多巴胺轉運體基因敲除小鼠認知功能的變化
2017-05-10 12:41:13黃國明鈔雪林陳建云
中國老年學雜志 2017年8期
黃國明 鈔雪林 陳建云
(南昌大學第一附屬醫院精神科,江西 南昌 330006)
多巴胺轉運體基因敲除小鼠認知功能的變化
黃國明 鈔雪林 陳建云
(南昌大學第一附屬醫院精神科,江西 南昌 330006)
目的 觀察多巴胺轉運體(DAT)基因敲除小鼠認知功能的變化。方法 采用Western印跡及PCR方法檢測DAT基因敲除C57BL/6 J小鼠模型,通過對DAT基因敲除小鼠進行Morris 水迷宮試驗及曠場試驗研究小鼠認知能力的變化。結果 在Morris 水迷宮試驗中,與WT組小鼠相比,DAT基因敲除小鼠平均逃避潛伏期明顯延長,顯示其學習能力下降;在撤掉平臺后,其在原平臺象限停留時間縮短,表明其記憶能力亦下降。在曠場試驗中,與WT組小鼠相比,DAT基因敲除小鼠總的探索路程降低,雖然其自主興奮性運動增加,但多為無目的性運動,其在中央格停留時間及次數也下降,表明其探索好奇能力明顯下降。結論 敲除小鼠DAT基因可導致小鼠多動,同時損傷小鼠的認知功能,表明DAT基因可能和學習、記憶、探索能力有關。
多巴胺轉運體;認知功能;Morris 水迷宮試驗;曠場試驗
多巴胺(DA)在行為、認知、情感、獎賞等多種中樞神經系統功能活動中具有重要作用。DA能系統異常參與導致精神分裂癥、藥物成癮、帕金森疾病、遲發性運動障礙、路易體癡呆、阿爾茨海默病、注意力缺陷多動癥等多種神經精神疾病。DA能神經遞質信號的傳遞受突觸間隙DA濃度的影響,即同時受DA能神經元合成釋放DA到突觸間隙的能力及DA轉運體(DAT)重攝取突觸間隙內DA到神經末梢的能力調控〔1,2〕。本研究采用DAT基因敲除小鼠模型(DAT-KO),探討DAT基因敲除后小鼠認知功能的變化。
1 材料和方法
1.1 材料 純合子DAT基因敲除C57BL/6 J小鼠(基因型DAT-/-)為HO組,DAT基因敲除C57BL/6 J小鼠(基因型DAT+/-)為HET組,野生型C57BL/6 J小鼠(基因型DAT+/+)為WT組,每組13只,20月齡,來源于上海軟隆科技發展有限公司。WT組小鼠均重40~43 g,皮毛光滑,飲食、 活動、體質量增長正常;HET組小鼠均重35~40 g,皮毛光滑,飲食、 活動、體質量增長正常;HO組小鼠均重32~36 g,皮毛不整,飲食正常,多動。
1.2 方法 各組取3只小鼠斷頭處死,取大腦組織。(1)PCR驗證基因型:以100 mg組織加入1 ml Trizol勻漿提取總RNA。按照TAKA逆轉錄試劑盒的要求進行逆轉錄反應得到cDNA,-80℃冷凍保存。引物設計及合成:根據GenBank所給出的基因序列,采用Primer 5.0軟件設計各基因的引物,GAPDH上游引物5'-GAGACCTTCAACACCCCAGCG-3',下游引物5'-TCGGGGCATCGGAACCGCTCA-3';DAT上游引物5'-GGCGGGAGTGGTATTATGAA-3',下游引物5'-GTGATCCGGAAGGACTGATT-3'。引物由上海生工生物工程技術服務有限公司合成。real-time PCR反應:以cDNA為模板,在7500 real-time PCR反應體系中進行定量PCR反應擴增,采用2-ΔΔCT法計算各組的RNA表達差別。
(2)Western印跡驗證基因型:以1 g組織加入RIPA裂解液,冰上裂解30 min。4℃,10 000 r/min離心15 min,取上清液,用BCA法測定蛋白濃度為1 μg/μl,分裝后-80℃冷凍保存。取相同質量的裂解緩沖液上樣進行SDS-PAGE膠電泳,分離后轉移至PVDF膜,5%牛奶封閉1 h,分別用抗GAPDH抗體(1∶1 000;Abcam公司)及抗DAT抗體(1∶1 000;Abcam公司)孵育4℃過夜,用TBST溶液洗膜后,加入1∶5 000稀釋二抗室溫孵育1 h,洗膜后ECL顯色掃描。應用Image J分析軟件進行灰度掃描分析,實驗重復3次。
1.2.1 Morris水迷宮試驗 水迷宮裝置為一圓形水池,直徑150 cm,高50 cm,水深28 cm,水溫(25±1)℃,可升降平臺直徑8 cm,高25 cm。水迷宮分為 4個象限,水池上方由攝像頭與計算機相連,記錄小鼠游泳軌跡,水迷宮測試時環境應保證黑暗和安靜,避免小鼠的行為受外界干擾。
(1)定位航行試驗(反映空間記憶的獲得能力):在水池內某一位置固定平臺,將小鼠面向池壁放入水中,記錄其在 2 min 內尋找到并爬上平臺的時間(逃避潛伏期)和路程,小鼠找到并爬上平臺后,讓其停留5 s,若小鼠入水后2 min內未能找到平臺,則將其引導置于平臺上,并停留10 s,此時記錄逃避潛伏期為 120 s。每日于固定的時間訓練2次(上午9:00,下午2:00),連續5 d。每次訓練完成后迅速將小鼠擦干。每日各小鼠逃避潛伏期用2次訓練的均值表示。
(2)空間探索實驗(檢測空間記憶的保持能力):在第6天(即為期5 d的定位航行實驗結束后)去除平臺,將小鼠從固定象限中點面朝池壁放入水中,記錄小鼠2 min池內活動情況,通過分析獲得2 min內小鼠經過原平臺所在靶象限的時間。
1.2.2 曠場試驗 曠場實驗是評價實驗動物在新環境中自主行為、探究行為與恐懼焦慮感的一種常用方法。小鼠由于對新開闊環境恐懼而主要在周邊區域活動,但其探究特性又促使其產生在中央區域活動的動機,小鼠在中央格停留時間及次數代表其探索好奇的程度。實驗采用長寬高均為50 cm的無頂塑料箱,內壁為黑色,底部分為25個等份小方格。將小鼠從中央格放入箱內,由攝像系統記錄小鼠5 min內的探索活動,追蹤其探索路程及在中央格停留時間及次數。每只老鼠適應結束后用酒精擦拭箱子,避免前只老鼠氣味干擾。
1.3 統計學分析 采用SPSS17.0軟件進行單因素方差分析和Tukey法。
2 結 果
2.1 Western印跡及PCR驗證小鼠基因型 DAT mRNA在HO組基本不表達(0.09±0.02),HET組(0.65±0.14)較WT組(1.19±0.78)下降50%,差異有統計學意義(P<0.01)。進一步采用Western印跡對小鼠腦組織蛋白進行檢測發現,DAT蛋白在HO組也基本不表達(0.07±0.01),HET組DAT蛋白水平(0.34±0.11)則較WT組(0.79±0.24)下降70%,差異有統計學意義(P<0.01),見圖1。表明建立DAT基因敲除小鼠模型成功。
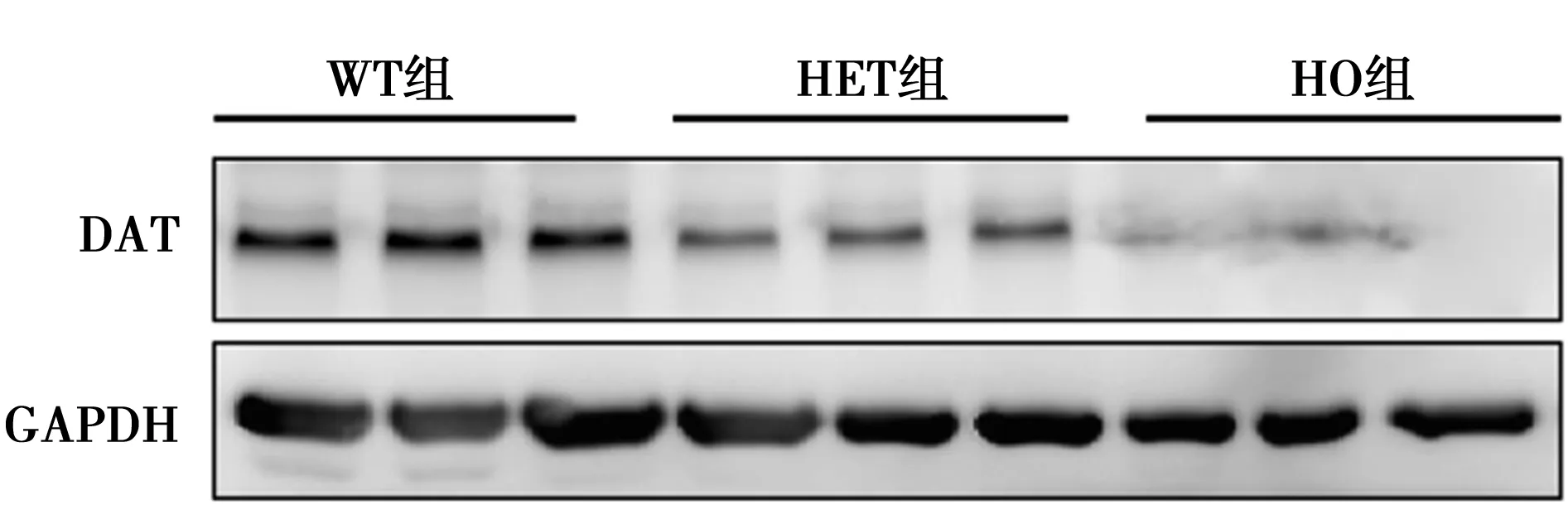
圖1 各組大鼠DAT蛋白相對表達水平
2.2 Morris水迷宮試驗結果
2.2.1 DAT基因敲除對小鼠學習能力的影響 各組小鼠的平均逃避潛伏期均逐漸縮短(圖2)。經過5 d的Morris水迷宮定位航行實驗訓練,WT組、HET組、HO組第5天逃避潛伏期均值分別為(36.32±8.76)s,(68.12 ± 8.15)s,(90.03 ±11.81)s,均短于第1天的(53.39 ±10.45)s,(85.97 ± 8.10)s,(110.20 ±2.32) s(P<0.01)。表明隨著定位航行訓練次數的增加,各組小鼠都具有了一定的空間學習能力。同時HET組及HO組平均逃避潛伏期均明顯高于WT組,表明其空間學習能力較WT組下降。
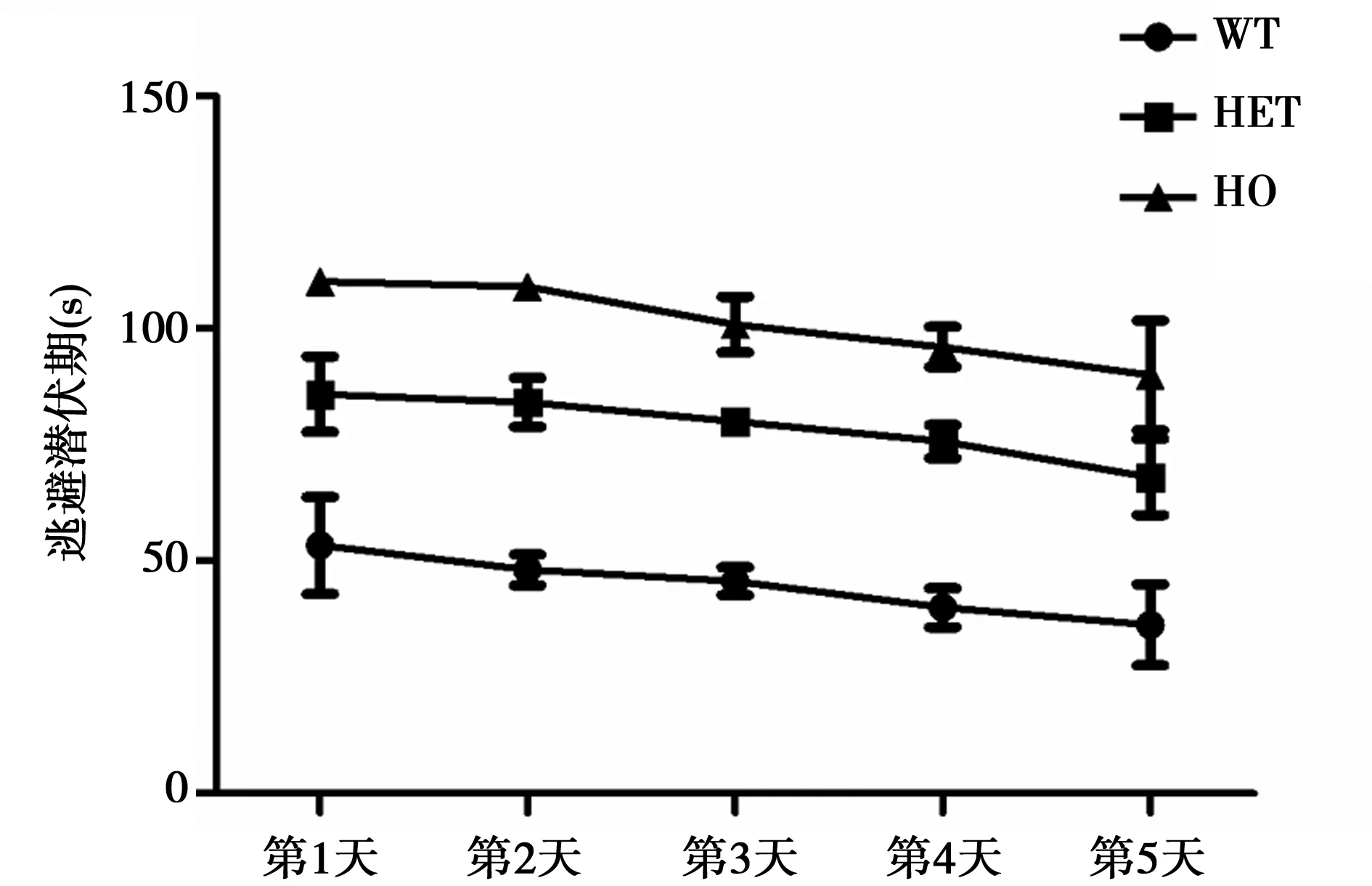
圖2 各組大鼠定位航行試驗平均逃避潛伏期變化
2.2.2 DAT基因敲除對小鼠記憶能力的影響 在訓練小鼠完成5 d的 Morris水迷宮定位航行實驗后,于第6天進行空間探索試驗,與WT組〔43.14±7.23)s〕比較,HET組〔(23.31±4.28)s〕在原平臺所在象限的活動時間稍有延長,HO組〔14.21±5.31)s〕明顯短于WT組(P<0.05,P<0.01),表明基因敲除可損傷小鼠的記憶能力。
2.3 曠場試驗結果
2.3.1 各組小鼠探索路程 WT組探索路程(1 708.30±58.95)cm,HET組(1 671.40±59.96)cm,HO組(909.70±38.15)cm,HET組與WT組比較差異無統計學意義(P>0.05),而HO組小鼠明顯低于WT組(P<0.01)。
2.3.2 各組小鼠中央格停留時間及次數 與WT組〔(43.15±9.78)s、(10.12±2.33)次〕比較,HET組〔(21.15±7.08)s、(5.42±1.33)次〕中央格停留時間及次數均低于WT組(P<0.05);HO組〔(10.04±3.28)s、(2.12±1.01)次〕明顯低于WT組(P<0.01),表明基因敲除可降低小鼠探索好奇能力。
3 討 論
DA是重要的神經遞質,在腦內發揮極其重要的功能〔3〕,DA參與的神經通路有黑質-紋狀體通路(調節軀體自主運動),該通路受損可導致帕金森病及其他運動障礙性疾病〔4〕;以及中腦-皮層傳導束和中腦-邊緣傳導束(參與情緒、行為動機和學習記憶等心理和認知功能的調節)障礙〔5〕,該通路異常則與精神分裂、藥物成癮相關〔6〕。DAT位于DA能神經元突觸周圍的細胞膜上,是一種Na+/Cl-依賴轉運體〔7〕,它的主要功能是重攝取突觸間隙內的DA,從而調控突觸間隙內DA的有效濃度,維持DA循環,減少對新合成DA的需求,保持DA在神經元中的穩態〔8〕。敲除 DAT基因破壞突觸間隙內DA的重攝取及清除過程,Isingrini等〔9,10〕研究結果顯示敲除DAT基因導致突觸間隙內DA存在時長增加100倍;Gainetdinov等〔11,12〕研究也表明敲除DAT基因導致突觸間隙內DA水平至少升高 5 倍,存在時間延長 300 倍。因此,敲除DAT基因可能會對小鼠的運動、認知、情緒等各方面產生重要影響。
本研究通過對DAT基因敲除小鼠進行Morris 水迷宮試驗及曠場試驗研究小鼠認知能力的變化,結果顯示,敲除DAT基因可能導致小鼠學習、記憶、探索能力均下降,導致小鼠認知減退。
研究發現DAT基因敲除小鼠的行為學表型研究發現,完全敲除DAT的 DAT-KO小鼠表現為自發性興奮性運動明顯增加,同時其曠場試驗及水迷宮試驗能力下降〔13〕,但Spielewoy等〔14〕認為曠場試驗及水迷宮試驗能力下降的原因為小鼠的自發性運動導致其無法進行學習、探索試驗,未明確提出其認知能力下降。Del′Guidice等〔15〕采用H型迷宮試驗檢測DAT基因敲除小鼠的行為學表現,認為小鼠多動不干擾其認知能力,敲除小鼠DAT基因導致小鼠認知能力下降,與本研究結果一致。利用PET檢測人大腦發現DAT的表達量下降與大腦老化相關,DAT的下降與路易體癡呆相關,同時DAT SPECT技術正被探索用于區分帕金森病、路易體癡呆及阿爾茨海默病〔16〕,這可能與DAT缺失導致突觸間隙DA量升高有關,Morice等〔17〕發現給予DAT-KO小鼠DA競爭性抑制藥物可以改善認知能力下降,認為DAT基因敲除導致的突觸間隙高DA水平參與導致認知能力下降。目前認為DA、五羥色胺、煙堿等均與認知功能相關,有研究表明給予DAT-KO小鼠尼古丁可以改善其認知能力下降〔18,19〕。運動和認知能力的維持可能有賴于腦內多種遞質間的平衡〔20〕,包括DA在內的大腦遞質系統與認知功能之前的關系尚需更多機制研究。
本研究結果表明敲除小鼠DAT基因不僅導致小鼠自主性興奮性運動增加,而且導致小鼠認知能力降低,DAT基因參與學習、記憶、探索能力的調控,為進一步研究相關的神經疾病提供了理論基礎。
1 Weinstein JJ,Chohan MO,Slifstein M,etal.Pathway-specific dopamine abnormalities in schizophrenia 〔J〕.Biol Psychiatry,2017;81(1):31-42.
2 Ayers-Ringler JR,Jia YF,Qiu YY,etal.Role of astrocytic glutamate transporter in alcohol use disorder 〔J〕.World J Psychiatry,2016;6(1):31-42.
3 Arnsten AF,Wang M,Paspalas CD.Dopamine's actions in primate prefrontal cortex:challenges for treating cognitive disorders 〔J〕.Pharmacol Rev,2015;67(3):681-96.
4 Yulug B,Hanoglu L,Kilic E.The neuroprotective role of vesicular monoamine transporter 2 in neurodegenerative diseases 〔J〕.Med Chem,2015;11(2):104-8.
5 Arnsten AF,Girgis RR,Gray DL,etal.Novel dopamine therapeutics for cognitive deficits in schizophrenia 〔J〕.Biol Psychiatry,2017;81(1):67-77.
6 dela Pena I,Gevorkiana R,Shi WX.Psychostimulants affect dopamine transmission through both dopamine transporter-dependent and independent mechanisms 〔J〕.Eur J Pharmacol,2015;764:562-70.
7 Brooks DJ.Molecular imaging of dopamine transporters 〔J〕.Ageing Res Rev,2016;34:114-21.
8 Raiteri L,Raiteri M.Multiple functions of neuronal plasma membrane neurotransmitter transporters 〔J〕.Prog Neurobiol,2015;134:1-16.
9 Isingrini E,Perret L,Rainer Q,etal.Resilience to chronic stress is mediated by noradrenergic regulation of dopamine neurons 〔J〕.Nat Neurosci,2016;19(4):560-3.
10 Isingrini E,Perret L,Rainer Q,etal.Selective genetic disruption of dopaminergic,serotonergic and noradrenergic neurotransmission:insights into motor,emotional and addictive behaviour 〔J〕.J Psychiatry Neurosci,2016;41(3):169-81.
11 Gainetdinov RR.Dopamine transporter mutant mice in experimental neuropharmacology 〔J〕.Naunyn Schmiedebergs Arch Pharmacol,2008;377(4-6):301-13.
12 Homberg JR,Kyzar EJ,Nguyen M,etal.Understanding autism and other neurodevelopmental disorders through experimental translational neurobehavioral models 〔J〕.Neurosci Biobehav Rev,2016;65:292-312.
13 Reith ME,Blough BE,Hong WC,etal.Behavioral,biological,and chemical perspectives on atypical agents targeting the dopamine transporter 〔J〕.Drug Alcohol Depend,2015;147:1-19.
14 Spielewoy C,Gonon F,Roubert C,etal.Increased rewarding properties of morphine in dopamine-transporter knockout mice 〔J〕.Eur J Neurosci,2000;12(5):1827-37.
15 Del'Guidice T,Lemasson M,Etievant A,etal.Dissociations between cognitive and motor effects of psychostimulants and atomoxetine in hyperactive DAT-KO mice 〔J〕.Psychopharmacology (Berl),2014;231(1):109-22.
16 Shingai Y,Tateno A,Arakawa R,etal.Age-related decline in dopamine transporter in human brain using PET with a new radioligand 〔(1)(8)F〕FE-PE2I 〔J〕.Ann Nucl Med,2014;28(3):220-6.
17 Morice E,Billard JM,Denis C,etal.Parallel loss of hippocampal LTD and cognitive flexibility in a genetic model of hyperdopaminergia 〔J〕.Neuropsychopharmacology,2007;32(10):2108-16.
18 McCleery J,Morgan S,Bradley KM,etal.Dopamine transporter imaging for the diagnosis of dementia with Lewy bodies 〔J〕.Cochrane Database Syst Rev,2015;1:CD010633.
19 Weiss S,Nosten-Bertrand M,McIntosh JM,etal.Nicotine improves cognitive deficits of dopamine transporter knockout mice without long-term tolerance 〔J〕.Neuropsychopharmacology,2007;32(12):2465-78.
20 Salamone JD,Correa M,Yohn S,etal.The pharmacology of effort-related choice behavior:dopamine,depression,and individual differences 〔J〕.Behav Process,2016;127:3-17.
〔2016-12-26修回〕
(編輯 滕欣航)
黃國明(1979-),男,碩士,副主任醫師,主要從事老年精神障礙研究。
R395.4
A
1005-9202(2017)08-1866-04;
10.3969/j.issn.1005-9202.2017.08.019
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
發明與創新(2022年30期)2022-10-03 08:40:56
動漫星空(興趣百科)(2020年12期)2020-12-12 05:31:40
學苑創造·A版(2020年9期)2020-10-13 09:41:02
人大建設(2018年6期)2018-08-16 07:23:10
文理導航·科普童話(2017年5期)2018-02-10 19:42:14
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
無人機(2017年10期)2017-07-06 03:04:36
小星星·閱讀100分(低年級)(2015年10期)2015-10-22 08:30:04
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
