食物濃度和溫度對萼花臂尾輪蟲種群動態(tài)的影響
2017-05-17 12:24:36鄧平張生元艾桃山喻運珍
江蘇農(nóng)業(yè)科學(xué) 2016年1期
鄧平 張生元 艾桃山 喻運珍


摘要:以濃度分別為2.5×106、5×106、10×106、20×106cells/mL的蛋白核小球藻為食物,在溫度分別為25、28、30℃條件下對萼花臂尾輪蟲進行群體累積培養(yǎng)研究。結(jié)果表明,食物濃度對輪蟲的種群動態(tài)、最大種群密度、種群增長率及非混交卵的日產(chǎn)生量動態(tài)、產(chǎn)量均有明顯影響;溫度對輪蟲的種群動態(tài)、最大種群密度及非混交卵的產(chǎn)量、形成效率、最大形成效率均有顯著影響。食物濃度為20×106cells/mL的小球藻投喂輪蟲,在25~30 qC均能達到較高的種群密度,但25℃培養(yǎng)的輪蟲較28℃和30℃推遲到達峰值。非混交卵的日產(chǎn)生量與種群密度有相似的變化趨勢,28℃和30℃培養(yǎng)的輪蟲非混交卵較25℃提前到達峰值。當輪蟲的種群密度還處于增長階段,非混交卵日生產(chǎn)量已達峰值。因此在輪蟲的培養(yǎng)過程中可考慮監(jiān)測非混交卵的動態(tài)變化預(yù)測輪蟲密度達到峰值的時間。
關(guān)鍵詞:萼花臂尾輪蟲;種群;非混交卵;食物濃度;溫度
中圖分類號:S955.3+2 文獻標志碼:A 文章編號:1002—1302(2016)01—0251—03
食物濃度和溫度是影響輪蟲種群動態(tài)的2個重要生態(tài)因子。有關(guān)食物濃度和溫度對輪蟲種群動態(tài)的影響研究已有較多報道,內(nèi)容涉及溫度和食物濃度對輪蟲種群增長率、最大種群密度、懷卵率、休眠卵產(chǎn)量及形成效率等方面的影響。研究表明溫度和食物濃度對輪蟲的種群增長及休眠卵形成均有明顯的影響,但不同種類、同種種類的不同品系得出的種群增長最適溫度和最適食物濃度各不相同。迄今為止,關(guān)于食物濃度和溫度對輪蟲非混交卵影響方面的研究未見報道。非混交雌體所產(chǎn)的卵為非混交卵,與輪蟲種群數(shù)量密切相關(guān)。
萼花臂尾輪蟲(Brachionus calyciflorus)是淡水中常見的輪蟲種類,也是淡水輪蟲工廠化培養(yǎng)的首選種類。本研究以萼花臂尾輪蟲為對象,研究了適宜溫度范圍內(nèi)不同食物濃度對輪蟲種群及非混交卵的影響,以豐富此方面的內(nèi)容,同時為該輪蟲的規(guī)模化培養(yǎng)提供理論參考。
1材料與方法
1.1輪蟲來源及餌料
萼花臂尾輪蟲于2012年6月采自湖北省武漢市水產(chǎn)科學(xué)研究所旁的水庫,所采水樣首先用多層紗布濾去其中的枝角類和橈足類等大型浮游動物及雜質(zhì),然后置于解剖鏡下觀察,用微吸管吸出其中的萼花臂尾輪蟲并接種到事先準備好的輪蟲培養(yǎng)液中進行培養(yǎng)。試驗時先用紗布過濾,去除在培養(yǎng)輪蟲的過程中產(chǎn)生的廢棄物和剩余餌料的絮凝,再用300目的篩絹濾取輪蟲,并用蒸餾水沖洗輪蟲數(shù)次。
選用蛋白核小球藻(Chlorella pyrenoidsa)作為培養(yǎng)輪蟲的餌料。藻種購自中國科學(xué)院水生生物研究所。小球藻采用sE培養(yǎng)液在光暗周期12 h-12 h實驗室條件下培養(yǎng),光照度2 000~3 000 k,培養(yǎng)溫度25℃,充氣培養(yǎng)。每7~10 d轉(zhuǎn)接1次。投喂輪蟲的藻液經(jīng)300目篩絹過濾,以去除藻液中的絮凝。
1.3計算方法
輪蟲及其非混交卵的計數(shù)方法、有關(guān)參數(shù)的定義和計算方法均與文獻[7]相同。
1.4統(tǒng)計分析
不同培養(yǎng)條件下的輪蟲種群數(shù)量及非混交卵日產(chǎn)生量的動態(tài)變化采用重復(fù)測量的方差分析(repeated measure ANO-VA)。對不同食物濃度、不同溫度條件下的輪蟲種群最大密度、種群增長率的差異顯著性采用兩因素方差分析進行分析。對非混交卵的產(chǎn)量、形成效率及最大形成效率的差異顯著性也采用兩因素方差分析(two-way ANOVA)進行分析(SPSS19.0.P<0.05)。
2結(jié)果與分析
2.1食物濃度和溫度對輪蟲種群增長的影響
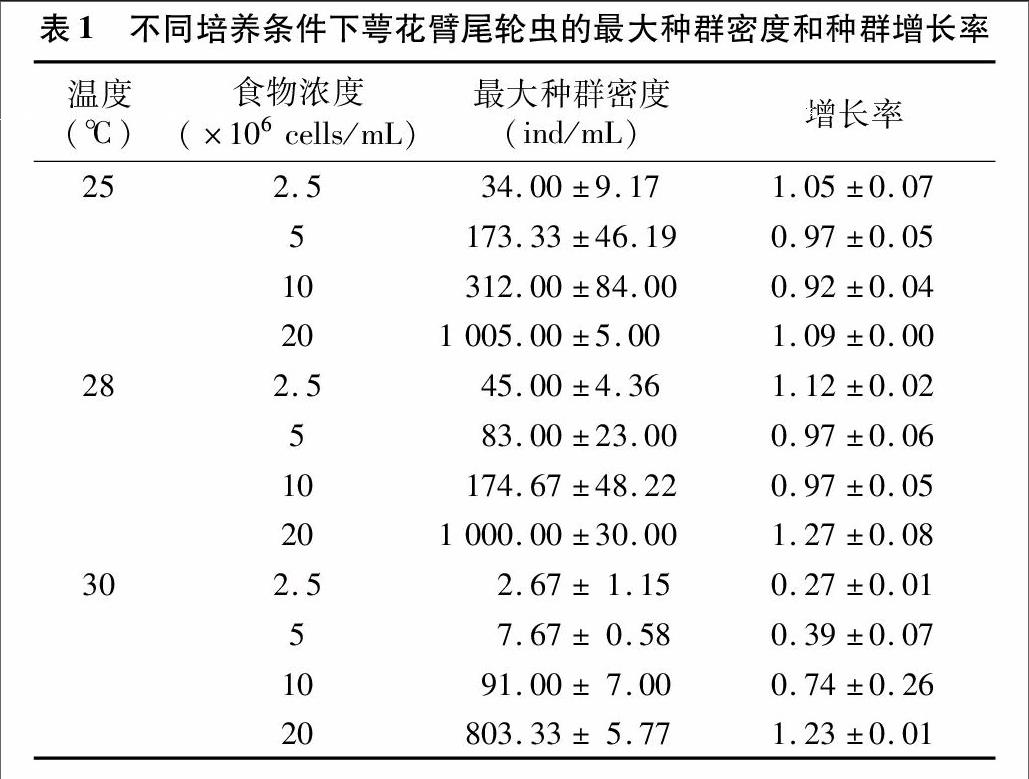
2.1.1食物濃度和溫度對輪蟲種群數(shù)量的影響 在各食物濃度及溫度條件下,輪蟲種群的動態(tài)變化如圖1所示。重復(fù)測量的方差分析結(jié)果表明食物濃度和溫度對輪蟲種群動態(tài)有顯著作用(P<0.05)。隨著培養(yǎng)溫度的升高,輪蟲繁殖高峰值出現(xiàn)的時間也總體上呈現(xiàn)逐漸前移的趨勢。溫度為28、30℃,食物濃度為20×106 cells/mL,培養(yǎng)4~6 d輪蟲種群的密度培養(yǎng)增長極顯著(P<0.01),然后又迅速下降。25℃時,20×106 cells/mL食物濃度下,輪蟲種群密度從培養(yǎng)5 d開始顯著增長(P<0.05)。
2.1.2食物濃度和溫度對輪蟲最大種群密度和種群增長率的影響各條件組合下,輪蟲的最大密度和種群增長率見表1。兩因素方差分析結(jié)果表明,溫度、食物濃度以及兩者之間的交互作用均對最大種群密度具有極顯著影響(P<0.01)。種群增長率的兩因素方差分析結(jié)果顯示,溫度30℃與其他2個溫度之間的種群增長率差異極顯著(P<0.01),而25℃和28℃條件下種群增長率無明顯差異(P>0.05);食物濃度為20×106 cells/mL時輪蟲的種群增長率遠高于其他3種食物濃度(P<0.01);溫度和食物濃度的交互作用對輪蟲的種群增長率具有極顯著影響(P<0.01)。
2.2食物濃度和溫度對輪蟲非混交卵的影響
2.2.1食物濃度和溫度對輪蟲非混交卵日產(chǎn)生量的影響 各食物濃度及溫度條件下,輪蟲非混交卵日產(chǎn)生量的動態(tài)變化如圖2所示。重復(fù)測量的方差分析結(jié)果表明,食物濃度為20×106 cells/mL的非混交卵形成動態(tài)與其他各組差異極顯著(P<0.01),30℃條件下除外。25℃食物濃度為2.5×106 cells/mL輪蟲非混交卵增長不明顯,其他3個食物濃度培養(yǎng)4 d非混交卵日產(chǎn)生量開始增加,20×106cells/mL食物濃度的非混交卵日產(chǎn)生量在培養(yǎng)6 d時達到峰值105 ind/mL。28℃條件下各食物濃度均在培養(yǎng)3 d非混交卵日產(chǎn)生量開始增加,20×106 cells/mL食物濃度的非混交卵日產(chǎn)生量在培養(yǎng)4 d達到峰值131 ind/mL。30℃條件下,20×106 cells/mL食物濃度的非混交卵日產(chǎn)生量在培養(yǎng)4 d顯著增長,培養(yǎng)5 d達到峰值90 ind/mL,其他3個食物濃度非混交卵日產(chǎn)生量增長不明顯。
2.2.2食物濃度和溫度對輪蟲非混交卵產(chǎn)量和形成效率的影響 在輪蟲非混交卵的第1個形成周期內(nèi),各條件組合下非混交卵的產(chǎn)量、形成效率和最大形成效率見表2。兩因素方差分析結(jié)果顯示,在培養(yǎng)全過程中,溫度、食物濃度以及兩者之間的交互作用均對非混交卵產(chǎn)量具有極顯著影響(P<0.01)。對非混交卵形成效率進行的兩因素方差分析結(jié)果表明,溫度對非混交卵的形成效率具有極顯著影響(P<0.01);食物濃度對非混交卵形成效率沒有明顯作用(P>0.05);溫度與食物濃度的交互作用對非混交卵的形成效率具有極顯著影響(P<0.01)。對最大形成效率進行的兩因素方差分析結(jié)果顯示,溫度對非混交卵的最大形成效率具有極顯著影響(P<0.01);食物濃度為2.5×106cells/mL的最大形成效率與其他各組差異極顯著(P<0.01);溫度與食物濃度的交互作用對非混交卵的最大形成效率具有極顯著影響(P<0.01)。
3討論
3.1食物濃度和溫度對輪蟲種群增長的影響
已有研究結(jié)果表明,同一溫度下萼花臂尾輪蟲種群密度隨食物濃度的升高而增加。本研究結(jié)果與已有的研究結(jié)果一致,25、28、30℃條件下萼花臂尾輪蟲的種群動態(tài)與食濃度密切相關(guān)。在同一食物濃度下,種群密度隨溫度的升高而增大。本研究中25℃和28℃條件下各食物濃度條件下的種群密度有相同的變化趨勢,但30℃條件下各食物濃度沒有表現(xiàn)出這種變化趨勢,可能與較高溫度條件對低起始物密度的輪蟲種群增長有抑制作用有關(guān)。
黃祥飛研究發(fā)現(xiàn),萼花臂尾輪蟲種群的最大密度隨溫度的升高而有所增加,而種群達到高峰所需的時間則隨著溫度的升高而縮短。本研究結(jié)果也表明,輪蟲種群在28℃和30℃條件下較25℃提前1 d達到最大密度,但25℃與28℃條件下的輪蟲種群增長率差異不顯著,且明顯高于30℃。
3.2食物濃度和溫度對輪蟲非混交卵的影響
已有關(guān)于輪蟲繁殖方面的研究主要是通過考察輪蟲的懷卵量這一指標,懷卵輪蟲既有產(chǎn)非混交卵的非混交雌體又有產(chǎn)休眠卵的混交雌體。非混交雌體與輪蟲種群增長密切相關(guān),考察非混交雌體所產(chǎn)非混交卵的動態(tài)變化將能更準確地預(yù)測出輪蟲種群密度的變化。本研究中同一溫度下萼花臂尾輪蟲種群密度隨食物濃度的升高而增加,非混交卵的日產(chǎn)生量也表現(xiàn)出相似的變化趨勢。同一食物濃度下,非混交卵動態(tài)與種群數(shù)量動態(tài)也表現(xiàn)出極大的相似性,即25℃和28℃條件下各食物濃度條件下的非混交卵日產(chǎn)生量與種群密度有相同的變化趨勢,30℃條件下非混交卵日產(chǎn)生量沒有表現(xiàn)出這種變化趨勢。
溫度為25℃時輪蟲的非混交卵產(chǎn)量、形成效率和最大形成效率均明顯高于28℃和30℃條件下,這可能與25℃條件下輪蟲非混交卵的日產(chǎn)生量增長時間較長有關(guān)。研究還發(fā)現(xiàn)當輪蟲的種群密度還處于增長階段,非混交卵日生產(chǎn)量已達峰值。因此可用非混交卵的動態(tài)變化提前預(yù)測出輪蟲種群的動態(tài)變化。
