巨菌草纖維特性分析及其木素結(jié)構(gòu)表征
2017-04-19 12:33:50陳嘉川楊桂花劉姍姍胡長青
中國造紙 2017年3期
關(guān)鍵詞:結(jié)構(gòu)
沈 鋒 陳嘉川 楊桂花,* 王 強 劉姍姍 胡長青
(1.齊魯工業(yè)大學制漿造紙科學與技術(shù)教育部重點實驗室,山東濟南,250353;2.山東晨鳴紙業(yè)集團股份有限公司,山東壽光,262700)
·巨菌草纖維·
巨菌草纖維特性分析及其木素結(jié)構(gòu)表征
沈 鋒1陳嘉川1楊桂花1,*王 強1劉姍姍1胡長青2
(1.齊魯工業(yè)大學制漿造紙科學與技術(shù)教育部重點實驗室,山東濟南,250353;2.山東晨鳴紙業(yè)集團股份有限公司,山東壽光,262700)
對巨菌草纖維形態(tài)、化學成分組成、木素結(jié)構(gòu)及其微區(qū)分布進行了研究。利用纖維質(zhì)量分析儀(FQA)測得纖維平均長度為0.95 mm,寬度為23.1 μm,長寬比為41.1。高效液相色譜(HPLC)定量分析堿性硝基苯氧化產(chǎn)物確定巨菌草木素結(jié)構(gòu)為典型的GSH型。利用核磁共振磷譜(31P-NMR)定量分析木素各酚羥基功能基團,結(jié)果顯示脂肪族羥基含量最高,為3.96 mmol/g,其中非縮合型羥基占總酚羥基的比例較大,為96.3%;二維核磁共振(2D-HSQC NMR)波譜分析表明,β-O- 4′芳基醚鍵結(jié)構(gòu)比例最高,為84.4%;其次為β-β′樹脂醇結(jié)構(gòu)和β-5′苯基香豆?jié)M結(jié)構(gòu),所占比例分別為9.9%和5.7%;在芳香區(qū)含有大量的紫丁香基和愈創(chuàng)木基結(jié)構(gòu)單元,其中S/G比例為1.25。共聚焦拉曼光譜(CRM)分析表明,木素在細胞不同區(qū)域的濃度由大到小的順序為細胞角隅CC>復(fù)合胞間層CML>次生細胞壁SW;羥基肉桂酸的濃度分布規(guī)律與木素的分布變化相似,但其濃度低于木素的濃度。
巨菌草;木素結(jié)構(gòu);核磁共振磷譜;二維核磁共振
木質(zhì)纖維生物質(zhì)資源作為一種豐富的可再生資源,其富含的多糖組分經(jīng)過發(fā)酵可生產(chǎn)生物燃料和生物質(zhì)基化學品[1]。常見的生物質(zhì)原料主要包括木材和禾本科植物、農(nóng)林廢棄物、能源植物等,其主要組分為纖維素、半纖維素和木素[2]。在這些生物質(zhì)原料中,能源植物由于具有產(chǎn)量大且培育成本低的特點而成為潛在的能夠滿足大規(guī)模生產(chǎn)的原料,目前在制漿造紙、紡織、高附加值化工產(chǎn)品等領(lǐng)域有一定的應(yīng)用。因此,高產(chǎn)低投資的能源植物成為科技人員的研究熱點,其中就包括巨菌草(Pennisetumsinese)[3]。
巨菌草是由福建農(nóng)林大學林占熺研究員于1983年引進中國的。這種植物是由多種狼尾草屬雜交而成的單子葉多年生植物,對溫度和土壤要求不高,適應(yīng)能力強,植株高大,不易倒伏,熱利用率較高,生物量大,一年可以長高5~7 m,最高產(chǎn)量達300 t/hm2,為已知人工種植植物的最高產(chǎn)量[4-5]。目前,巨菌草在生物質(zhì)發(fā)電、纖維板、制造燃料乙醇等領(lǐng)域已有應(yīng)用。若以巨菌草為原料,研發(fā)出各種具有優(yōu)異性能、高附加值的生物質(zhì)材料,可進一步擴大巨菌草的經(jīng)濟效益和環(huán)境效益[6-7]。
本實驗主要利用高效液相色譜分析(HPLC)、傅里葉紅外光譜(FT-IR)、核磁共振磷譜(31P-NMR)、二維核磁共振(2D-HSQC NMR)、凝膠滲透色譜(GPC)和共聚焦拉曼光譜(CRM)技術(shù)分析探討巨菌草的化學組成、木素的基本結(jié)構(gòu)及其分布,為巨菌草全組分的高值化利用提供理論指導(dǎo)。
1 材料與方法
1.1 實驗原料
巨菌草,一年生,取自廣東省。巨菌草除去葉、根,然后粉碎,取40~60目木粉使用。
1.2 實驗方法
1.2.1 巨菌草纖維形態(tài)測定及化學成分分析
采用加拿大OPTEST公司的纖維質(zhì)量分析儀(FQA)分析纖維長度、寬度等形態(tài)學參數(shù)。
纖維素、半纖維素、木素、苯-醇抽出物、灰分等按照國家標準(GB/T 2677.10—1995,GB/T 2677.9—1994,GB2677.8—81,GB2677.7—81, GB2677.3—81)進行測定。
1.2.2 木粉的堿性硝基苯氧化
實驗具體方法參見謝益民教授[8]的改良方法,對降解產(chǎn)物的分析采用高效液相色譜分析(HPLC)來定性定量。用10 mL二氧六環(huán)溶解經(jīng)過減壓蒸發(fā)除去溶劑的降解產(chǎn)物,再取其1 mL稀釋5倍后用HPLC進行定性定量分析。精確稱量紫丁香醛(S)、紫丁香酸(S)、香草醛(G)、香草酸(G)、對羥基苯甲醛(H)、對羥基苯甲酸(H)作為準樣,定量分析巨菌草中木素結(jié)構(gòu)單元的含量。
HPLC分析條件為:色譜柱Nova-PakC18;流動相為乙腈∶水=12∶88(體積比),并加0.1%的磷酸;流速為1.2 mL/min;進樣量為20 μL;檢測器為紫外吸收檢測器,280 nm。
1.2.3 木素的分離提取
取40~60目巨菌草木粉用丙酮抽提8 h,抽提結(jié)束后用去離子水洗滌。風干后的試樣用球磨機球磨72 h后供制備酶解-溫和酸解木素(EMAL)用。按照文獻[9-10]進行EMAL的分離制備,測量純化后木素樣品的純度和糖含量等。
1.2.4 FT-IR分析
取1.0 mg巨菌草EMAL試樣,與300 mg KBr經(jīng)瑪瑙研缽研磨后壓片,采用美國ANALECT公司的PFX- 65紅外光譜儀進行掃描,掃描范圍為4000~400 cm-1。
1.2.531P-NMR分析
稱取干燥的巨菌草EMAL樣品40 mg(精確至0.0001 g),溶解于0.5 mL氘代吡啶-氘代氯仿(體積比為1.6∶1)溶劑中,使木素完全溶解,加入200 μL 濃度為80 mg/mL內(nèi)標(內(nèi)標劑:環(huán)己醇)溶液和50 μL濃度為11 mg/mL松弛劑(即馳豫劑:乙酰基丙酮絡(luò)酸鉻)溶液,充分混合均勻,再加入100 μL磷化試劑(2-氯- 4,4,5,5-四甲基-1,3,2-二氧磷雜戊環(huán),純度是95.0%,TMDP),迅速將混合溶液轉(zhuǎn)移至測試管中進行檢測[11]。
磷化后的EMAL樣品用核磁共振波譜儀(AVANCEⅡ400)測定木素中的基團含量。掃描時間為1 h左右。
1.2.6 2D-HSQC NMR分析
將20 mg巨菌草EMAL樣品溶于DMSO-d6有機溶劑中,然后在核磁共振波譜儀上測定,具體條件為:弛豫時間1.5 s,采樣時間0.17 s,采樣128次,256增量,即128×256[12]。
1.2.7 巨菌草EMAL分子質(zhì)量分布
巨菌草EMAL的乙酰化參照文獻方法[12]進行。取100 mg巨菌草EMAL樣品溶于5 mL體積比為1∶2的吡啶-醋酸酐溶液中,避光條件下充入氮氣保護,攪拌24 h后停止反應(yīng)。然后將溶液緩慢倒入攪拌著的200 mL乙醚中,析出乙酰化的木素樣品,用乙醚洗滌數(shù)次直至無吡啶氣味,真空干燥6 h,得到的粉末即為乙酰化的木素樣品。乙酰化的木素樣品通過凝膠滲透色譜(GPC)來測定木素質(zhì)均相對分子質(zhì)量(Mw)和數(shù)均相對分子質(zhì)量(Mn)[12]。
1.2.8 共聚焦拉曼光譜(CRM)分析
選取合適位置的巨菌草試樣用單面刀片分割成1 cm 長小段,然后經(jīng)煮沸軟化后用聚乙二醇包埋劑進行包埋,然后使用滑走切片機(Leica SM2010R,Germany)切取厚度為8 μm的橫切面切片,將切好的樣品置于載玻片上,滴加1滴氘水,然后蓋上蓋玻片。采用共聚焦拉曼顯微鏡(Horiba JobinYvon, Longjumeau, France)進行檢測。具體方法見參考文獻[13]。
2 結(jié)果與討論
2.1 巨菌草的纖維形態(tài)
表1為巨菌草的纖維形態(tài)數(shù)據(jù)。由表1可知,巨菌草纖維的平均長度為0.95 mm,平均寬度為23.1 μm,長寬比為41.1。纖維長度略低于一般禾本科原料(麥草平均長度1.32 mm,蘆葦平均長度1.12 mm),長寬比數(shù)值較大,有較好的纖維交織能力,可作為制漿造紙纖維原料使用。
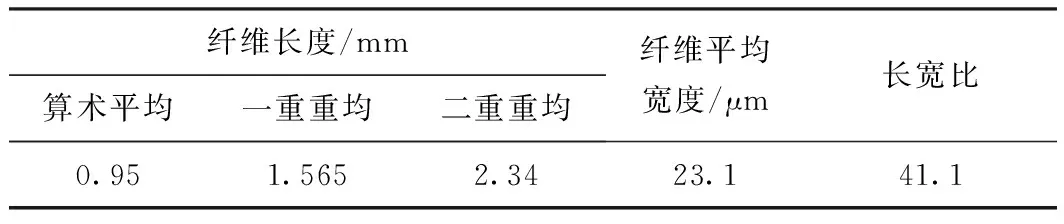
表1 巨菌草纖維形態(tài)
2.2 化學成分分析
2.2.1 巨菌草的化學組分分析
根據(jù)國家相關(guān)標準,對巨菌草的化學成分進行了分析測定,分析結(jié)果見表2。
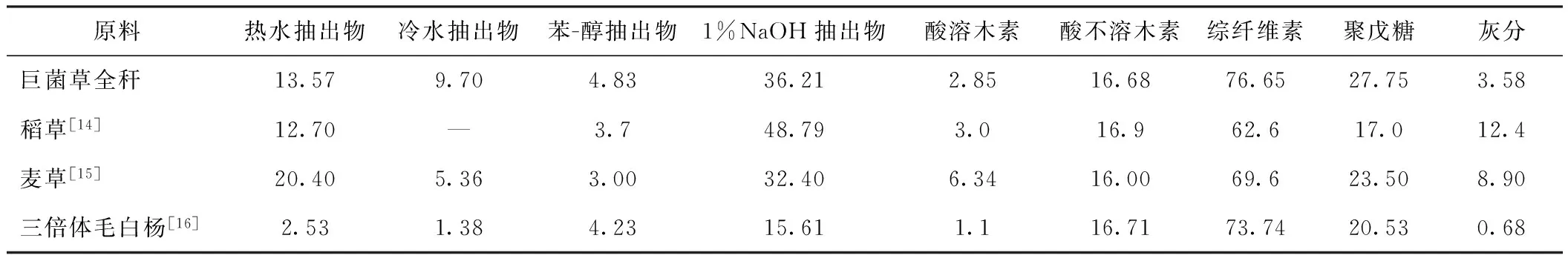
表2 巨菌草的化學成分 /%
從表2中數(shù)據(jù)可以看出,巨菌草的灰分含量小于稻草和麥草的,熱水抽出物和苯-醇抽出物含量高于三倍體毛白楊[14-16]的。巨菌草的綜纖維素含量為76.65%,木素含量為19.53%,聚戊糖含量為27.75%。基于纖維素含量、木素含量、灰分含量等數(shù)據(jù)分析,巨菌草易于蒸煮,化學藥品消耗量低,制漿得率較高;其制漿造紙性能優(yōu)于麥草,稍差于三倍體毛白楊,是較好的制漿造紙用非木材纖維原料[5, 17]。
2.2.2 巨菌草EMAL的成分分析
巨菌草EMAL成分分析結(jié)果見表3。表3中數(shù)據(jù)表明,通過酶解-溫和酸解法制備的EMAL純度為92.5%。木素中含有約7.5%的碳水化合物,且碳水化合物中主要是木糖,這說明木素與部分聚木糖存在部分交聯(lián),且在酶解-溫和酸解法制備過程中并未完全去除。
2.3 硝基苯氧化
堿性硝基苯氧化巨菌草原料中木素的非酚型結(jié)構(gòu)時,主要生成紫丁香醛/酸(S)、香草醛/酸(G)、對羥基苯甲醛/酸(H)這3種小分子醛/酸,而其對應(yīng)的木素基本結(jié)構(gòu)單元為紫丁香基丙烷(S)、愈創(chuàng)木基丙烷(G)和對羥基苯基丙烷(H)結(jié)構(gòu)。因此,通過測定硝基苯氧化產(chǎn)物中幾種酚醛和酚酸的含量,就可推斷出木素非縮合型結(jié)構(gòu)單元的比例。巨菌草所含EMAL結(jié)構(gòu)單元的含量見表4。

表3 巨菌草EMAL的成分分析 /%

表4 巨菌草木素結(jié)構(gòu)單元的含量
注 *S代表紫丁香醛與紫丁香酸之和;G代表香草醛與香草酸之和。
由表4中數(shù)據(jù)可知看出,木素經(jīng)過硝基苯氧化之后,酚醛/酸降解產(chǎn)物得率較低,為40.8%。巨菌草木素結(jié)構(gòu)單元中紫丁香基丙烷結(jié)構(gòu)單元(S)含量最高,為38.34%;愈創(chuàng)木基丙烷結(jié)構(gòu)(G)單元次之,為27.42%;對羥基苯基丙烷結(jié)構(gòu)(H)單元最少,為8.5%;而且降解產(chǎn)物中還含有較多的阿魏酸(FA)和對香豆酸(PCA)等小分子產(chǎn)物。其中S/G的比值為1.40,由于紫丁香基丙烷結(jié)構(gòu)不易形成縮合型結(jié)構(gòu),因此S/G的比值與木素中非縮合性結(jié)構(gòu)的比例有很大的關(guān)系,而且巨菌草中木素結(jié)構(gòu)是典型的GSH型,其比例為S∶G∶H=1.4∶1.0∶0.4。此外,作為木素和半纖維素結(jié)合的交聯(lián)聚合物,阿魏酸和對香豆酸的含量較高,且具有草類原料禾本科植物的特征[18]。非縮合型結(jié)構(gòu)木素在制漿或漂白過程中容易降解,因而紫丁香基丙烷結(jié)構(gòu)含量越多的木素越容易降解。由此可知,巨菌草的木素結(jié)構(gòu)比較容易降解。
2.4 巨菌草EMAL的FT-IR分析


圖1 巨菌草EMAL的FT-IR圖
2.5 巨菌草EMAL的31P-NMR譜分析
經(jīng)過衍生化的巨菌草EMAL樣品,通過核磁共振磷譜儀測定得到的31P-NMR譜圖見圖2。

圖2 巨菌草EMAL的31P-NMR譜圖
圖2表明,巨菌草EMAL樣品中,主要為酚羥基、脂肪族羥基、羧基等,其中酚羥基又包括縮合型和非縮合型愈創(chuàng)木基型(G)、紫丁香基型(S)和對羥基苯基型(H)羥基,這些羥基都是木素結(jié)構(gòu)的功能基團。對這些基團的信號區(qū)域進行積分,可以得到積分值,然后再根據(jù)內(nèi)標用量,計算出木素中各官能團的含量,單位為mmol/g。
對于不同原料,其積分區(qū)間也不相同。巨菌草EMAL中紫丁香基酚羥基出現(xiàn)在δ=143.1~142.4范圍之內(nèi),它和縮合酚羥基出現(xiàn)的范圍δ=144.2~141.5相重疊,因此在積分的時候,需要把紫丁香基從縮合酚羥基中分離出來(見圖2),從而使縮合酚羥基的積分區(qū)域分為δ=144.2~143.1和δ=142.4~141.5兩個積分區(qū)間。

表5 31P-NMR定量分析EMAL的功能基團及其含量
由表5中數(shù)據(jù)可知,巨菌草EMAL中脂肪族羥基含量最高,為 3.96 mmol/g,總酚羥基含量次之,為1.62 mmol/g,羧基含量最低,僅為0.05 mmol/g。在酚羥基中,非縮合型酚羥基含量較高,為1.56 mmol/g,縮合型酚羥基含量較低,為0.06 mmol/g。各非縮合型酚羥基占總酚羥基的含量分別為:S-酚羥基10.5%,G-酚羥基22.2%,H-酚羥基63.6%,而縮合型酚羥基(S—OH和G—OH)3.7%。需要說明的是,圖2中顯示的紫丁香基酚羥基的數(shù)量并不能真實反映木素結(jié)構(gòu)中紫丁香基單元的實際數(shù)量,因為木素分子質(zhì)量較大,結(jié)構(gòu)復(fù)雜,連接鍵較多且S-木素容易形成β-O- 4等芳基醚鍵,因而游離出較少的S-酚羥基,從而導(dǎo)致總酚羥基和各酚羥基含量較低[20]。
2.6 巨菌草EMAL的2D-HSQC NMR譜分析
為了進一步研究巨菌草EMAL結(jié)構(gòu)單元之間的連接鍵結(jié)構(gòu),以及各連接鍵的比例,使用2D-HSQC NMR技術(shù)進行了表征,見圖3。圖3(a)表示木素側(cè)鏈區(qū)域(δC/δH50~90/2.9~5.1),圖3(b)代表芳環(huán)區(qū)域(δC/δH98~135/5.2-7.6),圖4為木素的主要連接結(jié)構(gòu)及單元,表6為2D-HSQC 譜圖信號的化學位移歸屬[3, 21]。
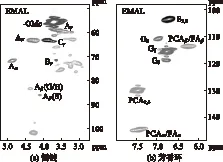
圖3 巨菌草EMAL側(cè)鏈和芳香環(huán)上C—H的 2D-HSQC NMR譜圖
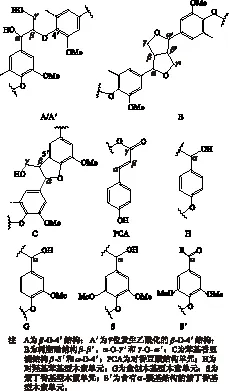
圖4 巨菌草EMAL的主要結(jié)構(gòu)
由圖3和圖4可知,巨菌草EMAL結(jié)構(gòu)之間的連接鍵主要為β-O- 4′芳基醚鍵結(jié)構(gòu)(A 和 A′),β-O- 4′結(jié)構(gòu)(A)上Cα—Hα、Cβ—Hβ、Cγ—Hγ的化學位移δC/δH分別為72.0/4.89,83.5/4.45和84.9/4.12,60.7/3.53,而γ位乙酰化的β-O- 4′結(jié)構(gòu)(A′)的Cγ—Hγ的相關(guān)信號δC/δH主要集中在63.0/4.29處,證明巨菌草EMAL的β-O- 4′結(jié)構(gòu)在γ位發(fā)生了一定的乙酰化反應(yīng),乙酰化廣泛存在于植物組織中[22]。在側(cè)鏈區(qū)域還檢測到木素其他縮合型結(jié)構(gòu),如樹脂醇結(jié)構(gòu)(resinol,β-β′,B),苯基香豆?jié)M結(jié)構(gòu)(phenylcoumaran,β-5′,C)等,在Cγ—Hγ的化學位移δC/δH分別為71.6/3.69和63.5/3.78。

表6 巨菌草EMAL 2D-HSQC NMR譜圖化學位移及歸屬
在木素的芳香環(huán)區(qū)域,可以明顯觀察到木素的3種結(jié)構(gòu)紫丁香基(S)、愈創(chuàng)木基(G)和對羥苯基(H)單元吸收信號,這說明巨菌草是典型的GSH型單元木素。S型單元的2,6位碳是相互對稱的,其C2,6—H2,6的化學位移δC/δH為103.6/6.69,S型單元α位發(fā)生了一定的氧化反應(yīng),S′(α-oxidation)的化學位移δC/δH為106.3/7.19。G型單元的信號峰出現(xiàn)在不同的位置:C2—H2(G2)、C5—H5(G5)、C6—H6(G6)的化學位移δC/δH分別為111.3/6.96,115.4/6.75和114.0/6.67,118.9/6.77。H型單元得吸收峰與G型單元的吸收峰比較接近,其中H3,5的化學位移主要與G型單元5號位上的相關(guān)信號發(fā)生重疊。對香豆酸的吸收信號較強,在不同位置均有吸收信號。其中C2,6—H2,6位的化學位移δC/δH為115.4/6.77,Cα—Hα和Cβ—Hβ位的化學位移δC/δH分別為144.4/7.39和114.3/6.25。有研究發(fā)現(xiàn)木素中的H單元主要以對香豆乙酰基的形式存在[23]。
利用2D-HSQC NMR圖半定量技術(shù)分析巨菌草EMAL中不同結(jié)構(gòu)單元及S/G比值,分析結(jié)果見表7。表7結(jié)果表明,巨菌草EMAL主要結(jié)構(gòu)單元為β-O- 4′芳基醚鍵(A/A′),其占總側(cè)鏈單元的84.4%,其次為β-β′樹脂醇結(jié)構(gòu)(B),占總側(cè)鏈單元的9.9%,而β-5′苯基香豆?jié)M結(jié)構(gòu)(C)占總側(cè)鏈單元的5.7%。二維圖譜得到的S/G比值為1.25,與硝基苯氧化中S/G為1.40相比略低,可能原因是硝基苯氧化主要與木素非縮合型單元發(fā)生反應(yīng),而且2D-HSQC NMR譜圖中是木素整體結(jié)構(gòu)單元的比例。

表7 巨菌草EMAL結(jié)構(gòu)2D-HSQC NMR譜圖定量分析

圖6 巨菌草細胞壁組分分布的CRM成像圖
2.6 巨菌草EMAL分子質(zhì)量分布
巨菌草EMAL的質(zhì)均相對分子質(zhì)量(Mw)和數(shù)均相對分子質(zhì)量(Mn)通過凝膠滲透色譜(GPC)來測定,以苯乙烯標樣作為標準,測定譜圖見圖5。由圖5可知,巨菌草EMAL的質(zhì)均相對分子質(zhì)量為7289,的多分散系數(shù)較小(小于2.0),這說明EMAL的分子質(zhì)量分布較小。

圖5 巨菌草的相對分子質(zhì)量分布
2.7 巨菌草木素共聚焦拉曼光譜(CRM)分析
圖6為巨菌草纖維細胞中木素、碳水化合物和羥基肉桂酸的分布圖像。由圖6(a)可以清晰地觀察到木素在細胞角隅(CC)的拉曼信號強度最強,這表明CC中木素濃度最高,其次是復(fù)合胞間層(CML),次生細胞壁(SW)木素信號強度最弱,說明木素的濃度在SW中最低。與木素在不同細胞層次中的分布相類似的是羥基肉桂酸的分布,羥基肉桂酸(HCA)主要由PCA 和FA組成,它們作為木素與半纖維素結(jié)合的交聯(lián)聚合物分子,其在細胞壁中的分布在植物細胞壁天然抗降解屏障中扮演著重要角色,因而了解其分布對于后續(xù)巨菌草的資源化利用以及生物質(zhì)預(yù)處理等工藝可提供理論指導(dǎo)。圖6(b)主要顯示碳水化合物在纖維細胞中的分布,其主要包括纖維素和半纖維素。從圖6中可以看出碳水化合物在SW中拉曼信號強度最強,表明碳水化合物在SW中濃度最高,而在CC和CML區(qū)域碳水化合物的信號強度較弱,其濃度也較低。
3 結(jié) 論
(1)巨菌草纖維長度較長,為0.95 mm,長寬比較大,為41.1,而且綜纖維素含量較高,木素含量較低,是較好的制漿造紙原料。
(2)通過硝基苯氧化可知,巨菌草酶解-溫和酸解木素(EMAL)中紫丁香基型(S)和愈創(chuàng)木基型(G)結(jié)構(gòu)單元含量較高,分別為38.3%和27.4%,且S/G的比例為1.40,是典型的GSH型禾本科植物。核磁共振磷譜(31P-NMR)圖譜分析表明巨菌草EMAL中含有脂肪族羥基、縮合型和非縮合型的愈創(chuàng)木基型、紫丁香基型和對羥基苯基型羥基和羧基,其中脂肪族羥基含量最高,為 3.96 mmol/g,酚羥基中非縮合型酚羥基含量較高,為1.56 mmol/g,縮合型酚羥基含量較低,為0.06 mmol/g,羧基含量最低,僅為0.05 mmol/g。
(3)二維核磁共振(2D-HSQC NMR)分析技術(shù)表明,巨菌草EMAL主要包含β-O- 4′芳基醚鍵、β-β′樹脂醇結(jié)構(gòu)、β-5′苯基香豆?jié)M結(jié)構(gòu),在芳香區(qū)含有大量的紫丁香基和愈創(chuàng)木基結(jié)構(gòu)單元,以及大量的對香豆酸結(jié)構(gòu)單元,從而構(gòu)成了木素和半纖維素結(jié)合的交聯(lián)聚合物分子,并在植物細胞壁的抗降解屏障中發(fā)揮重要作用。共聚焦拉曼光譜(CRM)表明巨菌草木素在細胞不同區(qū)域的濃度分布不均一,其中細胞角隅(CC)中木素濃度最高,復(fù)合胞間層(CML)濃度次之,次生細胞壁(SW)中木素濃度最低。羥基肉桂酸的濃度分布規(guī)律與木素的分布變化相似,但其濃度低于木素的濃度,碳水化合物主要分布在次生細胞壁中。
[1] Zhao X, Zhang L, Liu D. Biomass recalcitrance. Part II: Fundamentals of different pre-treatments to increase the enzymatic digestibility of lignocellulose[J]. Biofuels, Bioproducts and Biorefining, 2012, 6(5): 561.
[2] You T, Zhang L, Guo S, et al. Unraveling the structural modifications in lignin ofArundodonaxLinn. during acid-enhanced ionic liquid pretreatment [J]. Journal of Agricultural & Food Chemistry, 2015, 63(50): 10747.
[3] Río JC Del, Prinsen P, Jorge R, et al. Structural characterization of the lignin in the cortex and pith of elephant grass (Pennisetumpurpureum) stems[J]. Journal of Agricultural & Food Chemistry, 2012, 60(14): 3619.
[4] Lin Xing-sheng, Lin Zhan-xi, Lin Dong-mei,et al. Preliminary Study on Juncao as Biomass Fuel[J]. Journal of Fujian College of Forestry, 2013, 33(1)1: 82. 林興生, 林占熺,林冬梅, 等. 菌草作為生物質(zhì)燃料的初步研究[J]. 福建林學院學報, 2013, (1): 82.
[5] Lu Nan, Shen Feng, Wang Sheng-dan, et al. Biological Structure and KP Pulp Properties of Pennisetum Sinese Grasses[J]. Paper and Paper Making, 2015, 34(3): 27. 魯 南, 沈 鋒, 王勝丹, 等. 巨菌草的生物結(jié)構(gòu)及制漿造紙性能 [J]. 紙和造紙, 2015, 34(3): 27.
[6] Lin Zhan-xi. Juncao Technology Status and Application Prospect[J]. Academic Review. 1996(z1): 80. 林占禧. 菌草技術(shù)現(xiàn)狀及其應(yīng)用前景[J]. 學術(shù)評論, 1996(z1): 80.
[7] Li Zhi-wen. Study on Characteristics of Pennisetum Sinese as an Energy Source [J]. Agriculture Engineering Technology(New Energy Industry), 2013(6): 12. 李志文. 巨菌草作為能源草的特性研究[J]. 農(nóng)業(yè)工程技術(shù)·新能源產(chǎn)業(yè), 2013(6): 12.
[8] Xie Y, Robert D R, Terashima N. Selective carbon 13 enrichment of side chain carbons of ginkgo lignin traced by carbon 13 nuclear magnetic resonance[J]. Plant physiology and biochemistry, 1994, 32(2): 243.
[9] LU Nan,SHEN Feng,WANG Sheng-dan, et al. Studies on Lignin and Hemicellulose of Pennisetum Sinese Grasses [J]. China Pulp & Paper, 2016, 35(8):1. 魯 南, 沈 鋒, 王勝丹, 等. 巨菌草的木素及半纖維素分析 [J]. 中國造紙, 2016, 35(8): 1.
[10] Liu Yu, Zhan Huai-yu, Chen Jia-chuan, et al. Structural analysis of raw material lignin from triploid of polulus tomentosa[J]. China Paper & Paper Industry, 2006, 27(11): 88. 劉 玉, 詹懷宇, 陳嘉川, 等. 三倍體毛白楊原料木素結(jié)構(gòu)分析 [J]. 中華紙業(yè), 2006, 27(11): 88.
[11] Argyropoulos D.31P NMR in wood chemistry: A review of recent progress [J]. Research on chemical intermediates, 1995, 21(3/5): 373.
[12] Wen J-L, Sun S-L, Yuan T-Q, et al. Structural elucidation of whole lignin from Eucalyptus based on preswelling and enzymatic hydrolysis[J]. Green Chemistry, 2015, 17(3): 1589.
[13] Zhe J, Xun Z, Zhe L, et al. Visualization of Miscanthus×giganteus cell wall deconstruction subjected to dilute acid pretreatment for enhanced enzymatic digestibility[J]. Biotechnology for Biofuels, 2015, 8(1): 1.
[14] Wang Wang-xia, Jing Lei, Jin Yong-can, et al. Effects of Acetic Acid Pre-hydrolysis on the Chemical Compositions and Enzymatic Hydrolysis of Rice Straw[J]. Jounrnal of Cellulose Science and Technology. 2011, 19(2): 35. 王旺霞, 荊 磊, 金永燦, 等. 醋酸預(yù)水解對稻草化學成分及酶水解性能的影響[J]. 纖維素科學與技術(shù), 2011, 19(2): 35.
[15] Li You-sen, Yang Shu-hui, Wu Jie-sheng. Study on the Production of Linearboard with Ammonium Sulfite Kenaf and Wheat Straw Pulps[J]. Journal of Northwest Institute of Light Industry, 1990, 8(3): 73. 李友森, 楊淑惠, 吳解生. 亞銨法紅麻、 麥草漿配抄掛面牛皮箱紙板試驗研究 [J]. 西北輕工業(yè)學院學報, 1990, 8(3): 73.
[16] Yao Chun-li, Pu Jun-wen. Timber Characteristics and Pulp Properties of the Triploid ofPopulustomentosa[J]. Journal of Beijing Forestry University, 1998, 20(5): 18. 姚春麗, 蒲俊文. 三倍體毛白楊化學組分纖維形態(tài)及制漿性能的研究[J]. 北京林業(yè)大學學報, 1998, 20(5): 18.
[17] Pu Jun-wen, Song Jun-long, Xie Yi-min, et al. Characteristics of lignin structure of triploid clones of Populus tomentosa Carr[J]. Journal of Beijing Forestry University, 2002, 24(56): 211. 蒲俊文, 宋君龍, 謝益民,等. 三倍體毛白楊木質(zhì)素結(jié)構(gòu)特性研究[J]. 北京林業(yè)大學學報, 2002, 24(56): 211.
[18] Wu Qin, Chen Jia-chuan, Yang Gui-hua, et al. Fiber morphology, chemical composition, and APMP pulp properties of fast growing black poplar crotches[J]. Chemistry and Industry of Forest Products, 2010, 30(3): 35. 吳 芹, 陳嘉川, 楊桂花, 等. 速生黑楊枝椏材纖維形態(tài)、化學組分及APMP制漿性能的研究[J]. 林產(chǎn)化學與工業(yè), 2010, 30(3): 35.
[19] Faix O. Classification of lignins from different botanical origins by FT-IR spectroscopy[J]. Holzforschung-International Journal of the Biology, Chemistry, Physics and Technology of Wood, 1991, 45(s1): 21.
[20] Li Bing-yun, Zhan Huai-yu, Liang Chen, et al. Analysis of lignin structure in Bambusa Chungii SCMP by 2D NMR[J]. Paper Science & Technology, 2010, 29(5): 62. 李兵云, 詹懷宇, 梁 辰, 等. 粉丹竹SCMP木素結(jié)構(gòu)的2D NMR分析[J]. 造紙科學與技術(shù), 2010, 29(5): 62.
[21] Jorge R, Pepijn P, Ana G, et al. Isolation and structural characterization of the milled wood lignin, dioxane lignin, and cellulolytic lignin preparations from brewer′s spent grain[J]. Journal of Agricultural & Food Chemistry, 2014, 63(2): 603.
[22] Río J C Del, Gisela M, Jorge R, et al. Occurrence of naturally acetylated lignin units[J]. Journal of Agricultural & Food Chemistry, 2007, 55(14): 5461.

(責任編輯:董鳳霞)
Fiber Morphology, Lignin Structure and Lignin Distribution in Cell Wall of thePennisetumsinese
SHEN Feng1CHEN Jia-chuan1YANG Gui-hua1,*WANG Qiang1LIU Shan-shan1HU Chang-qing2
(1.KeyLabofPulp&PaperScienceandTechnologyofEducationMinistry,QiluUniversityofTechnology,Ji’nan,ShandongProvince, 250353; 2.ChenmingPulp&PaperCo.,Ltd.,Shouguang,ShandongProvince, 262700)
(*E-mail: ygh2626@126.com)
The morphologiy of fibers, chemical components, lignin structure and the lignin distribution in cell wall ofPennisetumsinesewere analyzed. Fiber Quality Analyzer (FQA) analysis showed that average fiber length was 0.96mm and the width was 23.1 μm, fiber length-width ratio reached to 41.1. Alkaline nitrobenzene oxidation quantitative analysis revealed thatPennisetumsineselignin was typical GSH-type. It was found that the highest content of hydroxyl groups was aliphatic OH (3.96 mmol/g), and non-condensed phenolic hydroxyl accounted for a large proportion content of the total phenolic hydroxyl (96.3%). In addition, 2D-HSQC NMR analysis indicated that the alkyl aryl ether linkages was predominant (β-O- 4′, 84.4% of total linkages), followed by resinols (β-β′, 9.9%) and phenylcomumarans (β-5′5.7%). Moreover, aromatic regions showed large quantity of S-type and G-type lignin, and the S/G ratio was 1.25. Confocal Raman microscopy (CRM) indicated that the concentrations of lignin and hydroxycinnamic acid (HCA) in the cell walls followed the decreasing order, as CC>CML>SW, but the concentration of HCA waslowerthan that of lignin.
Pennisetumsinese; lignin structure;31P-NMR; 2D-HSQC NMR
沈 鋒先生,在讀碩士研究生;主要從事植物纖維結(jié)構(gòu)方面的研究。
2016- 12- 05(修改稿)
國家自然科學基金(31370580, 31470602);山東省重大科技專項(2015ZDZX09002);泰山學者工程專項經(jīng)費。
TS721
A
10.11980/j.issn.0254- 508X.2017.03.003
*通信作者:楊桂花,博士生導(dǎo)師;研究領(lǐng)域:制漿造紙與生物質(zhì)精煉綠色化學技術(shù)。
猜你喜歡
小獼猴智力畫刊(2023年4期)2023-04-23 08:49:58
哲學評論(2021年2期)2021-08-22 01:53:34
中華詩詞(2019年7期)2019-11-25 01:43:04
模具制造(2019年3期)2019-06-06 02:10:54
中學生數(shù)理化·高一版(2018年1期)2018-02-10 05:20:03
影視與戲劇評論(2016年0期)2016-11-23 05:26:01
七彩語文·寫字與書法(2016年7期)2016-07-28 21:40:22
七彩語文·寫字與書法(2016年6期)2016-07-15 19:36:34
人間(2015年21期)2015-03-11 15:23:21
現(xiàn)代企業(yè)(2015年9期)2015-02-28 18:56:50
