生長調節劑對低溫脅迫下酸橙幼苗抗逆生理指標的影響*
2017-04-27 06:44:20梁永富王康才陳興忠
林業科學 2017年3期
梁永富 王康才 薛 啟 隋 利 葉 軍 陳興忠
(1.南京農業大學園藝學院 南京 210095; 2.泰州市林業技術指導站 泰州 225300)
?
生長調節劑對低溫脅迫下酸橙幼苗抗逆生理指標的影響*
梁永富1王康才1薛 啟1隋 利1葉 軍2陳興忠2
(1.南京農業大學園藝學院 南京 210095; 2.泰州市林業技術指導站 泰州 225300)
【目的】分析不同濃度BRs、BT和SA對低溫脅迫下酸橙幼苗抗逆生理指標的影響,尋找緩解酸橙幼苗低溫傷害的可行方法,為酸橙的推廣種植提供技術參考。【方法】以盆栽酸橙幼苗為材料,葉面分別噴施不同濃度油菜素內酯(BRs)、甜菜堿(BT)和水楊酸(SA)溶液后,置于4 ℃/0 ℃(晝/夜)、光照600 μmol·m-2s-1的人工氣候箱內,進行低溫脅迫處理。以常溫(溫度25 ℃/23 ℃(晝/夜),光照600 μmol·m-2s-1)下噴施蒸餾水為CK1,以低溫脅迫(4 ℃/0 ℃(晝/夜),光照600 μmol·m-2s-1)下噴施蒸餾水為CK2。低溫脅迫處理3天后,測定酸橙幼苗葉片細胞膜透性、丙二醛、脯氨酸、可溶性蛋白、葉綠素含量,以及SOD、POD、CAT抗氧化酶活性。【結果】與CK1相比,低溫顯著抑制CK2酸橙幼苗葉片抗氧化酶活性,加重低溫對細胞膜的傷害,致使細胞膜結構被破壞,不能正常行使其功能,進而引起酸橙幼苗葉片相對電導率和丙二醛含量顯著上升。低溫處理也會導致葉綠體結構被破壞,引起光合色素的分解,致使葉綠素和類胡蘿卜素含量顯著下降,不能正常進行光合作用。與CK2相比,葉面噴施生長調節劑可以顯著提高酸橙幼苗葉片脯氨酸和可溶性蛋白含量,減輕酸橙幼苗葉片因低溫而導致的細胞失水; 提高SOD、POD、CAT抗氧化酶活性,有效清除細胞內過多的活性氧,避免膜脂過氧化的發生,顯著降低葉片丙二醛含量和相對電導率,使細胞膜系統代謝正常,減輕低溫脅迫對酸橙幼苗葉片造成的傷害。同時,葉面噴施生長調節劑顯著提高葉片葉綠素與類胡蘿卜素含量,減弱低溫對光合作用的抑制影響。隨著濃度的升高,生長調節劑對酸橙幼苗低溫傷害的緩解作用呈現先增強后減弱的趨勢,其中以葉面噴施0.15 mg·L-1BRs、10 mmol·L-1BT、1 mmol·L-1SA溶液的效果最為明顯。【結論】葉面噴施濃度為0.15 mg·L-1BRs、10 mmol·L-1BT、1 mmol·L-1SA溶液可明顯減輕低溫脅迫對酸橙幼苗葉片的傷害。關鍵詞: 酸橙幼苗; 低溫脅迫; 外源物質; 抗逆
蕓香科植物酸橙(Citrusaurantium)是一種經濟及觀賞價值較高的果樹,也是中藥材枳殼的基原植物。枳殼具有理氣寬中、行滯消脹的功效,用于治療胸脅氣滯、脹滿疼痛、食積不化、痰飲內停、臟器下垂,為常用中藥材。酸橙主要分布于江蘇、浙江、廣東、貴州、四川、江西等省。在江蘇泰州大田種植過程中發現,冬春季持續低溫會導致酸橙葉片逐漸枯黃掉落,此時酸橙已開始進入春芽和開花期,持續低溫給酸橙的花芽分化及坐果造成一定的傷害,嚴重影響枳殼產量。因此,如何緩解低溫傷害,對酸橙的推廣種植和提高枳殼產量具有重要意義。
目前研究表明,葉片表面噴施生長調節劑可顯著提高植物抗寒性,緩解植物的低溫傷害。低溫脅迫下,植物激素油菜素內酯(BRs)通過增加電子流向卡爾文循環的轉移,減少活性氧(ROS)的產生,從而提高農作物的抗逆性(Yuetal., 2002; 2004; Khripachetal., 2000; Krishnaetal., 2003; Ogwenoetal., 2008)。甜菜堿(BT)能夠保護細胞膜和穩定抗氧化酶等生物大分子的結構,增強細胞有效清除ROS的能力,減輕低溫對細胞造成的傷害(郭啟芳等, 2004; 張波等, 2005)。同時,細胞內BT的增加,還可以降低胞內水勢和冰點,減輕植物因失水和結冰而造成的低溫傷害(劉文英, 2015)。水楊酸(SA)是植物體內普遍存在的酚類化合物,能夠導致逆境反應信號轉導分子H2O2含量的上升,從而引發系統獲得性抗性反應的產生(Shirasuetal., 1997; Luoetal., 2001; Kratschetal., 2000),幫助植物度過不良環境(Chenetal., 1993a; 1993b)。相關研究表明,用BRs和SA噴施黃瓜(Cucumissativus)幼苗可以緩解其幼苗光合系統的低溫傷害(Huetal., 2010; 劉偉等, 2009),提高植物的凈光合速率。康國章等(2003)發現SA通過對植物進行類似于冷馴化的鍛煉過程,提高抗氧化酶活性,保護光合器官,從而增強香蕉(Musasapientum)抗逆性。低溫脅迫下,葉面噴施BT可以顯著提高結縷草(Zoysiajaponica)抗氧化酶活性(梁小紅等, 2015),外源BT還能提高煙草(Nicotianatabacum)葉片類囊體膜功能,增強煙草對低溫脅迫的抵抗力(Wangetal., 2008)。雖然BRs、BT、SA在提高植物抗逆性方面的研究已經很多,但對于低溫脅迫下酸橙幼苗抗逆性的影響卻未見報道。本文根據酸橙栽培過程中遇到的實際問題,研究噴施不同濃度BRs、BT和SA對低溫脅迫下酸橙幼苗葉片生理生化特性的影響,以探討利用外源物質提高酸橙幼苗抗寒能力的可行性,以期為酸橙的推廣種植提供技術參考。
1 材料與方法
1.1 試驗材料和處理方法
試驗用酸橙幼苗由湖南程錦農業科技開發有限公司提供,是2015年4月中旬經種子發芽的實生苗,試驗于2015年9—11月在南京農業大學溫室內進行。于2015年9月1日選取生長基本一致、無病蟲害、高10 cm 左右的酸橙幼苗栽種于高16 cm、直徑14 cm的塑膠盆中,栽培基質為蛭石與營養土以2∶1混合,基質體積約為花盆體積的80%,每盆4株。常規水肥管理,每5天澆水1次,每次澆水量為150 mL,緩苗1個月后進行生長調節劑和低溫脅迫處理,此時苗高11.3 cm左右,平均基莖為0.96 cm。
1.2 試驗設計
選用SA、BT、BRs 3種生長調節劑處理酸橙幼苗,SA設0.2,0.5,1.0,1.5,2.0 mmol·L-15個濃度,BT設5,10,15,20,25 mmol·L-15個濃度, BRs設0.05,0.1,0.15,0.2,0.25 mg·L-15個濃度。于2015年10月1—3日18:00對酸橙進行葉面噴施,以葉片正反面浸潤、向下滴液為度,每處理4盆,連續噴施3天。第3天噴施生長調節劑2 h后(康國章等, 2003; 宋金鳳等, 2012),將幼苗連同塑膠盆放入人工氣候箱(QHX-800L人工氣候箱,金壇市良友儀器有限公司)中進行低溫脅迫處理,溫度為4 ℃/0 ℃(晝/夜),光照為600 μmol·m-2s-1,同時設置2組對照。對照CK1為置于溫度25 ℃/23 ℃(晝/夜),噴施蒸餾水; CK2為低溫脅迫,噴施蒸餾水。低溫處理3天后(康國章等, 2003; 宋金鳳等, 2012; Huetal., 2010)取出苗木,在室溫下恢復24 h后,隨機采取4~6片成熟葉(幼苗中部)進行相關生理指標測定。
1.3 指標測定
可溶性蛋白含量測定采用考馬斯亮藍G-250染色法; 脯氨酸含量測定采用茚三酮法; 丙二醛(MDA)含量測定采用硫代巴比妥酸(TBA)法; 超氧化物歧化酶(SOD)和過氧化氫酶(CAT)活性測定分別采用氮藍四唑(NBT)法和高錳酸鉀滴定法; 過氧化物酶(POD)活性測定采用愈創木酚法; 葉綠素含量采用乙醇提取分光光度法; 相對電導率采用電導儀法(李合生, 2000)。
1.4 數據統計與分析
采用Excel 2010和統計軟件SPSS 20.0進行統計分析,LSD法檢驗差異顯著性。
2 結果與分析
2.1 低溫脅迫下生長調節劑對酸橙幼苗葉片細胞膜透性的影響
2.1.1 MDA含量變化 MDA是膜脂過氧化產物,通過測量MDA含量可以了解細胞膜受到ROS氧化傷害的程度。從表1可知,與CK1相比,CK2酸橙幼苗葉片MDA含量顯著增加254.66%。經不同濃度生長調節劑處理后,酸橙幼苗葉片MDA含量較CK2均顯著降低,說明葉面噴施生長調節劑能顯著減少ROS的產生,降低膜脂過氧化的程度,從而有效抵御低溫對酸橙幼苗的傷害。其中0.15 mg·L-1BRs、10 mmol·L-1BT、1 mmol·L-1SA處理效果最為顯著,相對于CK2,分別降低57.03%,60.54%,60.54%。2.1.2 相對電導率的變化 相對電導率是衡量細胞膜透性的重要指標,其值越大,說明細胞膜透性越大,細胞受到傷害的程度就越高。從表1可知,與CK1相比,CK2葉片相對電導率顯著升高124.23%,說明低溫脅迫破壞了酸橙幼苗的細胞膜系統。噴施3種生長調節劑后,與CK2相比,均不同程度降低酸橙幼苗葉片相對電導率,隨著3種生長調節劑噴施濃度的升高相對電導率出現先降低后升高的趨勢。與CK2相比,濃度為0.15 mg·L-1BRs、10 mmol·L-1BT、1 mmol·L-1SA處理的酸橙幼苗葉片相對電導率顯著下降,分別降低41.86%,44.49%,43.07%。
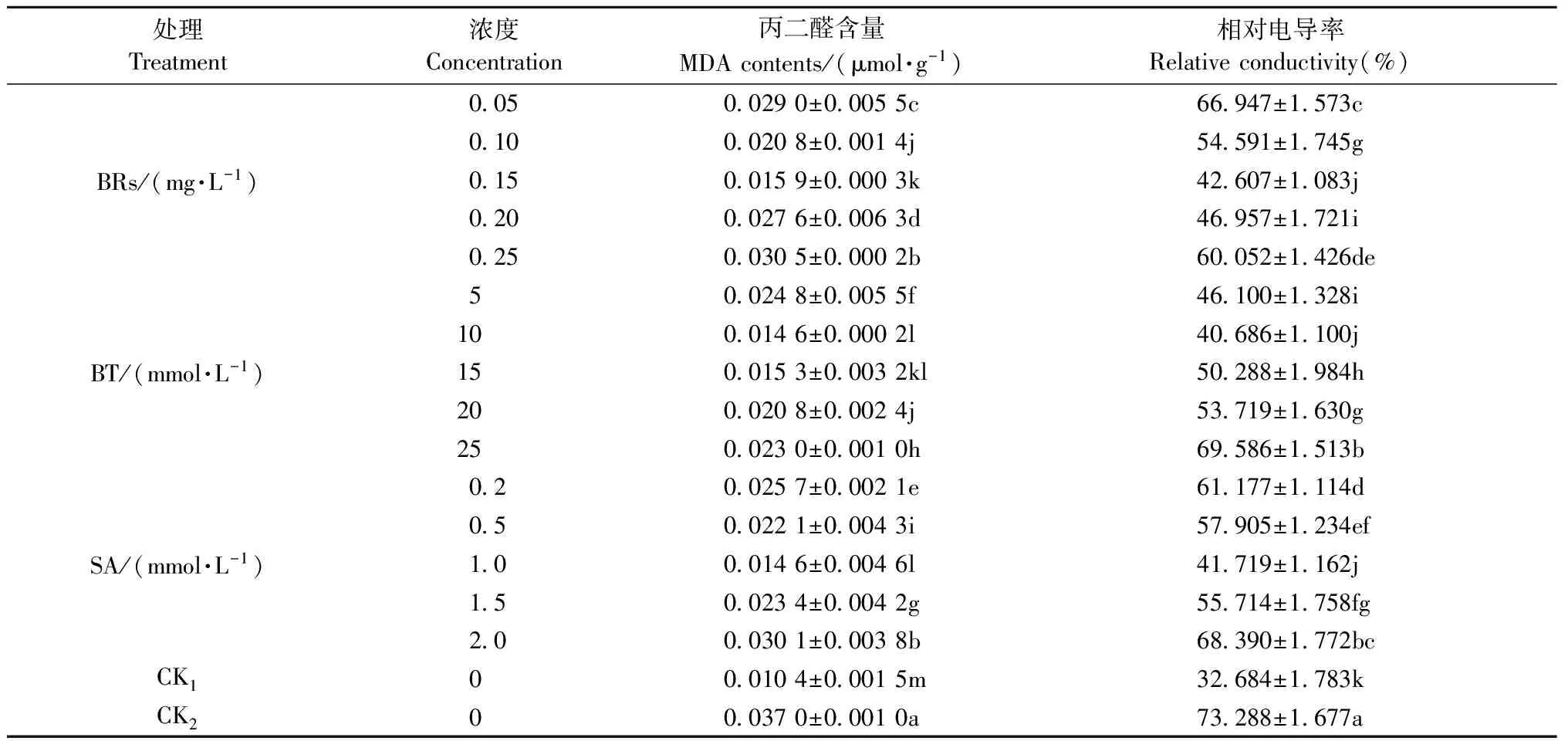
表1 生長調節劑對低溫脅迫下酸橙幼苗葉片MDA含量和相對電導率的影響①
① 同一列不同英文字母表示差異顯著(P<0.05)。下同。The different letters within the same column mean the significance at the 0.05 level.The same below.
2.2 低溫脅迫下生長調節劑對酸橙幼苗葉片抗氧化酶活性的影響
從表2可知,與CK1相比,在低溫脅迫下CK2酸橙葉片POD、SOD酶活性顯著降低,分別降低33.17%和13.51%,表明低溫脅迫顯著抑制POD、SOD酶活性。相對于低溫處理CK2來說,葉面噴施生長調節劑顯著提高酸橙幼苗葉片抗氧化酶POD、SOD和CAT的活性,且均隨著生長調節劑濃度的增加呈現先升高后降低的趨勢。與CK2相比,酸橙幼苗葉片抗氧化酶POD、SOD和CAT活性均以0.15 mg·L-1BRs、10 mmol·L-1BT、1 mmol·L-1SA處理提高效果最為顯著。噴施0.15 mg·L-1BRs,酸橙幼苗葉片POD、SOD和CAT酶活性分別提高49.31%,11.07%,27.6%,噴施10 mmol·L-1BT,酸橙幼苗葉片POD、SOD和CAT酶活性分別提高58.29%,13.34%,27.71%,噴施1 mmol·L-1SA,酸橙幼苗葉片POD、SOD和CAT酶活性分別提高67.00%,15.86%,26.51%。
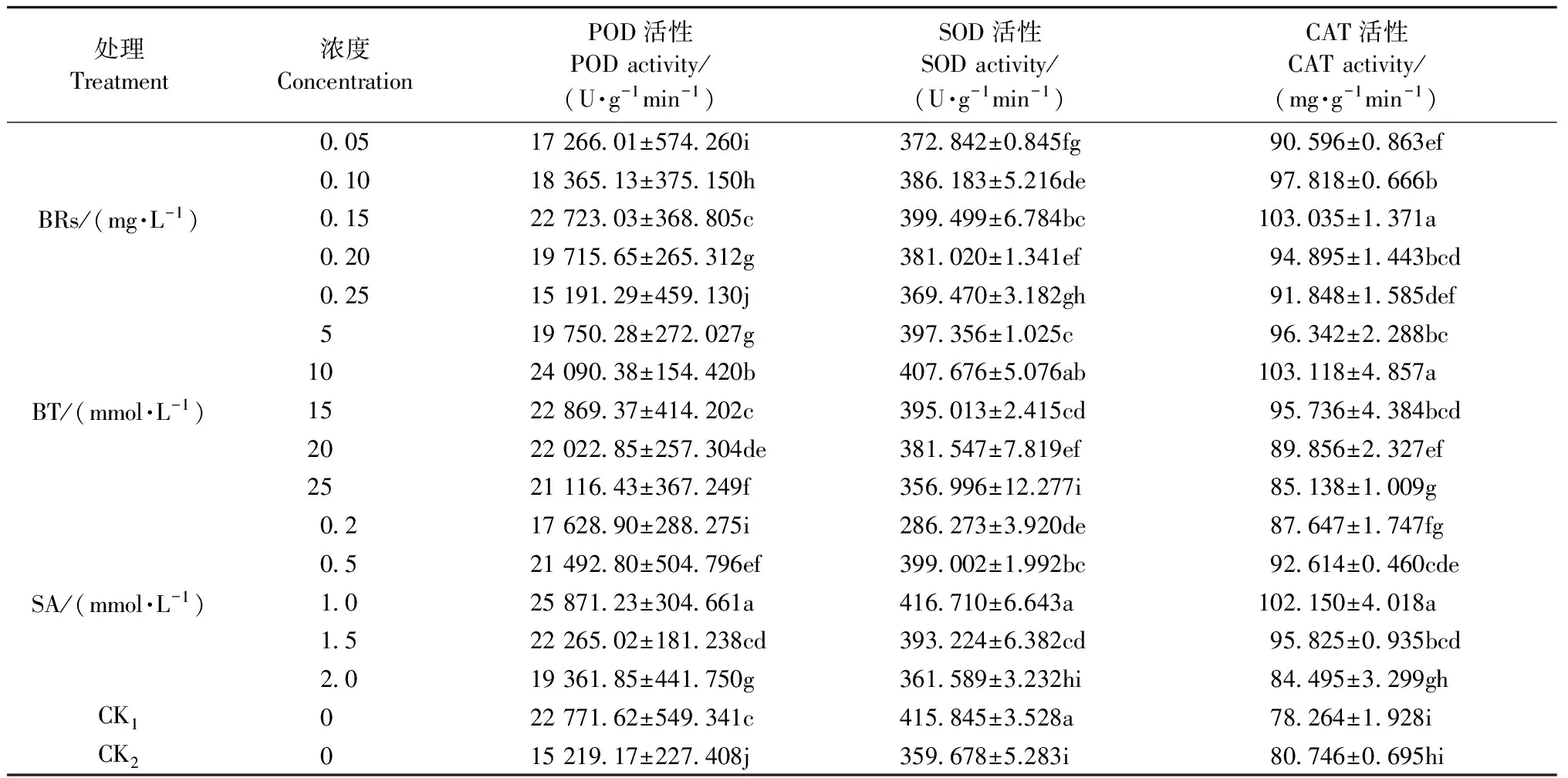
表2 生長調節劑對低溫脅迫下酸橙幼苗葉片抗氧化酶活性的影響
2.3 低溫脅迫下生長調節劑對酸橙幼苗葉片滲透調節物質含量的影響
2.3.1 脯氨酸含量變化 脯氨酸是植物重要的抗逆生理指標之一,參與植物細胞的滲透調節過程。從表3可知,酸橙幼苗經低溫脅迫處理后,葉片中脯氨酸含量較CK1顯著增加。說明低溫脅迫下酸橙幼苗能自發產生某種適應低溫脅迫的機制,誘導并積累脯氨酸,緩解低溫傷害。相對于CK2,葉面噴施不同濃度的生長調節劑可明顯提高酸橙幼苗葉片脯氨酸含量。經0.15 mg·L-1BRs、10 mmol·L-1BT、1 mmol·L-1SA處理的酸橙幼苗,葉片脯氨酸含量顯著提高33.22%,29.44%,28.41%。
2.3.2 可溶性蛋白含量變化 可溶性蛋白也是植物體內重要的滲透調節物質。從表3可知,低溫脅迫下,酸橙葉片可溶性蛋白含量的變化趨勢與脯氨酸含量一致,可溶性蛋白含量相對于CK1明顯增加。隨著BRs、BT、SA濃度的增大,酸橙幼苗葉片可溶性蛋白含量呈現先增加后降低的趨勢。相對于CK2,葉面噴施0.15 mg·L-1BRs,酸橙幼苗可溶性蛋白含量顯著提高34.43%;葉面噴施10 mmol·L-1BT,酸橙幼苗可溶性蛋白含量顯著提高65.20%;葉面噴施1 mmol·L-1SA,酸橙幼苗可溶性蛋白含量顯著提高67.88%。
2.4 低溫脅迫下生長調節劑對酸橙幼苗葉片葉綠素含量的影響
從表4可知,經低溫處理后,光合色素含量顯著下降,與CK1相比,CK2酸橙幼苗葉片中葉綠素a、葉綠素b、類胡蘿卜素和葉綠素總量分別顯著下降18.39%,18.78%,26.46%,18.46%。與CK2相比,葉面噴施不同濃度的生長調節劑能夠顯著減緩酸橙幼苗葉片葉綠素的下降趨勢,其中以0.15 mg·L-1BRs、10 mmol·L-1BT、1 mmol·L-1SA噴施效果最好。相對于CK2,葉面噴施0.15 mg·L-1BRs,類胡蘿卜素和葉綠素含量分別增加22.9%,18.67%;葉面噴施10 mmol·L-1BT,類胡蘿卜素和葉綠素含量分別增加25.18%,20.62%;葉面噴施1 mmol·L-1BRs,類胡蘿卜素和葉綠素含量分別增加34.98%,24.37%。
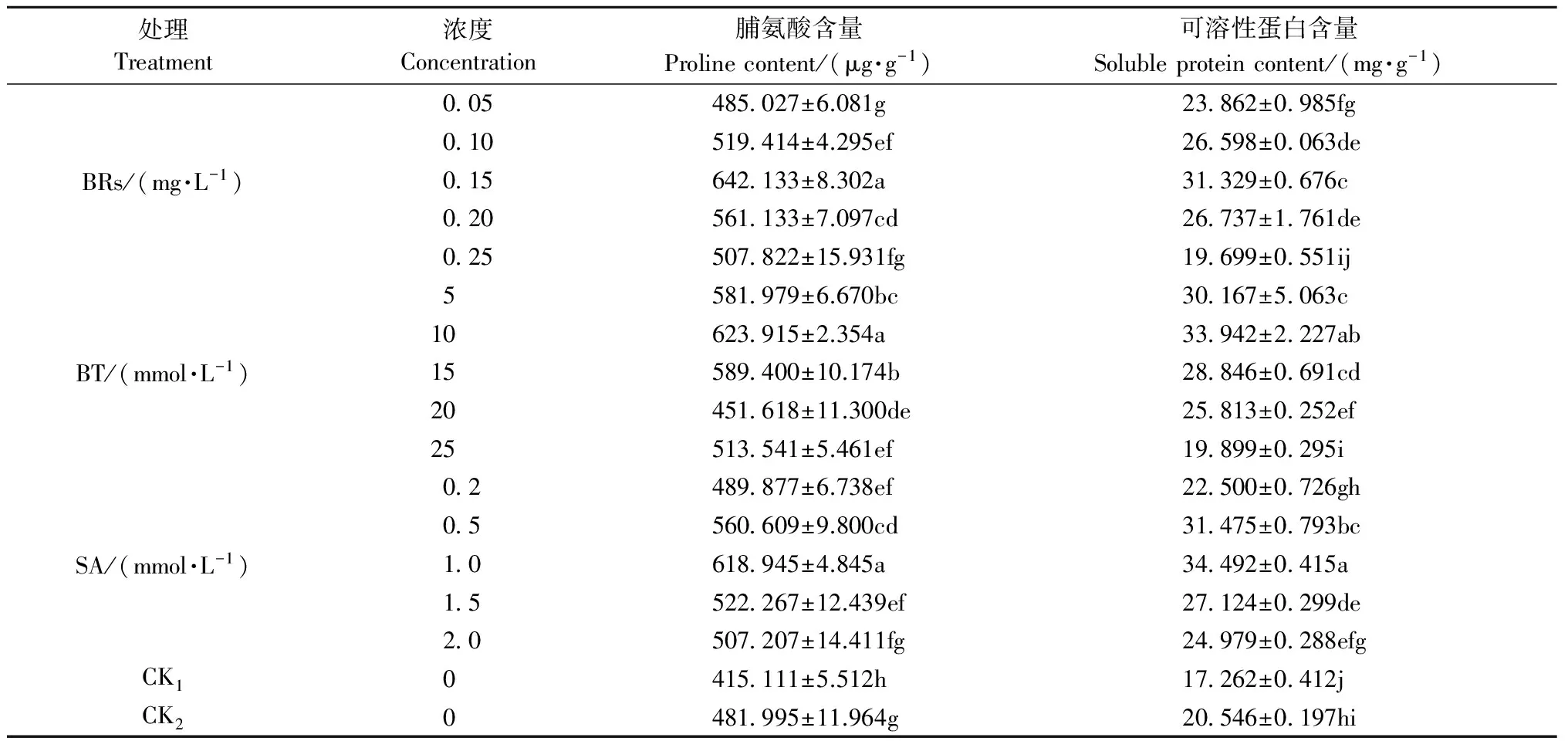
表3 生長調節劑對低溫脅迫下酸橙幼苗葉片滲透調節物質含量的影響
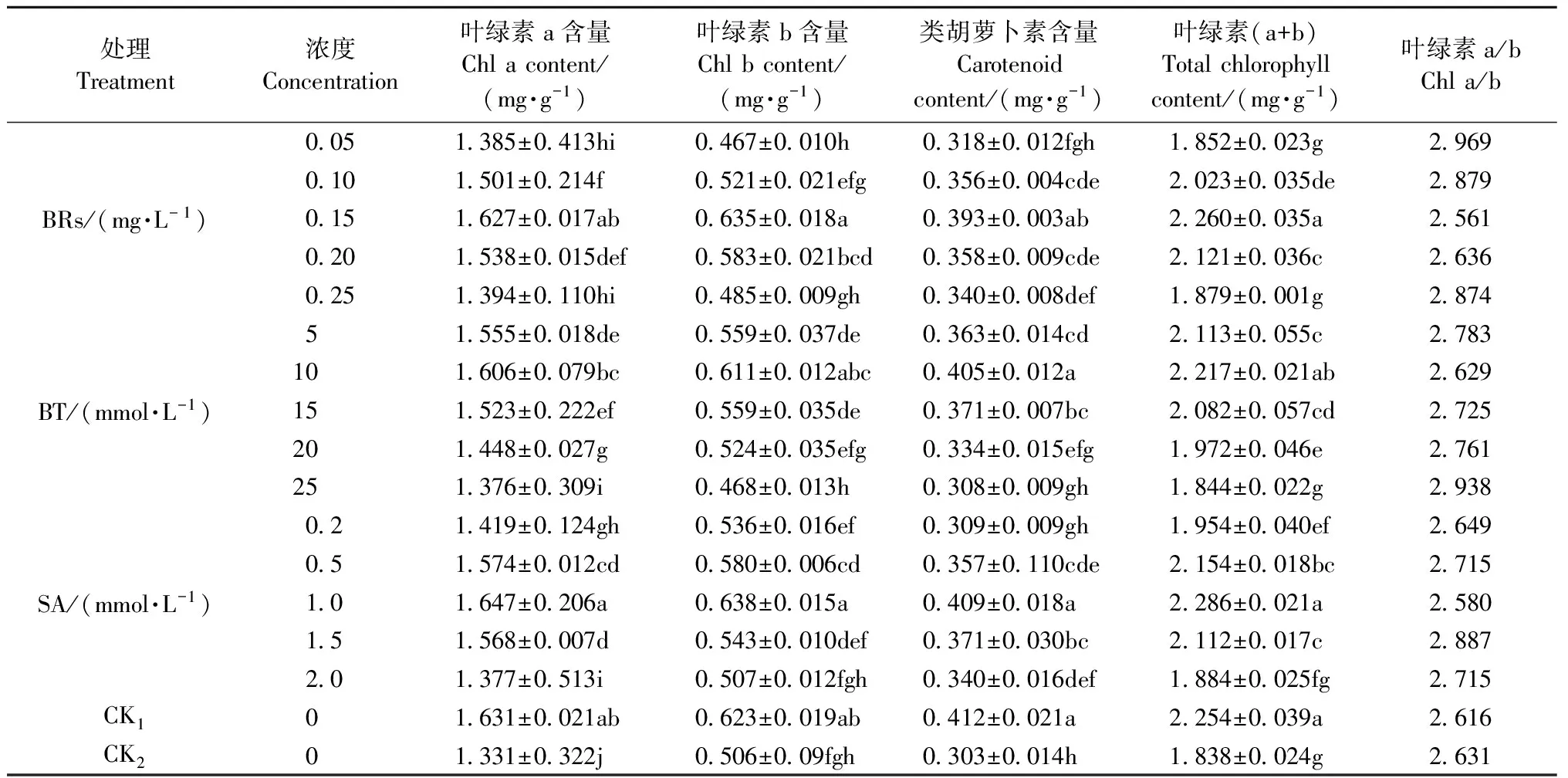
表4 生長調節劑對低溫脅迫下酸橙幼苗葉片葉綠素含量的影響
3 討論
3.1 低溫脅迫對酸橙幼苗葉片的傷害
3.2 生長調節劑對低溫脅迫下酸橙幼苗的滲透調節作用
低溫脅迫引起的植物水分損失與植物根吸收和運輸水分以及在隨后水分虧缺響應中關閉氣孔的能力下降有關(Bloometal., 2004)。植物體內缺水會引起植物葉片及嫩枝的萎蔫甚至死亡。本研究發現,酸橙幼苗頂部新葉在低溫處理第1天就出現萎蔫狀況,而下部葉片在低溫處理完成后仍沒有出現嚴重萎蔫,這可能是因為幼嫩的新葉較酸橙下部葉片對水分更加敏感。植物細胞缺水會促使植物主動積累脯氨酸和可溶性蛋白等滲透物質,以降低細胞滲透勢,增加細胞持水量。因此,低溫脅迫下植物體內滲透調節物質含量水平對于維持植物體內水分含量、減輕植物的低溫傷害具有重要意義。本研究發現,與CK2相比,低溫脅迫下用BRs、BT、SA處理的酸橙幼苗,其脯氨酸和可溶性蛋白含量顯著提高,進而可以推測SA可能誘導抗凍蛋白和脯氨酸的合成和積累(Chenetal., 1993a),而BRs可能減少ROS的產生(Yuetal., 2002),保護細胞內合成酶類等生物大分子,產生與BT一樣的保護作用(郭啟芳等, 2004)。3種生長調節劑作用的最終結果都是使細胞內脯氨酸和可溶性蛋白含量增加,減少細胞水分損失,同時穩定酸橙幼苗葉片細胞膜結構與功能,減輕膜脂過氧化程度,顯著降低MDA含量和相對電導率,提高酸橙幼苗的抗寒性。
3.3 生長調節劑對低溫脅迫下酸橙幼苗抗氧化酶活性的影響
SOD、POD和CAT是植物體內清除ROS和自由基的抗氧化酶,是植物抵抗逆境的第一道防線。低溫環境下,植物細胞自由基的產生和清除遭到破壞而出現ROS的過量積累,這種積累超過一定閾值,就會破壞膜系統的結構與功能,發生膜脂過氧化,導致植物死亡(Liuetal., 2013)。因此,抗氧化酶活性越高,植物對逆境的適應能力就越強。本研究發現,葉面噴施BRs、BT、SA能夠顯著提高低溫脅迫下酸橙幼苗葉片SOD、POD和CAT抗氧化酶活性。因此,可以推斷3種生長調節劑可以提高酸橙幼苗的抗逆性,增強酸橙對低溫環境的抵抗力。3種生長調節劑對抗氧化酶活性的誘導效果不同,可能是因為其對酸橙的作用機制不同: BRs通過促進光合作用電子的運輸,減少自由基的產生和自由基對細胞傷害,使細胞代謝正常(Yuetal., 2002); 而SA可能是直接誘導酶活性的產生(Chenetal., 1993a); BT則是通過其對細胞內大分子的保護作用,保護了抗氧化酶和膜系統的結構,使其正常行使功能(郭啟芳等, 2004)。3種外源物質通過其特有的作用方式減少ROS的產生,提高抗氧化酶的活性,維持植物體內ROS代謝的平衡,進而減輕低溫對酸橙幼苗的傷害。這與前人的研究一致(Scandalioetal., 1997)。
3.4 生長調節劑對低溫脅迫下光合色素的保護作用
光合作用為植物生長和發育提供所需的物質和能量,是植物生長發育的基礎。低溫脅迫對植物光合色素含量、PSⅡ活性及光合能量代謝等一系列生理生化過程都有明顯的影響(曾乃燕等, 2000; 劉文英, 2015)。具體表現為,低溫會抑制光合作用碳反應的酶活性,使植物發生光抑制現象,進而影響植物的各種生理代謝過程(Alametal., 2002; Powles,2003)。長時間的低溫脅迫會加重酸橙幼苗的光抑制現象,導致PSⅡ復合體中的D1蛋白受損,致使PSⅡ活性中心被破壞(鄭彩霞, 2013)。同時,光抑制會破壞葉綠體雙層膜結構,引起光合色素的降解。本研究發現,與低溫脅迫CK2相比,噴施BRs、BT、SA能顯著提高酸橙幼苗葉片葉綠素和類胡蘿卜素含量,這可能是因為BRs能夠促進光合電子向卡爾文循環轉移,減少葉綠體中光能積累(Ogwenoetal., 2008); BT保護光合作用的碳反應酶和光合色素合成酶的結構(郭啟芳等, 2004),使碳反應能夠快速并順利地進行; SA誘導抗氧化酶產生和保護光合色素合成酶的活性。3種生長調節劑通過其對細胞調節作用,最終可減少酸橙幼苗葉片細胞中ROS含量,避免ROS對葉綠體的破壞,在促進光合色素合成的同時,保護光合色素的結構,使細胞內光合色素的含量維持在一個較高水平。同時,類胡蘿卜素等光保護介質的增加,可加快過剩光能的耗散(Demmigetal., 1992),減少D1蛋白受損機率,保護光合系統。
4 結論
低溫是影響泰州地區酸橙生長的重要環境因子之一,在酸橙栽培過程中應采取相應措施降低酸橙樹苗的低溫傷害。本研究發現,在低溫來臨之前,葉面噴施油菜素內酯(BRs)、甜菜堿(BT)、水楊酸(SA)能夠降低低溫脅迫下酸橙幼苗MDA含量,提高滲透調節物質含量,減輕葉綠素下降趨勢,顯著提高SOD、POD、CAT等多種抗氧化酶活性,并最終提高苗木在低溫脅迫下的抗逆性。BRs、BT、SA處理酸橙幼苗的適宜濃度分別為0.15 mg·L-1,10 mmol·L-1,1 mmol·L-1。
郭啟芳,馬千全,孫 燦,等.2004.外源甜菜堿提高小麥幼苗抗鹽性的研究.西北植物學報, 24(9):1680-1686.
(Guo Q F, Ma Q J, Sun C,etal. 2004.Effects of exogenous glycinebetaine on the salt-tolerance of wheat. Acta Botanica Boreali-Occidentalia Sinica, 24(9):1680-1686. [in Chinese])
康國章,歐志英,王正詢,等.2003.水楊酸誘導提高香蕉幼苗耐寒性的機制研究.園藝學報, 30(2):141-146.
(Kang G Z, Ou Z Y, Wang Z X,etal. 2003.Salicylic acid alleviated the damage caused by low temperature to cell membrane and some photosynthetic functions of banana seedlings. Acta Horticulture Sinica, 30(2):141-146. [in Chinese])
李合生.2000.植物生理生化實驗原理和技術. 北京: 高等教育出版社, 134-263.
(Li H S. 2000.Principles and techniques of biochemical and physiological experiments for plants. Beijing: Higher Education Press, 134-263. [in Chinese])
梁小紅,安勐潁,宋 崢,等.2015.外源甜菜堿對低溫脅迫下結縷草生理特性的影響.草業學報, 24(9):181-188.
(Liang X H, An M Y, Song Z,etal. 2015. Effects of exogenous glycine betaine on the physiological characteristics ofZoysiajaponicaunder low-temperature stress. Acta Prataculturae Sinica, 24(9):181-188. [in Chinese])
劉文英.2015.植物逆境與基因.北京: 北京理工大學出版社, 41-76.
(Liu W Y. 2015.Plant stress and gene. Beijing: Beijing Institute of Technology Press, 41-76. [in Chinese])
劉 偉,艾希珍,梁文娟,等.2009.低溫弱光下水楊酸對黃瓜幼苗光合作用及抗氧化酶活性的影響.應用生態學報, 20(2):441-445.
(Liu W, Ai X Z, Liang W J,etal. 2009.Effects of salicylic acid on the leaf photosynthesis and antioxidant enzyme activities of cucumber seedlings under low temperature and light intensity. Chinese Journal of Applied Ecology, 20(2):441-445. [in Chinese])
宋金鳳,李 鋒,王亞軍,等.2012.低溫脅迫下外源有機酸對長白落葉松幼苗生理生化特性的影響.林業科學,48(2): 89-95.
(Song J F, Li F, Wang Y J,etal. 2012.Effects of exogenous organic acids on physiological and biochemical characteristics ofLarixolgensisseedlings under low temperature stress. Scientia Silvae Sinicae, 48(2): 89-95.[in Chinese])
曾乃燕,何軍賢,趙 文,等.2000.低溫脅迫期間水稻光合膜色素與蛋白水平的變化.西北植物學報, 20(1):8-14.
(Zeng N Y, He J X, Zhao W,etal. 2000.Changes in components of pigments and proteins in rice photosynthetic membrane during chilling stress. Acta Botanica Boreali-Occidentalia Sinica, 20(1):8-14. [in Chinese])
張 波,張懷剛.2005.甜菜堿提高植物抗鹽性的作用機理及其遺傳工程研究進展.西北植物學報, 25(9):1888-1893.
(Zhang B, Zhang H G. 2005.Mechanism for glycine betaine to improve plant salt resistance and its research advances in genetic engineering. Acta Botanica Boreali-Occidentalia Sinica, 25(9): 1888-1893. [in Chinese])
鄭彩霞.2013.植物生理學.北京: 中國林業出版社, 125-160.
(Zheng C X. 2013. Plant physiology. Beijing: China Forestry Publishing House, 125-160. [in Chinese])
鄭國華,張賀英,鐘秀榮.2009.低溫脅迫下枇杷葉片細胞超微結構及膜透性和保護酶活性的變化.中國生態農業學報,17(4): 739-745.
(Zheng G H, Zhang H Y, Zhong X R. 2009.Changes in cell ultra-structure, membrane permeability and protective enzyme activity inEriobotryajaponicaLindl. leaves under cold stress. Chinese Journal of Eco-Agriculture, 17(4): 739-745. [in Chinese])
Alam B, Jacob J. 2002. Overproduction of photosynthetic electrons is associated with chilling injury in green leaves. Photosynthetica, 40(1): 91-95.
Alscher R G, Erturk N, Heath L S. 2002. Role of superoxide dismutase (SOD) in controlling oxidative stress in plants. Journal of Experimental Botany, 53(372): 1331-1341.
Bloom A J, Zwieniecki M A, Passioura J B,etal. 2004.Water relations under root chilling in a sensitive and tolerant tomato species. Plant Cell and Environment, 27 (8): 971-979.
Chen Z, Ricigliano J W, Klessig D F. 1993a. Purification and characterization of a solube salicylic acid binding protein from tobacco. Pro Natl Acad Sci USA, 90(20): 9533-9537.
Chen Z, Silva H, Klessig D F. 1993b. Active oxygen species in the induction of plant systemic acquired resistance by salicylic acid. Science, 262(5141):1883-1886.
Demmig A B, Adams W W. 1992. Photoprotection and other responses of plant to high light stress. Annu Rev Plant Physiol Plant Mol Biol, 43(11): 599-607.
Hu W H, Wu Y, Zeng J Z,etal. 2010. Chill-induced inhibition of photosynthesis was alleviated by 24-epibrassinolide pretreatment in cucumber during chilling and subsequent recovery. Photosynthetica, 48 (4): 537-544.
Jung S, Steffen K L, Lee H J. 1998. Comparative photoinhibition of a high and a low altitude ecotype of tomato (Lycopersiconhirsutum) to chilling stress under high and low light condition. Plant Sci,134(1): 69-77.
Khripach V, Zhabinskii V, de Groot A. 2000. Twenty years of Brassinosteroids: Steroidal plant hormones warrant better crops for the XXI century. Ann Bot,86(3): 441-447.
Kratsch H A, Wise R R. 2000. The ultrastructure of chilling stress. Plant Cell and Environment, 23(4): 337-350.
Krishna P. 2003. Brassinoteroid-mediated stress responses. Plant Growth Regul, 22(4): 289-297.
Liu W Y, Yu K M, He T F,etal. 2013. The low-temperature induced physiological responses ofAvenanudaL.,a cold tolerant plant species. The Scientific World Journal, 2013(6):658-793.
Luo J P, Jiang S T, Pan L J. 2001. Enhanced somatic embryogenesis by salicylic acid ofAstragalusadsurgenspall.: relationship with H2O2production and H2O2-metabolizing enzyme activities. Plant Sci, 161(1): 125- 132.
Mittler R. 2002. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci, 7(9): 405-410.
Nilsen E T, Orcutt D M. 1996.The Physiology of Plants under Stress. New York,NY,USA:John Wiley & Sons, 70.
Ogweno J O, Song X S, Shi K,etal. 2008. Brassinosteroids alleviate heat-induced inhibition of photosynthesis by increasing carboxylation efficiency and enhancing antioxidant systems inLycopersiconesculentum. Plant Growth Regul, 27(1): 49-57.
Powles S B. 2003. Photoinhibition of photosynthesis induced by visible light. Annu Rev Plant Physiol Plant Mol Biol, 35(1): 15-44.
Scandalio J G. 1997. Oxidative stress and the molecular biology of antioxidant defenses. Woodbury,New York:Cold Spring Harbor Laboratory Press, 545-552.
Shirasu K, Nakajima H, Rajasekhar K. 1997. Salicylic acid potentiates an agonist-dependent gain control that amplifies pathogen signal in the activation of defense mechanisms. Plant Cell, 9(2): 261-270.
Tatsumi Y, Murata T. 1981.Relation between chilling sensitity of cucurbitaceae fruit and theme mbranceper meability.Society forHorticultural Science, 50 (1): 108-113.
Wang C, Ma X L, Hui Z,etal. 2008. Glycine betaine improves thylakoid membrane function of tobacco leaves under low-temperature stress. Photosyntheica, 46(3):400-409.
Weng J H, Jhaung L H, Jiang J Y,etal. 2006.Down-regulation of photosystem 2 efficiency and spectral reflectance in mango leaves under very low irradiance and varied chilling treatments. Photosynthetica,44(2): 248-254.
Yu J Q, Zhou Y H, Ye S F,etal. 2002. 2,4-epibrassinolide and abscisic acid protect cucumber seedling form chilling injury. Hort Sci Biotechnol, 77(14): 430-473.
Yu J Q, Huang L F, Hu W H,etal. 2004. A role for brassinosteroids in the regulation of photosynthesis inCucumissativus. Exp Bot, 55(399): 1135-1143.
(責任編輯 王艷娜 郭廣榮)
Effects of Exogenous Growth Substances on Physiological Traits of Cold Tolerance inCitrusaurantiumSeedlings
Liang Yongfu1Wang Kangcai1Xue Qi1Sui Li1Ye Jun2Chen Xingzhong2
(1.CollegeofHorticulture,NanjingAgriculturalUniversityNanjing210095; 2.ForestryTechnicalGuidanceStationofTaizhou,JiangsuProvinceTaizhou225300)
【Objective】 In order to reveal feasibility of different concentrations of BRs, BT and SA onCitrusaurantiumunder low temperature stress, the effects of BRs, BT and SA on physiological index were studied. 【Method】 PotC.aurantiumseedlings cultured under the same outdoor conditions were sprayed on the leaves with BRs, BT and SA separately, and then placed in the low incubator [0 ℃/4 ℃ (day/night), light intensity of 600 μmol·m-2s-1], and the seedlings sprayed with distilled water and set at 25 ℃ were served as control Ⅰ(CK1), and the seedlings sprayed with distilled water and set at low temperature stress (0 ℃/4 ℃, day/night) were served as control Ⅱ(CK2). The relative electric conductivity, contents of proline, malondialdehyde, soluble protein, chlorophyll, and activities of SOD, POD and CAT were measured. 【Result】 Compared with normal temperature CK1, low temperature significantly inhibit the antioxidant enzyme activity of CK2, enhanced the cell membrane damage and brought about loss of the normal functions of cell membrane. Thus, low temperature stress caused some increase in relative electric conductivity and MDA content ofC.aurantiumseedlings. Low temperature stress treatment also destroyed the structure of the chloroplast, causing the decomposition of photosynthetic pigment and reducing the content of chlorophyll and carotenoid. Compared with CK2, 0.15 mg·L-1BRs, 10 mmol·L-1BT, 1 mmol·L-1SA enhanced the activities of POD, SOD, and CAT, increased the contents of proline and soluble protein, and effectively reduced the damage of low temperature stress on cell membrane. At the same time, the exogenous BRs , BT and SA increased the contents of chlorophyll and carotenoid, the efficiency of leaf photosynthesis and thus effectively resisted the oxidative stress caused by low temperatures. But with the increasing concentration of spraying, the ability ofC.aurantiumto resist low temperature stress decreased. 【Conclusion】 The foliage spraying with BRs, BT and SA could reduce the damage of low temperature stress onC.aurantiumseedling leaves.
Citrusaurantiumseedlings; low temperature stress; exogenous substances; stress resistance
10.11707/j.1001-7488.20170308
2016-01-14;
2016-03-28。
江蘇省林業三新工程(LYSX[2014]48)。
S718.43
A
1001-7488(2017)03-0068-08
*王康才為通訊作者。
