山東稀有植物小果白刺天然群體表型變異研究
2017-04-19 08:45:02魯儀增楊海平栗寧寧解孝滿
林業科學研究 2017年2期
董 昕,王 磊,魯儀增,楊海平,韓 義,王 倩,栗寧寧,解孝滿
(1.山東省林木種質資源中心, 濟南 250014; 2.山東農業大學, 泰安 271018)
山東稀有植物小果白刺天然群體表型變異研究
董 昕1,王 磊1,魯儀增1,楊海平1,韓 義1,王 倩2,栗寧寧1,解孝滿*
(1.山東省林木種質資源中心, 濟南 250014; 2.山東農業大學, 泰安 271018)

小果白刺;天然群體;種實;葉片;表型;變異

小果白刺是山東自然分布的重要耐鹽植物,主要分布于東營、濱州、濰坊等濱海鹽堿地。近年來,由于城市建設和鹽堿地開發等原因,小果白刺的分布范圍急劇減小,生境趨于破碎化,現被列為山東(暖溫帶)稀有植物,如不加強保護,許多地方的小果白刺天然資源將會消亡。因此,搞清山東小果白刺天然群體的變異程度及分布特征,對于保護和利用這一珍稀樹種意義重大。目前,對于小果白刺的研究主要涉及育苗[7-9]、造林方法[10-11]、生理生化特性[12]及果實成分[13-14]的測定,有關其表型多樣性的研究鮮有報道。
本文對山東境內5個小果白刺天然群體種實和葉片的表型性狀進行比較分析,以期揭示山東小果白刺表型變異程度與變異規律,為小果白刺資源保護與合理利用提供依據。
1 材料與方法
1.1 材料來源及采樣方法

表1 小果白刺各取樣群體生境概況
1.2 表型性狀的測定
選取能夠表征小果白刺性狀且容易獲得的指標進行測量。對于種實指標,用游標卡尺測量果實橫徑(FTD)、果實縱徑(FLD)、種子橫徑(STD)、種子縱徑(SLD),每份隨機測量30粒,測量值保留3位小數,計算果徑縱比橫(FLD/FTD)和種徑縱比橫(SLD/STD)。用電子天平測量種子重量,重復3次,以百粒法計算種子千粒重(SM),結果保留2位小數。對于葉片指標,用直尺測量葉片長(LL)、葉片寬(LW),每份隨機測量30片葉,測量值保留2位小數,計算葉片長/寬(LL/LW)。
1.3 統計分析方法
對于各性狀值采用單因素隨機區組方法進行方差分析。以方差分量表示群體間和群體內表型分化程度,以各方差分量與總分量的比值作為表型分化系數:

2 結果與分析
2.1 小果白刺群體間和群體內的表型變異特征
由表2可知,小果白刺10個表型性狀在群體間和群體內差異均達到顯著或極顯著水平,說明小果白刺的表型性狀在群體間和群體內均存在廣泛的變異。
綜上,小果白刺群體間果實、種子和葉片的表型指標值差異較大,均達到顯著或極顯著水平,但相同群體的果實大小和種子大小沒有呈現一一對應關系。
2.2 小果白刺群體間、群體內表型性狀的形態變異
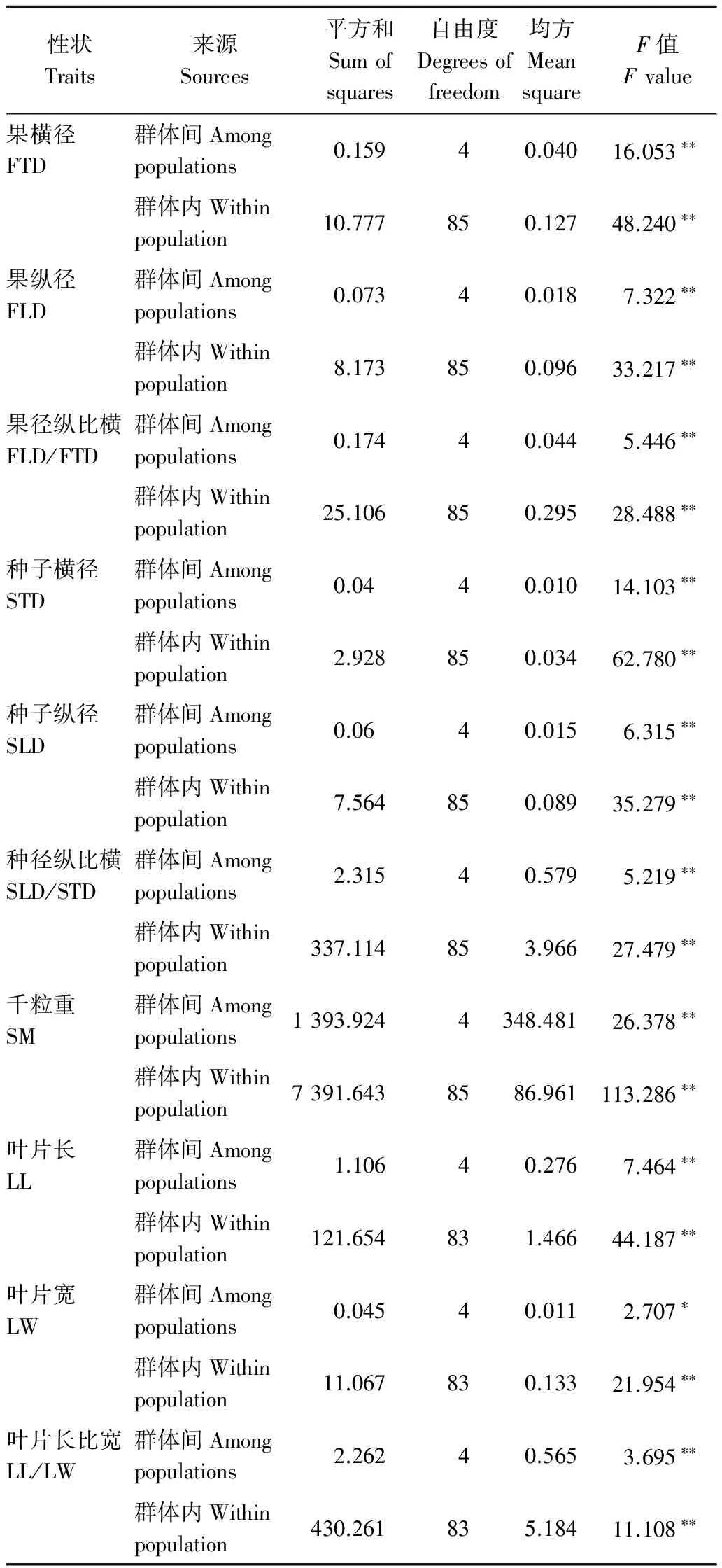
表2 小果白刺群體種實及葉片表型性狀方差分析
注:*表示差異顯著(p<0.05),**表示差異極顯著(p<0.01)。
Note: FTD, fruit transverse diameter; FLD, fruit longitudinal diameter; FLD/FTD, ratio of fruit longitudinal diameter to transverse diameter; STD, seed transverse diameter; SLD, seed longitudinal diameter; SLD/STD, ratio of seed longitudinal diameter to longitudinal diameter; SM,1000-seed weight; LL, leaf length; LW, leaf width; LL/LW, ratio of leaf length to width;*p<0.05; **p<0.01.

表3 小果白刺群體種實及葉片表型性狀變異(平均值±標準偏差)
2.3 小果白刺群體間的表型分化

表4 小果白刺群體間、群體內種實及葉片表型性狀的變異系數
2.4 小果白刺表型性狀的相關分析
以個體為單位進行表型性狀的相關分析,結果見表6。小果白刺10個果實、種子和葉片性狀中多數呈顯著或極顯著的相關關系,這些性狀是能夠代表小果白刺種實及葉片特征的重要性狀。這些性狀中,除了表征相同器官部位的性狀自相關外,表征不同器官部位的性狀也存在一定的相關關系。果實橫徑、果實縱徑分別與種子橫徑、種子縱徑、種子千粒重呈極顯著的正相關關系,表明小果白刺的果實越大,則種子越大,種子質量也越大;葉長、葉寬、葉長比寬分別與種子橫徑、種子縱徑、種徑縱比橫、種子千粒重呈顯著或極顯著的相關關系,說明小果白刺營養生長與生殖生長具有較為密切的聯系。
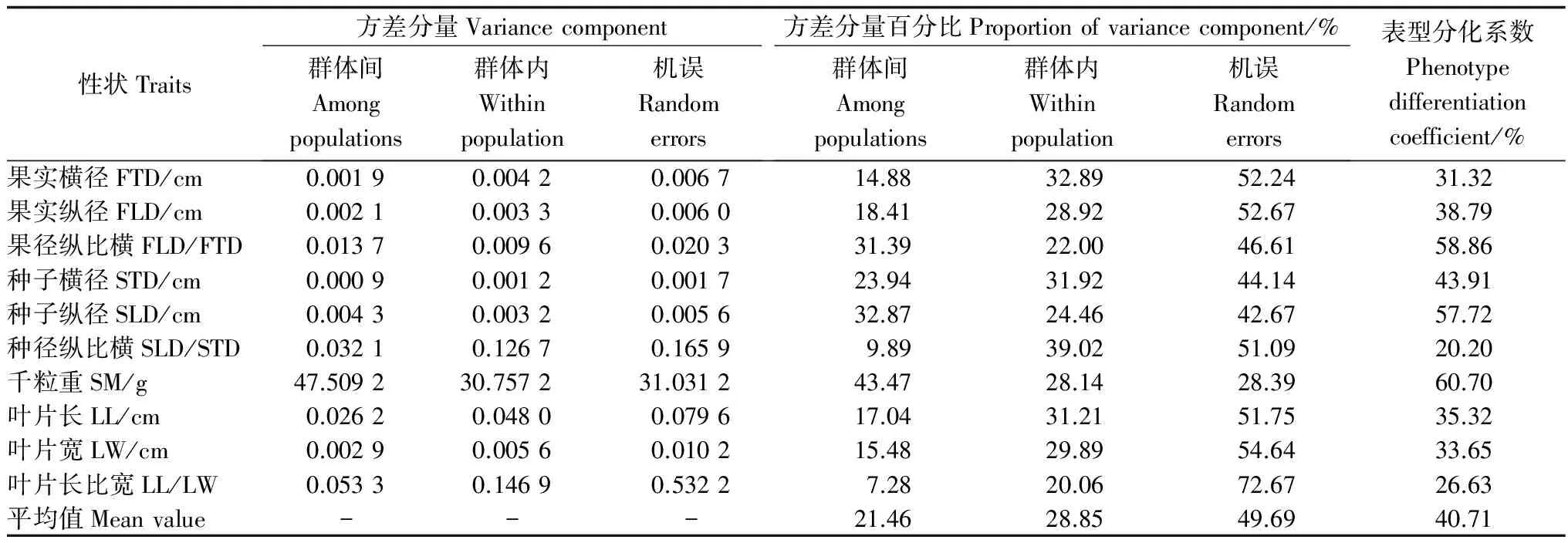
表5 小果白刺群體間的方差分量及表型分化系數
2.5 小果白刺表型性狀的地理變異
以群體為單位進行地理變異分析。由表6可知,除了果徑縱比橫和種子縱徑這兩個性狀分別與經度和緯度地理因子相關性顯著外,其他大部分性狀受地理因子影響較小,這與小果白刺的研究尺度和其在山東的分布范圍較小有關。其中,種子縱徑與經度因子呈顯著的正相關,表明越往東分布的群體,其種子縱徑越大。果徑縱比橫與緯度因子呈顯著的負相關,表明在小果白刺生境內,越往北分布的群體其果實縱徑與橫徑相差的越小,果實形狀越接近圓形。同時,果徑縱比橫受水分生態因子影響較大,與年均降水量呈極顯著的正相關關系,表明水分越大的地方,小果白刺群體果實縱徑有比橫徑大得越多的趨勢,果實形狀越趨于長圓形。日照時數對果實橫徑具有較明顯的影響,二者呈顯著的正相關關系,表明日照相對充足的地方,其果實橫徑越大。然而,樹種天然分布區內環境因素往往是復雜多樣的,其生長生活不只是受到單一環境因子的影響,因此,山東小果白刺群體的地理變異表現為隨機變異模式。
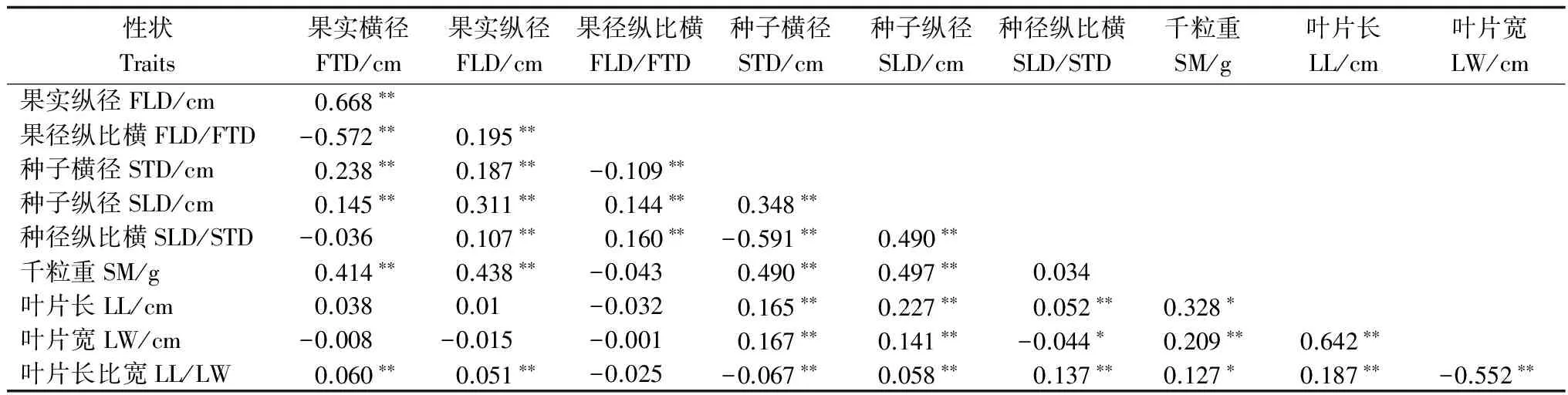
表6 小果白刺群體種實及葉片表型性狀間的相關分析
注:*表示差異顯著(p<0.05),**表示差異極顯著(p<0.01)。
Note:*p<0.05; **p<0.01.
2.6 小果白刺表型性狀的聚類分析
采用組間聯接法對小果白刺5個群體進行系統聚類分析,生成聚類樹系圖,見圖1。在閾值為23.90處,6個小果白刺群體被劃分成2類,第Ⅰ類包括沾化、無棣、孤島和壽光4個群體,第Ⅱ類由河口群體獨自組成。在閾值為15.11處,第Ⅰ類可再劃分成兩個亞類,第ⅰ亞類包括沾化、無棣、孤島3個群體,第ⅱ亞類僅包含壽光群體。通過聚類分析被劃分在同一類的群體其地理上并不都是相連接的(例如孤島和壽光群體),而且在地理上分布距離較近的群體沒有被優先聚合成一類(例如孤島和河口群體),由此說明山東小果白刺不同群體間葉片和種實性狀的變異在空間上是不連續的。

表7 小果白刺種實及葉片表型性狀與地理生態因子的相關分析
注:*表示差異顯著(p<0.05),**表示差異極顯著(p<0.01)。
Note:*, p<0.05; **, p<0.01.
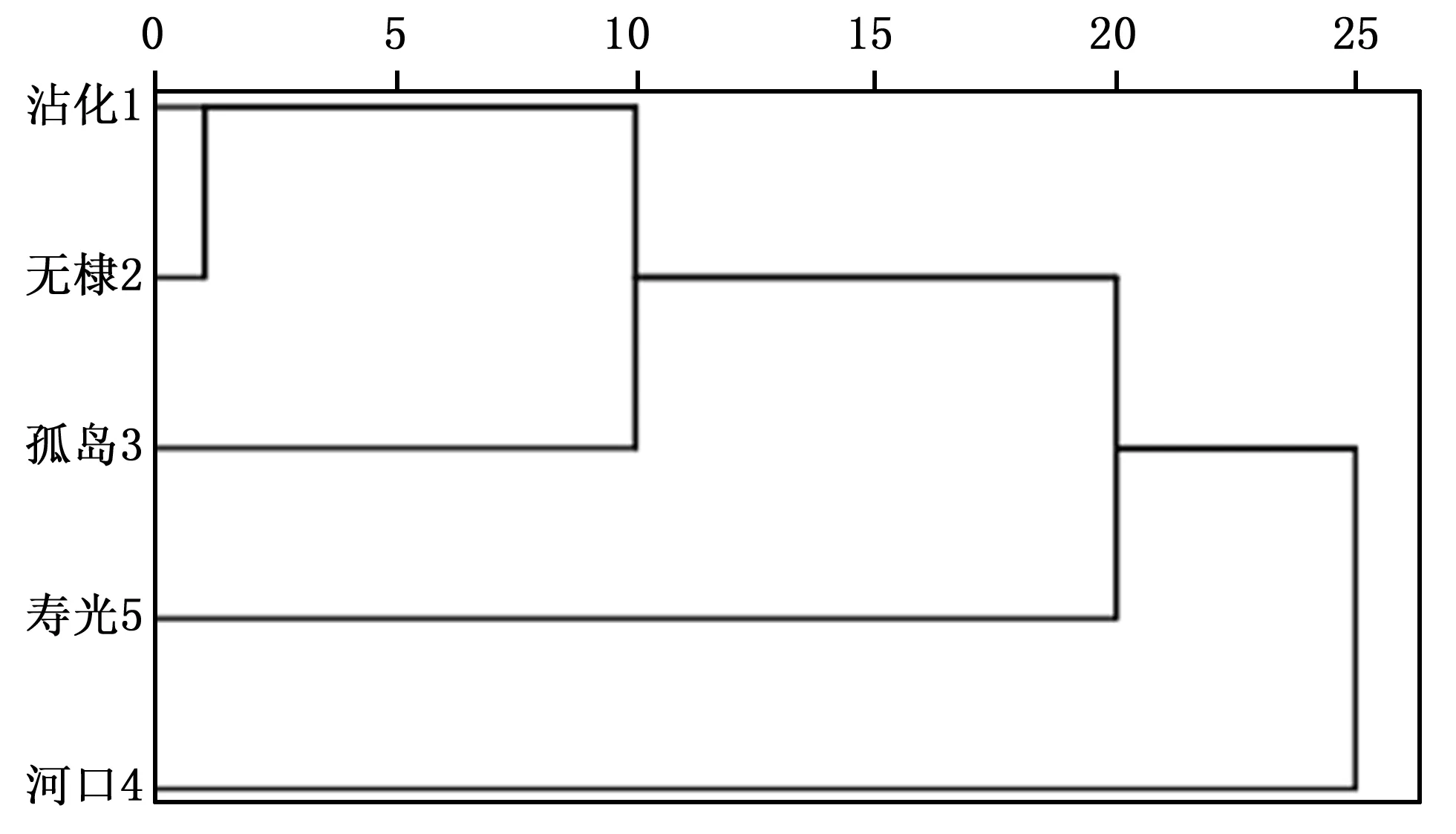
圖1 基于小果白刺群體種實及葉片表型性狀的聚類樹系圖Fig.1 UPGMA cluster based on the fruit and seed phenotypic traits of Nitraria sibirica Pall. populations
3 討論
3.1 山東小果白刺群體種實及葉片表型變異多樣性
對山東小果白刺5個群體的10個種實和葉片性狀調查研究發現,小果白刺在群體間和群體內存在顯著或極顯著的差異。對于各器官部位大小的變異程度有:葉片(8.99%)﹥種子(7.74%)﹥果實(6.77%),說明表征小果白刺果實大小的性狀更為穩定。對于各器官部位形態的變異程度有:種子(10.22%)﹥葉片(6.70%)﹥果實(4.35%),說明小果白刺葉片形狀是較穩定的性狀,表現為倒披針形,這與對青海高原西伯利亞白刺群體表型變異研究的結果正好相反[17]。分布在群體間的變異真正反映了群體在不同環境中的適應狀況,其大小在某種程度上說明了該生物對不同環境適應的廣泛程度,值越大,適應的環境越廣[18]。不同群體的相同性狀變異程度存在差異,這是由環境異質性導致的,各群體變異系數平均值從大到小排列順序為:孤島(18.52%)>沾化(17.66%)>壽光(17.57%)>無棣(16.74%)>河口(14.81%),說明孤島群體的表型多樣性最豐富,昌邑群體的表型多樣性最低。
對山東小果白刺表型性狀的變異來源進行定量分析,各性狀群體間表型分化系數平均值為40.71%,表明山東小果白刺天然群體表型變異群體間貢獻占40.71%,群體內貢獻占59.29%,群體內變異大于群體間變異,說明山東小果白刺的表型變異主要來源于群體內,群體內的多樣性大于群體間的。山東小果白刺的表型分化系數與其他具有防護作用的灌木樹種相比,分別大于康定柳(SalixpaaplesiaSchneid.)雄性群體(21.51%)[19]、康定柳雌性群體(27.32%)[19]、同為黃河三角洲的檉柳(Tamarixchinensis)(7.17%)[20]、檸條錦雞兒(Caragangkorshinskii)(19.50%)[21],小于剛毛檉柳(TamarixhispidaWilld.)(62.50%)[22]、山蒼子(Litseacubeba)(60.19%)[23],表明山東小果白刺群體間分化水平處于中等偏高的水平。然而,盡管山東小果白刺群體內的變異大于群體間的變異,但是群體間變異的意義卻遠大于群體內變異,因為存在于群體間的變異反映了地理、生殖隔離上的變異,群體間的多樣性變異是種內多樣性的重要組成部分[24]。這表明山東小果白刺種實和葉片性狀的群體間分化在一定程度上是由其本身的遺傳基礎決定的。山東小果白刺具有中等的群體間分化水平,可能與其交配系統和地理分布范圍雙層因素有關。一方面,小果白刺為蟲媒傳粉樹種,以種子繁殖為主要繁衍方式,與風媒傳粉的樹種相比,其花粉的傳播距離受到限制,從而導致群體間的基因流水平相對較低,這增加了小果白刺群體間分化的可能性;另一方面,小果白刺在山東的分布范圍比較集中,主要分布于黃河入海口及萊州灣附近的泥質海灘上,盡管近些年由于沿海灘涂開發力度加大,濱海植被遭到不斷地破壞,導致小果白刺資源呈現島嶼化、斑塊化的分布狀態,因地理隔離和遺傳漂變可能加劇了小果白刺的群體間分化,然而,在山東小果白刺的分布區內全部為平原地形,有利于種群基因交流,這會促進其種群基因頻率維持在較為穩定的水平,從而使群體間分化程度減弱。
3.2 山東小果白刺表型性狀的地理變異
山東小果白刺種實、葉片性狀與地理生態因子之間具有不同的相關性,是種實各部位對不同環境條件適應的外在表現。除了果實橫徑、果實形狀和種子縱徑性狀分別表現出受到日照時數、緯度和年均降水量、經度因子較大影響外,其他大部分性狀受地理生態因子的影響并不顯著,因此山東小果白刺群體的地理變異不是連續的,而是表現為隨機變異模式。通過聚類分析將5個小果白刺群體劃分成2類,然而,有些地理上相近的群體并沒有優先聚合成一類,這可能與群體間立地條件類型存在差異有關。
3.3 山東小果白刺種質資源的保護對策與意義
4 結論
綜合多種分析方法研究山東小果白刺天然群體種實和葉片表型變異,結果表明,其種實、葉片在群體間和群體內均存在著較為豐富的變異,這些變異是遺傳和環境共同作用的結果。群體內變異是山東小果白刺表型變異的主要來源。目前,山東小果白刺群體間分化水平與一些同類灌木相比處于中等偏上的水平,其地理變異遵照隨機變異模式。群體間和群體內多層次變異為山東小果白刺資源的保育和利用提供了物質基礎。
[1] 劉媖心. 中國植物志,第43卷(第1分冊)[M]. 北京: 科學出版社,1998: 118-122.
[2] 王國柱,王秉喆,黃雪峰,等. 蘇打鹽堿土區小果白刺造林效果的研究[J]. 吉林林業科技, 2009, 38(2): 11-13.
[3] 曾 杰,鄭海水,甘四明,等. 廣西西南樺天然居群的表型變異[J]. 林業科學, 2005, 41(2): 59-65.
[4] 楊 玲,沈海龍,等. 不同產區野生花楸果實和種子的型表多樣性[J]. 東北林業大學學報, 2009, 37(2): 8-10.
[5] 李文英,顧萬春. 蒙古櫟天然群體表型多樣性研究[J]. 林業科學, 2005, 41(1): 49-56.
[6] Naomi Chechowitz, Dorothy M. Chappell, Sheldon I. Guttman, Lee A. Weigt. Morphological, electrophoretic, and ecological analysis of Quercus macrocarpa populations in the Black Hills of South Dakota and Wyoming[J]. Canadian Journal of Botany, 1990, 68(10): 2 185-2 194. [7] 李振洲,陳 真,劉曉慧,等. 西伯利亞白刺播種育苗技術[J]. 北方園藝, 2011, (07): 71-72.
[8] 王德強,馬盈慧,杜海波,等. 優良堿地造林樹種西伯利亞白刺種苗繁育技術[J]. 農業科學, 2011, (4): 139.
[9] 張啟昌,李雙福,其其格. 西伯利亞白刺的微繁技術[J]. 東北林業大學學報, 2008, 36(4): 12-28.
[10] 徐 森. 西伯利亞白刺鹽堿地造林試驗[J]. 吉林林業科技, 2011, 40(4): 10-12.
[11] 王 麗,張英楠. 西伯利亞白刺人工栽培技術[J]. 吉林林業科技, 2010, 39(1): 47-51.
[12] 左鳳月,郝秀芬,陳占峰,等. 小果白刺和泡果白刺的耐鹽性[J]. 天津農學院學報, 2013, 20(2): 11-14.
[13] 王金梅,康文藝. 唐古拉特白刺和西伯利亞白刺葉揮發性成分研究[J]. 精細化工, 2009, 26(8): 774-780.
[14] 李永強,楊士花,朱仁俊,等. 西伯利亞白刺果實花色苷的初步鑒定[J]. 食品科學, 2009, 30(9): 77-79.
[15] Khalil M A K. Genetic variation in eastern white spruce (Piceaglauca(Moeneh) Voss) populations[J]. Can J For Res, 1985, 15(2): 444-452.
[16] 刁松鋒,邵文豪,姜景民,等. 基于種實性狀的無患子天然群體表型多樣性研究[J]. 生態學報, 2014, 34(6): 1451-1459.
[17] 張亞琦,王 寧,趙 越,等. 青海高原西伯利亞白刺天然居群表型多樣性研究[J]. 北方園藝, 2013, (9): 64-67.
[18] 龐廣昌,姜冬梅. 群體遺傳多樣性和數據分析[J]. 林業科學, 1995, 31(6): 543-550.
[19] 馮 毅,王朱濤,蔡應君,等. 川西北地區康定柳天然群體表型多樣性研究[J]. 西南林學院學報, 2010, 30(4): 11-15.
[20] 趙景奎,徐立安,解荷峰,等. 黃河三角洲檉柳群體遺傳多樣性RAPD分析[J]. 南京林業大學學報(自然科學版), 2008, 32(5): 56-60.[21] 王 贊,高新中,韓建國. 檸條錦雞兒表型多樣性研究[J]. 草地學報, 2006, 14(3): 201-205.
[22] 張 娟,尹林克,張道遠. 剛毛檉柳天然居群遺傳多樣性初探[J]. 云南植物研究, 2003, 25(5): 557-562.
[23] 田勝平,汪陽東,陳益存,等. 山蒼子天然種群葉片和種實性狀的表型多樣性[J]. 生態學雜志, 2012, 31(7): 1665-1672.
[24] Daniel L H, Andrew G C. Principles of Population Genetics[M]. Sinauer Associates: Inc., Sunderland, U.K..1997: 1-670.
(責任編輯:崔 貝)
Phenotypic Variation ofNitrariasibiricaNatural Populations in Shandong
DONGXin1,WANGLei1,LUYi-zeng1,YANGHai-ping1,HANYi1,WANGQian2,LINing-ning1,XIEXiao-man1
(1.Center for Forest Genetic Resource of Shandong Province, Ji’nan 250014, Shandong, China; 2.Shandong Agricultural University, Tai’an 271018,Shandong, China)
[Objective]To figure out the degrees and laws of phenotypic variation ofNitrariasibiricain Shandong Province. [Method]10 phenotypic traits about fruit and leaf of fiveN.sibiricapopulations in Shandong Province were analyzed by the methods of variance analysis, correlation analysis, and cluster analysis, etc.[Result]The variation within populations was proved as the main resource of the phenotypic variation. The phenotypic differentiation coefficients among the populations varied from 20.20% to 60.70%, with an average value of 40.71%. The average variation coefficient for all the traits was 9.34%. The relationship of the phenotypic variability degrees for different components are as follows: leaves size (8.99%) > seeds size (7.74%) > fruits size (6.77%) and seeds shape (10.22%) > leaves shape (6.70%) > fruits shape (4.35%). Most of those traits were significantly correlated with one another. The fiveN.sibiricapopulations were then clustered into 2 groups. [Conclusion]The results showed that extensive genetic variations were detected both within and among the fiveN.sibiricapopulations, whose geographic variations were in accordance with the random variation mode. These results could afford basis for protecting germplasm resources ofN.sibirica.
NitrariasibiricaPall.;natural population;fruit;leaf;phenotypic;variation
2016-06-12 基金項目: 山東省農業良種工程重大課題“林木種質資源收集保護與評價”(魯農良字[2010]6號);山東(暖溫帶)珍稀瀕危樹種種質資源保護與利用建設項目。 作者簡介: 董 昕(1987—),女,碩士,主要從事林木種質資源收集、保護與利用研究。E-mail: energy0126@sina.com * 通訊作者:解孝滿(1970—),男,碩士,研究員,主要從事林木種質資源收集、保護與利用研究。E-mail: xxm529@126.com
10.13275/j.cnki.lykxyj.2017.02.015
S793.9
A
1001-1498(2017)02-0293-07
