楊樹GRF1/2d負調控不定根的發生和發育*
2017-04-27 06:44:18周厚君魏凱莉趙巖秋宋學勤盧孟柱
林業科學 2017年3期
周厚君 魏凱莉 江 成 趙巖秋 宋學勤 盧孟柱
(林木遺傳育種國家重點實驗室 中國林業科學研究院林業研究所 北京100091)
?
楊樹GRF1/2d負調控不定根的發生和發育*
周厚君 魏凱莉 江 成 趙巖秋 宋學勤 盧孟柱
(林木遺傳育種國家重點實驗室 中國林業科學研究院林業研究所 北京100091)
【目的】 生長調節因子(GRFs)是一類植物特有的轉錄因子,調控植物生長發育的多個生物學過程。研究楊樹組織和器官發育中GRFs的作用,尤其是對不定根形成的調控,不僅可以豐富根發育的理論,而且對于楊樹的扦插繁殖具有實際應用價值。【方法】 從銀腺楊84K中分離了PtGRF1/2d基因和其啟動子,通過對其miR396靶位點核苷酸進行同義突變,獲得不受miR396調控的突變形式的mPtGRF1/2d,并將啟動子和該突變形式分別構建至含有GUS報告基因的植物表達載體和過量表達載體,通過遺傳轉化分別獲得PPtGRF1/2d∷GUS啟動子驅動GUS轉基因楊樹和mPtGRF1/2d過表達轉基因楊樹。通過GUS染色分析PtGRF1/2d楊樹啟動子的表達特性,并對過表達mPtGRF1/2d楊樹不定根的發生時間、數目和長度進行統計,利用qRT-PCR分析不定根發育早期相關轉錄因子的表達。【結果】 PtGRF1/2d主要在根的中柱鞘和根尖位置表達,說明其參與了根的形成; 過量表達mPtGRF1/2d基因影響了楊樹不定根的發生、發育,導致了不定根發生延遲、數目和長度均減少,且差異均達到顯著水平或極顯著水平,表明PtGRF1/2d對不定根的發生和發育具有負調控作用。qRT-PCR分析顯示, 過表達mPtGRF1/2d楊樹的不定根發育相關基因PtSCR,PtAIL9,PtBBM2,PtPLT1.2和PtWOX11b的表達量均被下調,表明PtGRF1/2d的過量表達抑制了根原基發生和不定根發育相關的關鍵調控因子的表達,影響了根原基的發生和不定根的形成,導致不定根數目和長度的變化,最終影響了楊樹的生長。【結論】PtGRF1/2d作為不定根形成的負調控因子,位于不定根調控途徑的上游,通過下調促進不定根形成的相關轉錄因子的表達來抑制根原基形成和不定根發育,導致不定根發生延遲、數目和長度減少。
PtGRF1/2d; 銀腺楊84K; 過量表達; 不定根; 負調控
生長調節因子(growth-regulating factor, GRF)是一類植物特有的轉錄因子,對植物的生長發育具有重要的調控作用(Kimetal., 2003; 2015; Zhangetal., 2008; Bazinetal., 2013; Wangetal., 2014; Omidbakhshfardetal., 2015)。GRFs基因通常在幼嫩的組織中表達量較高,在成熟的組織中表達量較低(Kimetal., 2003),提示GRFs基因可能參與植物的早期發育。大多數GRF是miR396的靶基因,與miR396一起參與多種植物生長發育過程,包括根的生長(Kimetal., 2015; Omidbakhshfardetal., 2015)。擬南芥(Arabidopsisthaliana)中大多數AtGRF在根中顯示出較高的表達水平(Baoetal., 2014),而且調控它們的miR396對根正常發育至關重要(Hewezietal., 2012)。干涉miR396a擬南芥根比野生型的長,而miR396a和miR396b過表達植株以及過表達不受miR396調控形式的AtGRF1或AtGRF3(靶位點堿基誘變)植株的根均變短(Hewezietal., 2012; Baoetal., 2014)。同樣,研究人員發現苜蓿(Medicagotruncatula)中MtGRF2、MtGRF4和MtGRF6的RNAi植株表現出根的長度比野生型短的表型(Bazinetal., 2013),而在miR396過表達苜蓿中,分生組織細胞數目變少、細胞周期特征基因表達量降低和處于復制期的細胞比例降低,表明miR396-GRF通過影響細胞周期途徑實現對根發育的調控(Bazinetal., 2013)。與大多數擬南芥AtGRFs基因在根中顯現出較高的表達水平不同的是,水稻(Oryzasativa)OsGRFs基因在根中低表達(Baoetal., 2014)。與此相反,大白菜(Brassicapekinensis)BrGRF16基因在根中高表達,表明它可能參與根的發育(Omidbakhshfardetal., 2015)。這些結果表明在根發育方面 GRFs的功能存在多樣性和差異性。
GRFs對根發育影響的研究主要集中在草本植物,在木本植物生長發育特別是不定根(adventitious roots)的發生和發育中的作用目前尚缺乏研究。不定根對木本植物的無性繁殖至關重要,也是決定繁殖成功與否的一個關鍵步驟。不定根的形成是一個復雜的過程,通常有4個時期: 激活期、誘導期、根原基的激活及新組織的形成期和根原基伸長及維管連接建立期。前期研究表明,不定根的這些過程都離不開轉錄因子的調控(Leguéetal., 2014)。轉錄因子SCARECROW(SCR)、APETALA2/ETHYLENE RESPONSE FACTOR(AP2/ERF)蛋白家族AINTEGUMENTA-like亞組[如PLETHORA(AtPLT),BABY BOOM(AtBBM),AINTEGUMENTA(AtANT)和 AINTEGUMENTA-like(AtAIL)]及WUSCHEL-related homeobox(WOX)(Krizeketal., 2000; Mizukamietal., 2000; Nole-Wilsonetal., 2005; Iminetal., 2007; Horstmanetal., 2014; Liuetal., 2014; Xuetal., 2015)均與不定根形成相關。楊樹不定根形成過程中會發生重要的轉錄組重塑,其中35個家族的轉錄因子的表達發生了顯著變化(Ramírez-Carvajaletal., 2009; Rigaletal., 2012; Leguéetal., 2014),表明楊樹不定根的形成同樣與轉錄因子的調控密切相關。但對楊樹不定根發生和形成的調控機制還缺乏認識。
楊樹作為研究樹木生長發育的模式樹種,研究其不定根的形成和形成過程中的分子調控機制,對研究樹木不定根的分子遺傳具有非常重要的理論指導意義。本研究從銀腺楊84K(Populusalba×P.glandulosa‘84K’)中分離了擬南芥AtGRF1和AtGRF2同源基因PtGRF1/2d,并對miR396靶位點的核苷酸進行同義突變,獲得了不受miR396調控的突變形式mPtGRF1/2d,通過遺傳轉化獲得過量表達mPtGRF1/2d的轉基因楊樹,并對其不定根的發生、發育進行分析,為了解GRF調控楊樹不定根的形成奠定基礎。
1 材料與方法
1.1 材料
銀腺楊84K組培苗為中國林業科學研究院林木遺傳育種國家重點實驗室保存并擴繁; pDNOR222.1、pCAMBIA1300和pMDC164載體、大腸桿菌(Escherichiacoli)DH5α及農桿菌(Agrobacteriumtumefaciens)GV3101菌種均由本實驗室保存; PCR引物合成和測序由英濰捷基生物公司完成; KOD PLUS酶購自TOYOBO公司; 重組酶購自 Invitrogen公司;TaqDNA聚合酶購自TaKaRa公司; 限制性內切酶購自NEB公司; DNA凝膠回收試劑盒、質粒提取試劑盒購自AxyGen公司; RNA提取試劑盒、DNA酶購自QIAGEN公司; 反轉錄試劑盒購自Invitrogen公司; 定量試劑購自TaKaRa公司。
1.2 方法
1.2.1 質粒和載體構建 利用擬南芥AtGRF1和AtGRF2基因序列在毛果楊(Populustrichocarpa)基因組數據庫(http://phytozome.jgi.doe.gov/pz/portal.html)進行比對,獲取擬南芥AtGRF1和AtGRF2同源基因PtGRF1/2d序列和其啟動子序列,據序列設計基因特異PCR引物(表1)。通過RT-PCR方法從銀腺楊84K cDNA中分離得到PtGRF1/2d基因,并在不改變其編碼氨基酸的基礎上,將PtGRF1/2d中miR396靶位點處6個核苷酸進行突變,使得mPtGRF1/2d與miR396成熟序列不能嚴格配對,從而避免在轉錄水平被miR396降解,獲取不受miR396調控的突變形式mPtGRF1/2d,將其克隆至pDNOR222.1載體,測序驗證后,重組至pCAMBIA1300過表達載體。從銀腺楊84K基因組DNA中分離PtGRF1/2d基因的啟動子,克隆至pDNOR222.1載體并重組至帶有GUS報告基因的pMDC164植物表達載體。將構建好的載體電擊轉化至農桿菌GV3101感受態細胞,利用農桿菌介導的葉盤法轉化銀腺楊84K。
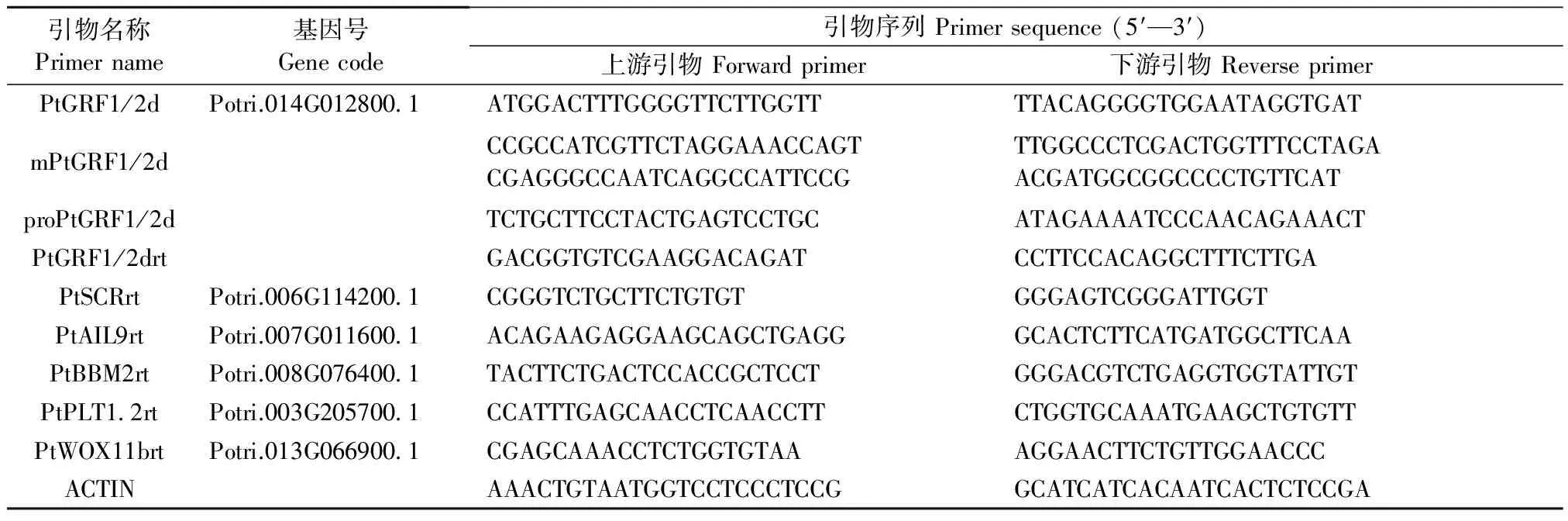
表1 PtGRF1/2d基因克隆和qRT-PCR所用引物序列
1.2.2 植物材料培養和遺傳轉化 植物材料培養和遺傳轉化具體方法見文獻(Liuetal., 2014)。選取表達量較高的3個過表達轉基因株系,以野生型銀腺楊84K為對照,每個株系至少擴繁15株,定植溫室。過量表達mPtGRF1/2d對楊樹株高影響的測定至少重復3次。
1.2.3 RNA提取和RT-PCR分析 組培30天過表達mPtGRF1/2d楊樹的葉用于PtGRF1/2d表達量鑒定; 組培扦插4天時處于不定根發育愈傷組織形成階段(Liuetal., 2014)的不定根發生部位,具體為培養基插入部分約1 cm的下端莖段,用于不定根發育相關基因的表達量鑒定。以上表達鑒定均采用實時定量的方法且至少重復3次。RNA提取和反轉錄具體方法見Liu等(2014),定量PCR的引物使用Primer 3軟件(http://frodo.wi.mit.edu/primer3/input.htm)設計(序列見表1)。實時定量PCR用SYBR Premix Ex TaqTM Ⅱ試劑盒(TaKaRa,Dalian,China)在Roche 480定量PCR儀(Roche Applied Science)上完成,每個樣品4次重復,ACTIN基因作內參。
1.2.4 GUS染色 組培30天的PPtGRF1/2d∷GUS轉基因楊樹用于GUS染色。GUS的組織化學染色具體方法見文獻(Liuet al., 2014)。
1.3 數據統計
用MicrosoftExcel和SPSS17.0軟件對數據進行處理,并用t檢驗法檢驗差異顯著性水平。
2 結果與分析
2.1 GRF1/2d在不定根中的表達情況
不定根的形成包括R1預誘導階段、R2愈傷組織形成階段、R3不定根出現階段和R4不定根延長階段(圖1A)(Liuet al., 2014)。銀腺楊84K不定根發育過程中,R1-R2時期PtGRF1/2d的轉錄活性先急劇下降后逐漸上升,R2時期PtGRF1/2d轉錄表達活性達到最強; 而R3-R4時期PtGRF1/2d轉錄表達急劇下降到起始時期水平(圖1B)。PtGRF1/2d的表達量在R2愈傷組織出現時期顯著上調(圖1B),提示它可能是根分生組織(根原基發生)的關鍵調控因子。
為進一步考察PtGRF1/2d的組織表達情況,創制了啟動子融合GUS報告基因的PPtGRF1/2d∷GUS轉基因材料。結果顯示GUS信號在葉、莖和根組織中都存在(圖1C)。進一步觀察發現,PtGRF1/2d在根部的信號集中在中柱鞘和根尖位置(圖1C:b,c),這與其調控根分生組織活性的假設相一致。
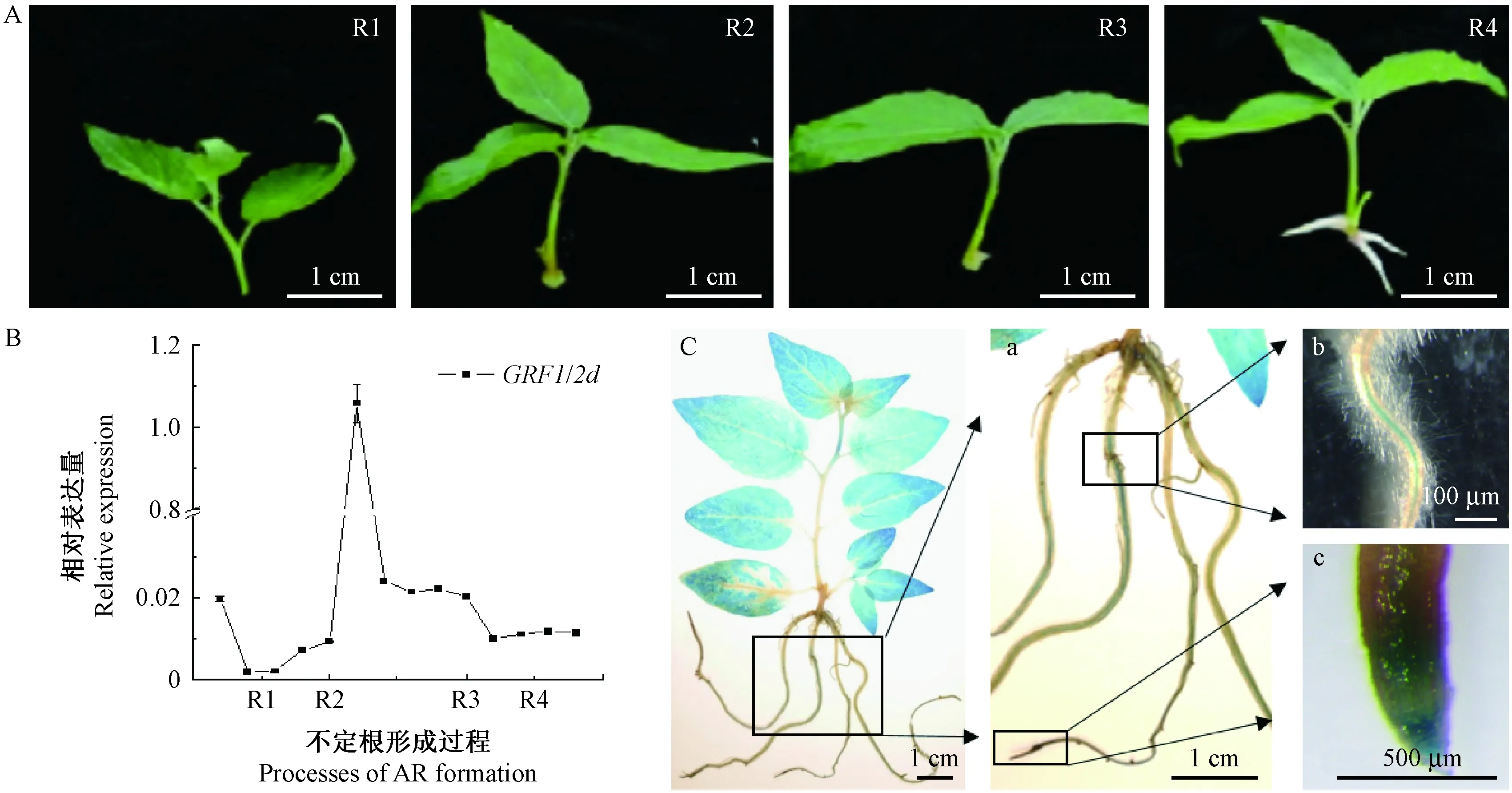
圖1 不定根中PtGRF1/2d的表達情況Fig.1 Expression of PtGRF1/2d during the successive stages of adventitious root (AR) formationA: 不定根形成過程(R1: 預誘導階段; R2: 愈傷組織形成階段; R3: 不定根出現階段; R4: 不定根延長階段); B: PtGRF1/2d動態表達變化; C: PPtGRF1/2d∷GUS楊樹GUS染色(a-c: 局部放大圖)。A: The successive stages of adventitious root formation(R1: Preinduction stage; R2: Callus regeneration stage; R3: Adventitious root emergence stage; R4: Adventitious root elongation stage); B: Dynamic expression of PtGRF1/2d during the successive stages of adventitious root formation; C: Total PPtGRF1/2d∷GUS poplar plant GUS staining(a, b and c represent partial enlarged drawing of picture C).
2.2 過表達mPtGRF1/2d對楊樹不定根發生、數目和長度的影響
為進一步研究PtGRF1/2d對楊樹不定根發育的影響,對miR396靶位點核苷酸進行同義突變,創制了不受miR396調控的過表達mPtGRF1/2d楊樹。從獲得的25個獨立轉基因株系中選取表達量不同的3個獨立轉基因株系(#8、#9和#10)作為進一步研究對象。
表型觀察發現,過表達mPtGRF1/2d楊樹根系發育明顯變弱(圖2A),具體表現為: 3個株系不定根的發育明顯延遲(圖2C); 且不定根數目減少,分別為3.1,3.6,2.8,對照楊樹為3.9(圖2D),不定根數目減少了10%~30%,其中2個株系與對照植株的差異達到顯著水平(*,P<0.05); 此外,3個株系不定根的長度變短,分別為4.241,4.174,4.236 cm,對照楊樹則為5.24 cm,不定根長度減少了20%左右,差異均達到極顯著水平(**,P<0.01)(圖2E)。以上結果表明,PtGRF1/2d可能是不定根發育的負調控因子。
不定根發育受多種轉錄因子調控(Leguéetal., 2014)。為了進一步明確PtGRF1/2d與這些不定根發育相關調控因子SCR,AIL,BBM2,PLT1以及WOX11的關系,利用qRT-PCR技術對這些基因的表達量進行了檢測。結果顯示,在不定根形成的起始階段,楊樹相應轉錄因子PtSCR,PtAIL9,PtBBM2,PtPLT1.2以及PtWOX11b基因的表達量在過表達mPtGRF1/2d楊樹中被顯著下調(圖2F),表明PtGRF1/2d在不定根發育的起始階段發揮作用,可能是通過影響與根原基形成及不定根發育的相關轉錄因子的表達,負調控楊樹不定根的形成和伸展。
生長90天的過表達mPtGRF1/2d楊樹株高生長明顯低于對照(圖3A)。統計結果顯示: 過表達mPtGRF1/2d楊樹3個株系株高分別為22.17,23.83,26.00 cm,而對照為36.5 cm(圖3B),3個株系的株高均低于對照,達到顯著水平(P<0.05)或極顯著水平(P<0.01),表明PtGRF1/2d對楊樹不定根發生和發育的負調控所導致的不定根數目及長度的減少,嚴重影響了楊樹的生長。
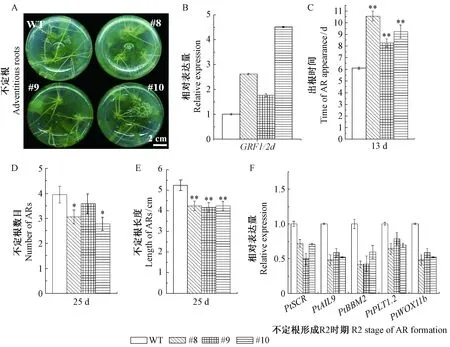
圖2 過表達mPtGRF1/2d楊樹的不定根相關性狀分析Fig.2 Analysis of adventitious root (AR) traits of mPtGRF1/2d overexpression transgenic poplarsWT: 對照植株; #8, #9, #10: 過表達mPtGRF1/2d楊樹。WT: Wide type; #8, #9, #10: mPtGRF1/2d overexpression transgenic poplars. t-test: *, P < 0.05; **, P < 0.01.下同。The same below.

圖3 溫室定植90天的對照植株(WT)和過表達mPtGRF1/2d楊樹(#8, #9, #10)Fig.3 Ninety-day-old wide type(WT) and mPtGRF1/2d overexpression transgenic poplars (#8, #9, #10) in greenhouse
3 討論
根作為重要的植物器官,解析生長調節因子(GRFs)對根的發育影響非常重要。但目前這類研究工作主要集中在擬南芥、苜蓿、水稻等草本植物中,在木本植物生長發育中的作用研究很少。決定木本植物扦插繁殖一個重要方面就是不定根的發生和發育,因此本研究在木本植物楊樹中分析GRFs的作用,對于揭示楊樹不定根發生、發育的調控機制及其在扦插繁殖技術上的應用具有重要的意義。
本研究克隆了楊樹PtGRF1/2d基因,表達分析顯示其在不定根發育的起始階段即根原基形成時期表達變化顯著(圖1B); 啟動子驅動的GUS染色分析表明PtGRF1/2d在不定根的中柱鞘和根尖高表達(圖1C: b, c),表明該基因在根的形成和發育中的重要作用。通過對miR396靶位點的核苷酸進行同義突變,獲取不受miR396調控的突變形式mPtGRF1/2d,用于獲得該基因的高表達轉基因植株。過表達mPtGRF1/2d楊樹生長受到抑制,主要是其不定根發育延遲,并且數目和長度均減少(圖2)。上述結果說明,GRF對楊樹不定根的影響與對其他植物如擬南芥根發育的影響一致,都具有負調控作用。
為了揭示其負調控作用的機制,利用qRT-PCR分析了過表達mPtGRF1/2d楊樹不定根發育早期與不定根形成有關的轉錄因子PtSCR,PtAIL9,PtBBM2,PtPLT1.2及PtWOX11b基因的表達。研究表明,SCR對不定根根原基形成過程中分生組織決定和維持有重要功能,而根原基的建立伴隨PtAIL1,PtAIL9,PtBBM2,PtPLT1.1和PtPLT1.2基因表達量的上調,根原基的分化則與PtAIL1,PtAIL5,PtAIL9,PtBBM2,PtPLT1.1和PtPLT1.2的表達調控相關(Leguéetal., 2014)。另外,WOX11a和WOX11b同樣參與不定根的發育(Liuetal., 2014; Xuetal., 2015)。過表達PtoWOX11/12a楊樹不定根數目增多(Liuetal., 2014),而過表達PeWOX11a和PeWOX11b的楊樹不僅不定根的數目增加,莖上還會誘導出不定根,此外過表達PeWOX11b楊樹還會在葉腋誘導出不定根和不定根上誘導出芽(Xuetal., 2015)。本研究發現,在過表達mPtGRF1/2d楊樹中其中5個基因的表達量被下調(圖2F),表明PtGRF1/2d的過量表達抑制了根原基發生和不定根發育的關鍵調控因子的表達。其中在過表達mPtGRF1/2d楊樹中,WOX11b的表達下調2倍,表明PtGRF1/2d可通過抑制WOX11b表達影響根原基的分化,起到負調控作用。值得注意的是,PtGRF1/2d能夠抑制多個根原基發生與不定根形成相關轉錄因子的轉錄,來抑制根原基的發生,延緩不定根的形成,最終導致不定根數目與長度的變化。這說明PtGRF1/2d位于不定根調控途徑的上游,具有重要的調控作用。
4 結論
本研究通過轉基因技術,分析了PtGRF1/2d在不定根發生和發育中的作用。結果表明PtGRF1/2d是楊樹不定根形成的負調控因子,主要通過下調能夠促進不定根形成的相關轉錄因子的表達,來抑制根分生組織和根原基的形成,最終導致不定根發生延遲、數目和長度減少。PtGRF1/2d如何抑制不定根轉錄因子,從而負調控不定根形成的具體機制有待進一步深入解析。
Bao M, Bian H, Zha Y,etal. 2014.miR396a-mediated basic helix-loop-helix transcription factor bHLH74 repression acts as a regulator for root growth inArabidopsisseedlings. Plant and Cell Physiology, 55(7): 1343-1353.
Bazin J, Khan G A, Combier J P,etal. 2013.miR396 affects mycorrhization and root meristem activity in the legumeMedicagotruncatula. The Plant Journal, 74(6): 920-934.
Hewezi T, Maier T R, Nettleton D,etal. 2012. TheArabidopsismicroRNA396-GRF1/GRF3 regulatory module acts as a developmental regulator in the reprogramming of root cells during cyst nematode infection. Plant Physiology, 159(1): 321-335.
Horstman A, Willemsen V, Boutilier K,etal. 2014. AINTEGUMENTA-LIKE proteins: hubs in a plethora of networks. Trends Plant Sci, 19: 146-157
Imin N, Nizamidin M, Wu T,etal. 2007. Factors involved in root formation inMedicagotruncatula. J Exp Bot, 58: 439-451.
Kim J H, Choi D, Kende H. 2003. TheAtGRFfamily of putative transcription factors is involved in leaf and cotyledon growth inArabidopsis. The Plant Journal, 36(1): 94-104.
Kim J H, Tsukaya H. 2015. Regulation of plant growth and development by the GROWTH-REGULATING FACTOR and GRF-INTERACTING FACTOR duo. Journal of Experimental Botany, 66(20): 6093-6107.
Krizek B A, Prost V, Macias A. 2000. AINTEGUMENTA promotes petal identity and acts as a negative regulator of AGAMOUS. Plant Cell, 12: 1357-1366.
Legué V, Rigal A, Bhalerao R P. 2014. Adventitious root formation in tree species: involvement of transcription factors. Physiologia Plantarum, 151(2): 192-198.
Liu B, Zhang J, Wang L,etal. 2014. A survey ofPopulusPIN-FORMED family genes reveals their diversified expression patterns. Journal of Experimental Botany, 65(9): 2437-2448.
Mizukami Y, Fischer R L. 2000. Plant organ size control: AINTEGUMENTA regulates growth and cell numbers during organogenesis. Proc Natl Acad Sci USA, 97: 942-947.
Nole-Wilson S, Tranby T L, Krizek B A. 2005.AINTEGUMENTA-like(AIL) genes are expressed in young tissues and may specify meristematic or division-competent states. Plant Mol Biol, 57: 613-628.
Omidbakhshfard M A, Proost S, Fujikura U,etal. 2015. Growth-regulating factors (GRFs): a small transcription factor family with important functions in plant biology. Molecular Plant, 8(7): 998-1010.
Ramírez-Carvajal G A, Morse A M, Dervinis C,etal. 2009. The cytokinin type-B response regulator PtRR13 is a negative regulator of adventitious root development inPopulus. Plant Physiology, 150(2): 759-771.
Rigal A, Yordanov Y S, Perrone I,etal. 2012. The AINTEGUMENTA LIKE1 homeotic transcription factor PtAIL1 controls the formation of adventitious root primordia in poplar. Plant Physiology, 160(4): 1996-2006.
Wang F, Qiu N, Ding Q,etal. 2014. Genome-wide identification and analysis of the growth-regulating factor family in Chinese cabbage (BrassicarapaL. ssp.pekinensis). BMC Genomics, 15(1): 1.
Xu M, Xie W, Huang M. 2015. TwoWUSCHEL-related HOMEOBOX genes,PeWOX11aandPeWOX11b, are involved in adventitious root formation of poplar. Physiologia Plantarum, 155(4): 446-456.
Zhang D F, Li B, Jia G Q,etal. 2008. Isolation and characterization of genes encoding GRF transcription factors and GIF transcriptional coactivators in Maize (ZeamaysL.). Plant Science, 175(6): 809-817.
(責任編輯 徐 紅)
NegativeRegulationofGRF1/2dontheFormationandDevelopmentofAdventitiousRootsinPopulus alba × P. glandulosa ‘84K’
ZhouHoujunWeiKailiJiangChengZhaoYanqiuSongXueqinLuMengzhu
(State Key Laboratory of Tree Genetics and Breeding Research Institute of Forestry, Chinese Academy of Forestry Beijing 100091)
【objective】 Growth-regulating factors (GRFs) are plant-specific transcription factors that are involved in various developmental events, especially in root development. It is interesting to investigate the roles of GRFs in growth and development of tissues and organs, rooting in particular, in woody plants, which would not only increase our knowledge on the mechanisms of the regulation of adventitious root formation and also be useful for cutting propagation in woody plants. 【Method】PtGRF1/2dgene and its promotor were isolated fromPopulusalba×P.glandulosa‘84K’. The overexpression vector of miR396-resistant construct 35S:mPtGRF1/2d-GFPand the promoter-testing vector ofPPtGRF1/2d∷GUSwasgenerated.ThemiR396-resistantversionofmPtGRF1/2dcontainedsixsilentmutationswithinthemiR396-complementarydomainofthePtGRF1/2dgenomicclone,therebyincreasingthenumberofmismatchesbetweenmiR396andmPtGRF1/2dwithoutalteringtheaminoacidsequenceoftheencodedPtGRF1/2dprotein. mPtGRF1/2doverexpressiontransgenicpoplarsandPPtGRF1/2d∷GUStransgenicpoplarswereobtainedrespectively.ExpressioncharactersinPPtGRF1/2d∷GUStransgenicpoplarsandtimeofadventitiousrootappearance,thenumberandlengthofadventitiousrootsinoverexpressiontransgenicpoplarswereanalyzed.TheexpressionofrelatedtranscriptfactorsatearlystageofadventitiousrootformationinoverexpressiontransgenicpoplarswereanalyzedbyqRT-PCR. 【Result】TheresultsshowedthattheexpressionofPtGRF1/2dwasmainlyinthepericycleandtipofroot,suggestinggeneralinvolvementofPtGRF1/2dintherootdevelopment.ThedevelopmentofadventitiousrootsinmPtGRF1/2doverexpressiontransgenicpoplarswassignificantlyaffectedincomparisontothenon-transgenicpoplarsusedascontrols.OverexpressionofPtGRF1/2dcausednotonlythedelayedinitiationofadventitiousrootsbutalsothereducednumberandlengthofadventitiousroots,whichindicatesthatPtGRF1/2dexhibitsanegativeregulationoninitiationanddevelopmentofadventitiousroot.qRT-PCRanalysisindicatedthattheexpressionof5transcriptfactorspreviouslyshowninvolvementinadventitiousrootformation,includingPtSCR, PtAIL9, PtBBM2, PtPLT1.2andPtWOX11b,wasdown-regulatedatearlystageofadventitiousrootformationinoverexpressiontransgenicpoplars,whichdemonstratedthatoverexpressionofPtGRF1/2dsuppressedtheexpressionofkeyfactorsforadventitiousrootformationthusresultedinthedelayedformationofadventitiousrootsandthereducednumberandlengthofadventitiousroots,andfinallyinfluencedthegrowthofpoplar.【Conclusion】PtGRF1/2disanegativeregulatorofpoplaradventitiousrootformation,functioningintheupstreamofadventitiousrootformationregulationpathway,down-regulatestheexpressionofadventitiousrootrelatedfactorsforadventitiousrootformationthussuppressestheinductionoftherootprimordiumandadventitiousrootformation,andfinallyleadstothedelayedadventitiousrootsappearanceandthereducednumberandlength.
PtGRF1/2d; Populus alba × P. glandulosa ‘84K’;overexpression;adventitiousroot;negativeregulation
10.11707/j.1001-7488.20170304
2016-04-25;
2016-06-17。
國家自然科學基金面上項目“生長調節因子PtGRF10參與楊樹次生生長調控的機理研究”(31570676)。
S
A
1001-7488(2017)03-0033-07
*盧孟柱為通訊作者。
