版社,2011.522-526.
[16]龔非力. 醫學免疫學[M]. 北京:科學出版社,2009.113.
[17]王德錄,吳學忠. 過敏反應中血小板功能的研究進展[J]. 臨床輸血與檢驗,2012,14(1):92-94.
[18]劉俊紅. 食物過敏幼鼠模型的建立與評價[D]. 福建:福建醫科大學,2012.
[19]黃建芳,王彩霞,向軍儉,等. 蝦過敏C57/BL6 小鼠動物模型的建立[J]. 食品工業科技,2012,33(23):358-361.
[20]孫佳益,王錫昌,李振興,等. 蝦類原肌球蛋白的Balb/c小鼠致敏動物模型構建研究[J]. 免疫學雜志,2013,29(2):161-164.
[21]孫拿拿,張倩男,賈旭東,等. BN 大鼠經口致敏動物模型研究[J]. 中國食品衛生雜志,2013,25(3):214-217.
[22]Masayuki Nakase,Takahiro Adachi,etc,et al. Rice(Oryza sativa L.)α-amylase inhibitors of 14-16 kDa are potential Allergens and Products of a Multigene Family[J]. Journal of Agricultural and Food Chemistry,1996,44(9):2624-2628.
[23]Hidehiko Izumi,Minoru Sugiyama. Structural characterization of the 16-kDa allergen,RA17,in rice seeds. Prediction of the secondary structure and identification of intramolecular disulfide bridges.[J]. Bioscience,Biotechnology,and Biochemistry,1999,63(12):2059-2063.
[24]Yumiko Usui,Masayuki Nakase. A 33-kDa Allergen from Rice(Oryza sativa L. Japonica)cDNA cloning,expression,and Identification as a novel glyoxalase[J]. The Journal of Biological Chemistry,2001,6(14):11376-11381.
[25]陳晨. 轉基因食品致敏性評價Balb/c小鼠模型和RBL-2H3細胞模型研究[D].北京:中國疾病預防控制中心,2013.
Establishment of a rice protein sensitization animal model based on BN rat
WANG Yi-xia,YIN Man,MA Tao,LU Jun,CAI Mu-yi,GU Rui-zeng*
(Beijing Engineering Research Center of Protein & Functional Peptides,China National Research Institute of Food and Fermentation Industries,Beijing 100015,China)
In order to establish a sensitization animal model of rice protein,the sensitinogen was intraperitoneal injected to female Brown-Norway(BN)rat. The general allergic reactions and the changes of weight,body temperature,immune organ index were observed,the level of antibody(specific IgG and IgE,total IgE)in sera,histamine and blood picture in plasma were measured to verify the animal model. Results showed that the general allergic reactions in each rat of experimental group were positive,the body temperature was decreased and the spleen index was increased significantly(p<0.01),and white blood cells and acidophilic granulocyte number were increased(p<0.05)after stimulated by rice protein at the time of 42nd day. Meanwhile,the level of specific IgG was increased significantly(p<0.01),total IgE and plasma histamine was increased(p<0.05)compared to the control group,on the 14th and 42nd days,the level of specific IgE was increased while it did not show obvious difference at the time of 28th day(p<0.05). This study indicates that the Brown Norway(BN)rat is a suitable animal model for evaluating rice protein allergenicity.
rice protein;food allergen;animal model;antibody;histamine
2016-08-19
王一俠(1990-),女,碩士,研究方向:食物過敏原,E-mail:wyx0105@126.com。
*通訊作者:谷瑞增(1973-),男,本科,教授級高級工程師,研究方向:食源性低聚肽,E-mail:guruizeng@163.com。
國家高技術研究發展計劃(863 計劃)項目(2013AA102205-02);十三五國家重點研發計劃(2016YFD0400604)。
TS201.4
A
1002-0306(2017)05-0345-05
10.13386/j.issn1002-0306.2017.05.057
猜你喜歡
世界科學技術-中醫藥現代化(2022年9期)2023-01-17 07:31:10
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
世界科學技術-中醫藥現代化(2021年5期)2021-11-05 06:54:30
中老年保健(2021年3期)2021-08-22 06:50:04
天津醫科大學學報(2021年2期)2021-03-29 05:31:08
現代臨床醫學(2021年1期)2021-01-26 00:56:02
世界科學技術-中醫藥現代化(2020年2期)2020-07-25 02:06:22
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
