菊花CmAP1基因克隆及其植物表達載體構建
2017-05-30 19:48:01梁芳黃萍袁秀云崔波李霞
南方農業學報 2017年6期
梁芳 黃萍 袁秀云 崔波 李霞


摘要:【目的】克隆菊花CmAPETALA1基因(CmAP1)并構建植物表達載體,為該基因功能的遺傳改良打下研究基礎。【方法】采用同源克隆及RT-PCR從菊花葉片中克隆CmAP1基因,運用在線生物信息學分析工具對其核苷酸及氨基酸序列進行分析,利用實時熒光定量PCR對該基因的組織表達差異進行分析,并用雙酶切法將目的基因與pCAMBIA1300載體連接,構建植物表達載體。【結果】克隆獲得的CmAP1基因(GenBank登錄號KU872999)編碼區開放閱讀框(ORF)長741 bp,編碼246個氨基酸;CmAP1蛋白屬親水性蛋白,分子質量約28.3 kD、理論等電點(pI)為8.76,預測該蛋白內含10個磷酸化位點和2個潛在的N-糖基化位點;蛋白氨基酸序列比對和系統發育進化分析結果表明,CmAP1蛋白與CDM111蛋白的同源性達98%,屬MADS-box基因家族A類基因的euAP1分支;實時熒光定量PCR分析結果表明,CmAP1基因在花蕾中表達量極高,在舌狀花和筒狀花中表達量較低,而在營養器官中僅痕量表達。將CmAP1基因連接到pCAMBIA1300載體上,可成功構建植物表達載體pCAMBIA1300-CmAP1。【結論】CmAP1基因在花的發育及形成過程中發揮重要作用,成功構建獲得的植物表達載體pCAMBIA1300-CmAP1可為后期利用基因工程手段改變菊花花形態結構、調控花期及遺傳改良打下基礎。
關鍵詞: 菊花;CmAPETALA1(CmAP1);基因克隆;植物表達載體;生物信息學
中圖分類號: S682.11 文獻標志碼:A 文章編號:2095-1191(2017)06-0000-08
Cloning of CmAP1 gene from Chrysanthemum morifolium and establishment of its plant expression vector
Abstract:【Objective】CmAPETALA1 gene(CmAP1) in Chrysanthemum morifolium was cloned and its plant expression vector was established, in order to provide references for further study on genetic improvement of CmAP1 gene function. 【Method】CmAP1 gene was cloned from leaf of C. morifolium by method of homology cloning and RT-PCR. The nucleotide and amino acid sequence were analyzed by online bioinformatics tool. Expression difference of this gene was determined by real-time fluorescence quantitative PCR. CmAP1 gene was linked to pCAMBIA1300 vector by double enzyme digestion to establish plant expression vector. 【Result】CmAP1 gene(GenBank accession number:KU872999) was obtained by cloning. The open reading frame(ORF) of CmAP1 was 741 bp encoding 246 amino acids. CmAP1 protein was a hydrophilic protein,and the molecular weight of this protein was about 28.3 kD. Its theoretical isoelectric point(pI) was 8.76. It was predicted that the protein contained ten phosphorylation sites and two potential N-glycosayation sites. Sequence and phylogenic analysis showed that CmAP1 protein was close to C. morifolium CDM111 protein with 98% similarity, which both belonged to euAP1 clade of A type gene in MADS-box gene family. Real fluorescence quantitative PCR analysis indicated that expression of CmAP1 gene was extremely high in buds. The expression level was low in tubiform and lingulate florets and was in trace expression in vegetative organs. CmAP1 was linked to pCAMBIA1300 vector and the plant expression vector pCAMBIA1300-CmAP1 was established successfully. 【Conclusion】CmAP1 gene plays an important role in growth and development of C. morifolium. The established plant expression vector pCAMBIA1300-CmAP1 is useful for further research on changing morphological structure of C. Morifolium by genetic engineering, regulation of flowering and genetic modification.
Key words: Chrysanthemum morifolium; CmAPETALA1(CmAP1); gene cloning; plant expression vector; bioinformatics
0 引言
【研究意義】成花過程是植物從營養生長向生殖生長轉折的關鍵期。調控植物成花過程的基因分為花序分生組織特征基因、花分生組織特征基因和花器官形態特征基因。大部分花器官形態特征基因屬于MADS-box轉錄因子家族基因,其中APETALA1/FRUITFULL(AP1/FUL)亞家族被認為既是花分生組織特征基因,又是花器官特征基因。AP1基因是花瓣和萼片正常發育的必需基因(Alvarez-Buylla et al.,2006)。菊花(Chrysanthemum morifolium)是重要的短日照作物之一,多數品種在自然條件下于每年10~11月開花,溫室種植的菊花需人工調控光照周期,成本較高,而通過分子育種可縮短菊花對短日照的響應時間,有效降低溫室供熱及人力物力等生產成本。因此,克隆菊花AP1基因(CmAP1)并進行植物表達載體構建對其基因功能的遺傳改良具有重要意義。【前人研究進展】迄今,已有很多植物的AP1同源基因被分離克隆,如馬鈴薯(樊春媛等,2010)、草莓(鄒冬梅等,2012)、萼脊蘭(崔波等,2013)、梨(Liu et al.,2013)、荷花(Kong et al.,2015)等。Ellul等(2004)將擬南芥AP1基因轉入番茄中,轉基因番茄營養生長期縮短,開花期比對照提前,且不影響植株正常發育。煙草中異源表達大豆AP1基因,表現為提前開花及花器官改變(Chi et al.,2011)。白樺AP1基因過表達能引起早花,且影響很多開花相關基因的表達及二萜化合物的生物合成(Huang et al.,2015)。在菊花MADS-box基因研究方面,Shchennikova等(2004)從菊花中分離出3個AP1/FUL亞家族基因CDM111、CDM8和CDM41及1個SEPALLATA3亞家族基因CDM44;Shulga等(2011)將CDM111、HAM75和HAM92基因分別轉入菊花中,在長日照條件下轉基因植株與對照在表型上無差異,短日照條件下花芽啟動比對照提前兩周,且轉基因植株花色著色更早;Shchennikova等(2011)克隆出菊花AGAMOUS的同源基因CDM37,該基因能促進萼片中蜜腺的發育。此外,將菊花CDM41(Goloveshkina et al.,2012)、CIM8(Wang et al.,2013)和SOC1(Fu et al.,2014)基因分別轉入煙草和擬南芥中,均可使煙草和擬南芥提前開花且引起花形態異常。【本研究切入點】目前,針對AP1同源基因克隆和生物信息學分析及植物表達載體構建的研究鮮見報道。【擬解決的關鍵問題】以菊花晚花品種獅子頭為試驗材料,利用RT-PCR克隆CmAP1基因,并對該基因核苷酸及其推導氨基酸序列進行生物信息學分析,利用實時熒光定量PCR對開花期菊花不同器官中該基因的表達量進行定量分析,構建植物表達載體pCAMBIA1300-CmAP1,以期為CmAP1基因功能的遺傳改良提供參考依據。
1 材料與方法
1. 1 試驗材料
1. 1. 1 植物材料 供試菊花品種為黃色大花品種獅子頭,由鄭州好日子園藝有限公司提供;試驗于2015年10月~2016年4月在鄭州師范學院生物工程研究所進行。取其葉片進行基因克隆,在盛花期分別取健康植株的根、莖、葉、花蕾、管狀花及舌狀花,用于分析目的基因在不同部位的表達量。所有材料迅速用液氮冷凍,-80 ℃保存備用。
1. 1. 2 菌株和質粒 大腸桿菌(Escherichia coli)DH5α、根癌農桿菌(Agrobactrium tumefaciens)EHA105及植物表達載體pCAMBIA1300由鄭州師范學院生物工程研究所分子實驗室保存提供,pGEM-T Easy Vector購于寶生物工程(大連)有限公司。
1. 1. 3 酶與化學試劑 XbaⅠ及SmaⅠ限制性內切酶、rTaq DNA聚合酶、T4 DNA連接酶、dNTPs、DNA Marker等購自寶生物工程(大連)有限公司。瓊脂糖凝膠回收試劑盒、質粒提取試劑盒購自天根生化科技(北京)有限公司。
1. 2 試驗方法
1. 2. 1 引物設計 運用Primer Premier 5.0和DNAMAN,根據NCBI上已登錄的菊科植物AP1基因序列ORF兩端設計引物CmAP1-F和CmAP1-R(表1);為構建真核表達載體,在引物5'端分別加上Xba I和Sma I酶切位點。
1. 2. 2 CmAP1基因克隆 采用Trizol(Invitrogen公司)試劑提取葉片總RNA。以提取的總RNA反轉錄合成的cDNA第一鏈為模板,根據設計合成的引物進行PCR擴增。PCR反應體系20.0 μL:ddH2O 13.2 μL、10×PCR Buffer 2.0 μL、dNTP(2.5 mmol/L)1.6 μL、CmAP1-F和CmAP1-R(10 mmol/L)各1.0 μL、cDNA 1.0 μL、rTaq DNA聚合酶0.2 μL。擴增程序:95 ℃預變性5 min;95 ℃ 40 s,55 ℃ 40 s,72 ℃ 40 s,進行35個循環;72 ℃延伸10 min。PCR產物用1%瓊脂糖凝膠檢測及回收,目的片段大小為756 bp。將回收的目的片段再經凝膠電泳檢測正確后,連接到pGEM-T載體上,然后轉入DH5α感受態細胞中,經藍白斑篩選及菌落PCR鑒定,陽性菌液送至上海英駿生物技術有限公司測序。
1. 2. 3 實時熒光定量PCR 根據克隆獲得的CmAP1基因序列,重新設計實時熒光定量PCR引物qRT-F和qRT-R,以Actin基因(GenBank登錄號AB770471)為內參基因,設計引物(表1)對CmAP1基因在不同組織部位的表達進行分析。分別提取根、莖、葉、花蕾、管狀花及舌狀花的總RNA,用PrimeScript RT Reagent Kit with gDNA Eraser試劑盒(TaKaRa)反轉錄合成cDNA第一鏈。采用SYBR Premix Ex TaqTM Ⅱ Kit(TaKaRa)進行實時熒光定量PCR擴增,反應體系20.0 μL:2×SYBR Premix Ex TaqTM Ⅱ10.0 μL、qRT-F和qRT-R各0.8 μL、cDNA 1.0 μL,ddH2O 7.4 μL。擴增程序:95 ℃預變性30 s;95 ℃ 15 s,58 ℃ 15 s,72 ℃ 15 s,進行40個循環。每個樣品3次重復,并設陰性對照。CmAP1相對表達量Rel.Exp=2-△△C t,其中△C t=C t(CmAP1)-
C t(Actin),△△C t=(各植物組織△C t)-(根△C t)。
1. 2. 4 CmAP1過表達載體構建 將測序正確的陽性菌液擴搖并進行PCR擴增,提取質粒pGEM- CmAP1,經限制性內切酶Xba I和Sma I酶切后,回收約750 bp的目的片段;同時pCAMBIA1300載體也用Xba I和Sma I進行酶切,回收大片段。將回收的目的片段和表達載體片段用T4 DNA連接酶于16 ℃下連接過夜,連接產物轉化DH5α感受態細胞,篩選陽性菌落,重組質粒命名為pCAMBIA1300-CmAP1。提取質粒后進行雙酶切鑒定,以確定植物表達載體構建成功,正確者將其質粒重組DNA轉化農桿菌EHA105感受態細胞,28 ℃培養2 d后,挑取單菌落進行兩次PCR鑒定,均為陽性者確定為陽性轉化子,-80 ℃保存或用于后續的農桿菌侵染試驗。
1. 2. 5 生物信息學分析 利用ProtParam(http://web.expasy.org/protparam/)對CmAP1蛋白的理化性質進行分析,用Protscale(http://web.expasy.org/protscale.html/)對蛋白親疏水性進行分析;通過NCBI的CDD數據庫對蛋白序列進行結構域分析(http://www.ncbi.nlm.nih.
gov/Structure/cdd/wrpsb.cgi)。采用NetPhos 2.0 Server(http://www.cbs.dtu.dk/services/NetPhos/)和NetNGlyc 1.0 Server(http://www.cbs.dtu.dk/services/NetNGlyc/)分別分析磷酸化位點和N-糖基化位點。用ExPASyGOR4 Secondary Structure Prediction Method(https://npsa-
prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_gor4.html)對蛋白的二級結構進行預測,利用SWISS-
MODEL(https://swissmodel.expasy.org/)同源建模方法對該蛋白三級結構進行預測;利用DNAMAN進行蛋白同源序列多重比對分析。從NCBI下載已知菊科所有MADS-box A類基因,采用ClustalX進行核酸序列比對,然后采用MEGA 6.0以鄰接法構建基于CmAP1基因的系統發育進化樹。
2 結果與分析
2. 1 CmAP1基因ORF的克隆
以菊花葉片cDNA為模板,利用引物對CmAP1-F和CmAP1-R進行PCR擴增,得到預期片段(756 bp),其ORF長741 bp,編碼246個氨基酸(圖1)。在NCBI上進行BLASTN在線分析,發現此序列與其他已登錄菊花AP1基因(AY173054、AB808650、AB808649、JX87-
2512、AB808648、AB808647和AB679275)的同源性均為99%,與向日葵(Helianthus annuus)AP1基因(AY173071、GU985582、GU985583和AF462152)的同源性為89%~90%,與非洲菊(Gerbera hybrid cultivar)AP1基因(AJ009727和FN298388)的同源性為83%~87%,故將該基因命名為CmAP1,GenBank登錄號KU872999。
2. 2 CmAP1蛋白的基本理化性質
CmAP1蛋白氨基酸序列的CDD分析結果(圖2)顯示,CmAP1蛋白氨基酸序列具有典型的MEF2_like MADS結構域和K-box結構域,MADS結構域位于CmAP1蛋白N端第2~75位氨基酸,K-box結構域位于第89~167位氨基酸,說明CmAP1基因是植物典型的MIKCC-Type MADS-box基因。
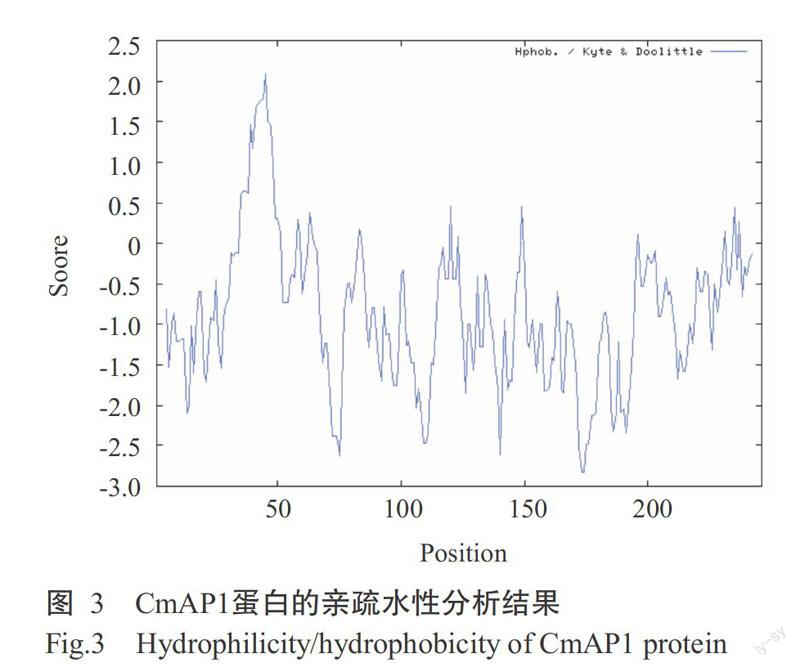
基于K-D法的CmAP1蛋白親疏水性預測結果見圖3,正值為疏水區,負值為親水區,多肽鏈第36~52位的氨基酸區域為疏水性最強區域,第46位的異亮氨酸(Ile)疏水性最強,分值最高,為2.089;第173位的谷氨酸(Glu)和第174位的精氨酸(Lys)分值最低,均為2.833,親水性最強。整條多肽鏈中親水性氨基酸較多,故表現為親水性。
ProtParam分析結果表明,CmAP1分子質量約28.3 kD、理論等電點(pI)為8.76,分子式為C1227H1976N362O385S11;負電荷殘基總數(Asp+Glu)為33,正電荷殘基總數(Arg+Lys)為37。在組成CmAP1蛋白的20種氨基酸中,亮氨酸(Leu)所占比例最高,為10.6%,色氨酸(Trp)所占比例最低,為0.8%。該蛋白的不穩定指數為52.08,半衰期30 h,脂肪指數為73.33,根據Guruprasad判定,CmAP1蛋白不穩定。NetPhos 2.0 Server預測結果表明,CmAP1蛋白序列中存在10個潛在的磷酸化位點,包括6個Ser位點(22Ser、61Ser、74Ser、89Ser、99Ser和120Ser),3個Thr位點(76Thr、86Thr和168Thr)和1個Tyr位點(191Tyr)。NetNGlyc 1.0 Server預測結果表明,CmAP1蛋白存在2個潛在的N-糖基化位點,分別為164Asn和214Asn。說明蛋白翻譯后修飾對CmAP1蛋白功能的實現起重要作用。
2. 3 CmAP1蛋白二級和三級結構的預測分析結果
采用GOR4進行CmAP1蛋白二級結構預測,結果表明,CmAP1蛋白二級結構由α螺旋、伸展鏈和無規則卷曲組成,其中,α螺旋(Hh)占54.88%,β折疊(Ee)占15.04%,無規則卷曲(Cc)占30.08%(圖4)。采用SWISS-MODEL同源建模方法對CmAP1蛋白三級結構進行預測,以3p57.1.A為模板,序列一致性為50.68%,三級結構詳見圖5,即CmAP1蛋白由α螺旋、β折疊和無規則卷曲組成。
2. 4 CmAP1蛋白氨基酸序列比對及系統發育分析結果
CmAP1蛋白與菊科其他AP1蛋白氨基酸序列的比對分析結果(圖6)表明,CmAP1蛋白序列與甘野菊(C. seticuspe,BAL14662)的序列完全相同,與向日葵HAM75(Helianthus annuus,AAL83209)蛋白的同源性為93%,與菊花CDM111(AAO22979)的同源性為98%,與非洲菊GSQUA3(Gerbera hybrid cultivar,CAX65662)的同源性為88%。
為進一步分析CmAP1基因與菊科及模式植物擬南芥MADS-box A類基因的進化關系,構建基于CmAP1基因的系統發育進化樹(圖7)。從圖7可看出,所有的CmAP1基因可分為三大分支,甘野菊的CsAFL1基因(AB679273)和CsM8基因(AB770472)、菊花的CmAFL1基因(AB451218)和cdm8基因(AY173056)、甘菊(C. lavandulifolium)的AP1-like基因(HM130686)及擬南芥AGL8基因(ATU33473)同屬euFUL分支;甘野菊的AP1/FUL-like基因(AB839770)、菊花的cdm41基因(AY173055)和CmFUL基因(KT894379)及擬南芥AGL79基因(NM_113925)同屬FUL-like分支;其他菊花AP1基因則與擬南芥AP1基因(NM_105581)及CAL基因(NM_102395)同屬euAP1分支。在euAP1分支中,向日葵的3個euAP1基因(AF462152、GU985592和GU985583)自成一個小分支,另一個分支則全部為菊科的euAP1類基因,說明euAP1基因的進化具有明顯種屬特異性。
2. 5 CmAP1基因在不同組織部位的表達情況
用Trizol法提取各組織部位的總RNA,以Actin基因為內參基因,利用實時熒光定量PCR對菊花花期不同組織部位CmAP1基因的表達量進行分析,結果顯示,各溶解曲線峰單一;CmAP1基因在花蕾中表達豐度最高,其次是管狀花和舌狀花,在根莖葉中均為痕量表達(圖8)。說明CmAP1基因可能在花的發育及形成中發揮重要作用。
2. 6 CmAP1基因植物表達載體的構建
將測序成功的質粒pGEM-CmAP1經Xba I和Sma I限制性內切酶酶切后,得到兩個目的片段,分別為756和約3000 bp(圖9),切膠回收小片段;質粒pCAMBIA1300經Xba I和Sma I酶切后,切膠回收大片段。將回收的pCAMBIA1300大片段與pGEM-CmAP1的小片段用T4 DNA連接酶連接,然后轉入DH5α感受態細胞。陽性轉化子經質粒酶切鑒定,得到一條長756 bp的小片段和一條長約10000 bp的pCAMBIA1300載體片段(圖10);測序驗證結果表明,植物表達載體pCAMBIA1300-CmAP1構建成功。用凍融法將pCAMBIA1300-CmAP1質粒轉化農桿菌EHA105感受態細胞,28 ℃培養2 d后,挑取單菌落進行PCR鑒定。電泳結果(圖11)顯示,約在750 bp處有很亮的目的條帶,說明轉化子為陽性,CmAP1基因已成功轉入農桿菌中。
3 討論
通過同源克隆從菊花中克隆獲得的AP1同源基因(命名為CmAP1),其ORF長741 bp,編碼246個氨基酸;CmAP1蛋白屬于親水性蛋白,分子質量約28.3 kD、理論等電點(pI)為8.76,預測該蛋白內含10個磷酸化位點和2個潛在的N-糖基化位點;利用SWISS- MODEL同源建模方法對CmAP1蛋白三級結構進行預測,以3p57.1.A為模板,二者序列一致性為50.68%,表明CmAP1蛋白由α螺旋、β折疊和無規則卷曲組成;氨基酸序列比對分析結果表明,CmAP1基因屬于MADS- box的AP1/FUL亞家族,與菊花CDM111序列一致性為98%,與向日葵HAM75序列一致性為93%。Litt和Irish(2003)、Shan等(2007)研究認為,AP1/FUL-like基因經過長期進化和基因復制產生euAP1、euFUL和FUL-like(AGL79)三大分支,本研究中的CmAP1基因屬于其中euAP1基因進化枝,euAP1類基因的進化具有明顯的種屬特異性,表明AP1基因在進化上具有高度保守性。Becker和Thei?茁en(2003)、Kaufman等(2005)研究表明,MADS-box基因依據進化及結構可分為Ⅰ型和Ⅱ型,Ⅱ型又分為動物和真菌的MEF2-like型和植物的MIKC型,多數已知植物的MADS-box基因均屬于MIKCC型,其編碼蛋白具有典型M、I、K和C結構。本研究通過對CmAP1蛋白進行在線分析,結果表明該蛋白屬于植物MADS-box超家族,具有該家族特有的MADS_MEF2_like高度保守結構域及半保守K-box域,MADS_MEF2_like域是序列特異的DNA結合基元,K-box域是蛋白分子間相互作用的區域(Yang and Jack,2004);C端可變最不保守,但MADS-box蛋白常含有一些保守基序,這些基序在蛋白復合體的形成和轉錄激活中起著重要作用(Honma and Goto,2001),同時C端不同的基序代表了MADS-box蛋白不同的功能(Lamb and Irish,2003),CmAP1基因在進化上屬于euAP1分支,euAP1類基因編碼的氨基酸序列在C末端具有典型的euAP1基序,常以CFAA結尾。
本研究運用實時熒光定量PCR對菊花花期CmAP1基因在不同部位的表達進行分析,結果表明,CmAP1基因在營養器官中表達量很低,在花蕾中表達量最高,在舌狀花和管狀花中表達量無明顯差異。梁芳等(2016)研究發現,菊花品種獅子頭CmFUL基因在管狀花和舌狀花中表達模式明顯不同,在管狀花中的表達量是舌狀花中的12倍,表明同為AP1/FUL亞家族基因其功能存在差異。本研究構建了以CaMV 35S為啟動子、GUS為報告基因、含有CmAP1目的基因的植物表達載體pCAMBIA1300-CmAP1,經雙酶切及PCR鑒定確定CmAP1基因已成功轉入農桿菌EHA105中。
4 結論
CmAP1基因在花的發育及形成過程中發揮重要作用,成功構建獲得的植物表達載體pCAMBIA1300- CmAP1可為后期利用基因工程手段改變菊花花形態結構、調控花期及遺傳改良打下基礎。
參考文獻:
崔波,蔣素華,劉佳,田云芳,袁秀云,馬杰. 2013. 萼脊蘭AP1-like基因的克隆與表達分析[J]. 西北植物學報,33(9):1739-1744. [Cui B,Jiang S H,Liu J,Tian Y F,Yuan X Y,Ma J. 2013. Cloning and expression analysis of AP1-like gene from Sedirea japonica[J]. Acta Botanica Boreali-Occidentalia Sinica,33(9):1739-1744.]
樊春媛,印敬明,王冰,韋青,李根濤,張云峰,楊清. 2010. 馬鈴薯StAP1基因的分子克隆與表達分析[J]. 植物生理學通訊,46(9):933-938. [Fan C Y,Yin J M,Wang B,Wei Q,Li G T,Zhang Y F,Yang Q. 2010. Molecular cloning and expression analysis of a potato StAP1 gene[J]. Plant Physiology Communications,46(9):933-938.]
梁芳,許申平,蔣素華,袁秀云,崔波,張靜. 2016. 菊花MADS-box基因的克隆與表達載體構建[J]. 華北農學報,31(3):25-31. [Liang F,Xu S P,Jiang S H,Yuan X Y,Cui B,Zhang J. 2016. Cloning,expression and vector construction of a MADS-box gene in Chrysanthemum morifo-
lium[J]. Acta Agriculturae Boreali-Sinica,31(3):25-31.]
鄒冬梅,劉月學,張志宏,李賀,馬躍,代紅艷. 2012. 草莓AP1同源基因的克隆、表達及啟動子分析[J]. 中國農業科學,45(10):1972-1981. [Zou D M,Liu Y X,Zhang Z H,Li H,Ma Y,Dai H Y. 2012. Cloning,expression and promo-
ter analysis of AP1 homologous gene from strawberry(Fragaria×ananassa)[J]. Scientia Agriculture Sinica,45(10):1972-1981.]
Alvarez-Buylla R E,Garcl?觃-Ponce B,Garay-Arroyo A. 2006. Unique and redundant functional domains of APETALA1 and CAULIFLOWER,two recently duplicated Arabidopsis thaliana floral MADS-box genes[J]. Journal of Experimental Botany,57(12):3099-3107.
Becker A,Theiβen G. 2003. The major clades of MADS-box genes and their role in the development and evolution of flowering plants[J]. Plant Molecular Evolution,29(3):464-489.
Chi Y J,Huang F,Liu H C,Yang S P,Yu D Y. 2011. An APETALA1-like gene of soybean regulates flowering time and specifies floral organs[J]. Journal of Plant Physiology,(168):2251-2259.
Ellul P,Angosto T,García-Sogo B,García-Hurtado N,Martín- Trillo M,Salinas M,Salinas V,Lozano R,Martínez-Zapater M J. 2004. Expression of Arabidopsis APETALA1 in tomato reduces its vegetative cycle without affecting plant production[J]. Molecular Breeding,13(2):155-163.
Fu J X,Qi S,Yang L W,Dai Y,Dai S L. 2014. Characterization of Chrysanthemum ClSOC1-1 and ClSOC1-2,homo-
logous genes of SOC1[J]. Plant Molecular Biology Reporter,32(3):740-749.
Goloveshkina N E,Shchennikova V A,Kamionshaya M A,Skryabin G K,Shulga A O. 2012. Influence of ectopic expression of Asteraceae MADS box genes on plant ontogeny in tobacco[J]. Plant Cell Tissue and Organ Culture,109(1):61-71.
Honma T, Goto K. 2001. Complexes of MADS-box proteins are sufficient to convert leaves into floral organs[J]. Nature,409(6819):525-529.
Huang H J,Chen S,Li H Y,Jiang J. 2015. Next-generation transcriptome analysis in transgenic birch overexpressing and suppressing APETALA1 sheds lights in reproduction development and diterpenoid biosynthesis[J]. Plant Cell Reports,34(9):1663-1680.
Kaufman K,Melzer R,Theiβen G. 2005. MIKC-type MADS-domain proteins:Structural modularity,protein interactions and network evolution in land plants[J]. Gene,347 (2):183-198.
Kong Z D,Shen Y X,Guo B,Dong X J,Li H Y,Liu P Y. 2015. Cloning and expression of an APETALA1-like gene from Nelumbo nucifera[J]. Genetics and Molecular Research,14(2):6819-6829.
Lamb R S,Irish V F. 2003. Functional divergence within the APETALA3/PISTILLATA floral homeotic gene lineages[J]. Proceedings of the National Academy of Sciences of the United States of America,100(11):6558-6563.
Litt A,Irish V F. 2003. Duplication and diversification in the APETALA1/FRUITFULL floral homeotic gene lineage:Implications for the evolution of floral development[J]. Genetics,165(2):821-833.
Liu Y X,Kong J,Li T Z,Wang Y,Wang A D,Han Z H. 2013. Isolation and characterization of an APETALA1-like gene from pear(Pyrus pyrifolia)[J]. Plant Molecular Biology Reporter,31(4):1031-1039.
Shan H Y,Zhang N,Liu C J,Xu G X,Zhang J,Chen Z D,Kong H Z. 2007. Patterns of gene duplication and functional diversification during the evolution of the AP1/SQUA subfamily of plant MADS-box genes[J]. Molecular Phylogenetics and Evolution,44(1):26-41.
Shchennikova V A,Shugla A O,Immink R,Skryabin G K,Angenent C G. 2004. Identification and characterization of four Chrysanthemum MADS-box genes,belonging to the APETALA1/FRUITFULL and SEPALLATA3 subfamilies[J]. Plant Physiology,134(4):1632-1641.
Shchennikova V A,Shugla A O,Sizeneva S E,Perkovskaya I N,Skryabin K G A. 2011. Diversification of functional activity of the chrysanthemum homeotic MADS-box gene CDM37[J]. Doklady Biochemistry and Biophysics,436(4):29-31.
Shulga A O,Mitiouchkina Y T,Shchennikova V A,Skryabin G K,Dolgov V S. 2011. Overexpression of AP1-like genes from Asteraceae induces early-flowering in transgenic Chrysanthemum plants[J]. In Vitro Cellular and Developmental Biology-Plant,47(5):553-560.
Wang Y,Ma Y P,Fu J X, Qi S,Ma H Z,Dai S L. 2013. Isolation and functional analysis of the ClM8-FRUITFULL-like MADS-box gene from Chrysanthemum lavandulifolium[J]. Scientia Horticulturae,161(2):125-133.
Yang Y Z,Jack T. 2004. Defining subdomains of the K domain important for protein-protein interactions of plant MADS proteins[J]. Plant Molecular Biology,55(1):45-59.
(責任編輯 思利華)
