有氧運動訓練和攝食對中華倒刺鲃幼魚力竭運動后代謝特征的影響
2017-04-12 09:49:15李秀明袁建明張耀光付世建
水生生物學報 2017年2期
李秀明袁建明,張耀光付世建
(1. 重慶師范大學進化生理與行為學實驗室, 重慶市動物生物學重點實驗室, 重慶 401331; 2. 西南大學淡水魚類資源與生殖發育教育部重點實驗室, 重慶 400715; 3. 重慶市水產技術推廣站, 重慶 400020)
有氧運動訓練和攝食對中華倒刺鲃幼魚力竭運動后代謝特征的影響
李秀明1,2袁建明2,3張耀光2付世建1
(1. 重慶師范大學進化生理與行為學實驗室, 重慶市動物生物學重點實驗室, 重慶 401331; 2. 西南大學淡水魚類資源與生殖發育教育部重點實驗室, 重慶 400715; 3. 重慶市水產技術推廣站, 重慶 400020)
為了探討有氧運動訓練和攝食對中華倒刺鲃(Spinibarbus sinensis)幼魚力竭運動后代謝特征的影響, 在(25±0.5)℃條件下, 將120尾實驗魚[體重(21.35±0.05) g, 體長(10.21±0.03) cm]隨機分成4組, 即: 對照組、1、2和4 BL/s (體長/秒, body length/s)訓練組, 分別放置于不同流速下處理8周。隨后測定各實驗組心臟和鰓指數以及禁食或攝食(輕度麻醉灌喂體重1.5%的餌料)狀態下的力竭運動后過量耗氧。結果發現: 4 BL/s訓練組的心臟和鰓指數都顯著高于其他實驗組(P<0.05); 無論攝食與否, 3個訓練組運動前代謝率都顯著高于對照組(P=0.001), 而各實驗組過量耗氧均沒有顯著差異; 在禁食狀態下, 僅4 BL/s訓練組的運動代謝峰值和代謝率增量顯著高于對照組, 而在攝食狀態下, 3個訓練組的運動代謝峰值和代謝率增量均顯著高于對照組(P<0.005)。與禁食組相比, 攝食導致各處理組的運動前代謝率顯著上升(P<0.001), 但對運動代謝峰值沒有顯著影響; 另外, 攝食對照組代謝率增量和力竭運動后過量耗氧顯著低于禁食對照組(P<0.05)。研究表明: (1)有氧運動訓練顯著提高了中華倒刺鲃幼魚的有氧代謝能力, 這可能與其呼吸和循環系統功能的改善有關; (2)力竭運動能夠誘導出中華倒刺鲃幼魚的最大有氧代謝率; (3)攝食削弱了中華倒刺鲃幼魚無氧代謝能力。
有氧運動訓練; 攝食; 運動后過量耗氧; 代謝率; 中華倒刺鲃
由于魚類具有逆流游泳的習性, 因此是運動訓練研究的理想對象[1]。根據對氧氣的需求與否, 魚類的運動訓練可以分為持續的有氧運動訓練和間歇性的無氧運動訓練兩種類型[2]。魚類在劇烈運動期間通過增加耗氧率來滿足其能量需求, 為了償還“氧債”其運動后耗氧率往往顯著高于運動前水平,這種運動后恢復期間超過運動前耗氧水平的耗氧量稱之為運動后過量耗氧(Excess post-exercise oxygen consumption, EPOC)[3,4]。力竭運動后的EPOC大小(EPOC magnitude)是魚類暴發游泳和無氧代謝能力的重要評價指標, 而EPOC峰值能夠反映某些魚類的最大有氧代謝能力[5,6], 這些能力與其捕食和避敵等生命活動密切相關。因此, 魚類EPOC特征一直受到廣泛的關注[3,4,7,8]。研究發現,魚類EPOC特征不僅與其種類相關, 而且還受到各種生態因素的顯著影響[9,10]。由于受到季節交替和人類活動干預等因素的影響, 魚類經常會面臨水流速度變動的環境, 進而形成各種相應的生理生態適應策略。攝食和運動都是魚類重要的生理活動, 由于呼吸和循環系統功能的限制, 當兩者同時進行時可能會出現對氧氣需求的競爭[11,12]。目前, 有關水流變動和攝食對魚類有氧運動能力及其代謝特征的影響有較多報道[11—13], 但對其無氧運動代謝能力影響的研究報道相對較少[9]。
中華倒刺鲃(Spinibarbus sinensis)屬于鯉形目,鯉科, 是一種長江中上游重要的經濟魚類, 喜歡生活在江河底層的激流環境中[14]。目前其生長、運動和繁殖等方面的研究已有大量報道[13,15,16], 但其EPOC及其影響因素的研究還未見報道。本研究以中華倒刺鲃幼魚為實驗對象, 探討了不同流速下有氧運動訓練和攝食消化過程對其EPOC的影響, 揭示其運動生理適應策略以及攝食和運動的代謝競爭模式, 為其資源保護和生產實踐提供必要的基礎數據。
1 材料與方法
1.1 實驗材料
實驗對象中華倒刺鲃幼魚購買于重慶市合川魚種場。在實驗室室內水泥池(2000 L)中暫養4周,用浮性顆粒飼料[通威公司, 飼料粗脂肪、粗蛋白質、碳水化合物和灰分比例分別為: (8.5±0.5)%、(41.2±0.9)%、(25.7±1.2)%和(12.3±0.4)%]每天飽食投喂兩次(09:00和18:00), 每天用曝氣24h后的自來水更換10%左右的養殖用水, 養殖水體溫度控制在(25±0.5)℃左右, 養殖水體溶氧控制在接近飽和(8 mg/L左右), 采用12L∶12D光周期制度。
1.2 實驗方案
在暫養期結束后, 隨機挑選健康且大小相當的實驗魚120尾[體重(21.35±0.05) g, 體長(10.21±0.03) cm]放置于自制“魚類游泳運動訓練儀”(儀器描述詳見文獻[16])中進行不同水流速度下的有氧運動訓練處理。將MS-222(3-氨基苯甲酸乙酯甲基磺酸鹽, 50 mg/L)輕度麻醉后的實驗魚隨機分成4個實驗組,即: 對照組、1、2和4 BL/s (體長/秒, body length/s)訓練組, 并分別放置于微水流 (約3 cm/s左右)、1 BL/s (約10 cm/s)、2 BL/s (約20 cm/s)和4 BL/s (約40 cm/s)流速的水道(每個水道30尾實驗魚)中。有氧運動訓練處理歷時8周。每隔2周測定一次魚體體長, 并據此調整一次水流速度。其他實驗條件如投喂制度、水體溶氧和水溫等與暫養期間相同。
在訓練周期結束后, 從每個實驗組中隨機挑選10尾實驗魚進行心臟指數和鰓指數的測定。另外再從各實驗組中隨機挑選16尾實驗魚(其中8尾作禁食處理, 另外8尾作攝食處理)單獨放入魚類代謝儀的呼吸室內馴化1d。禁食的實驗魚用溶氧儀(HQ30d, HACH, USA)測定其耗氧率作為其運動前代謝率(耗氧率測定方法見后)。由于實驗魚在魚類代謝儀的呼吸管中不會主動攝食, 且我們先前的研究表明, 輕度麻醉灌喂不會對中華倒刺鲃幼魚的耗氧率測定產生顯著影響[16], 因此本實驗采用國內外魚類研究廣泛接受的麻醉灌喂方法進行攝食處理。攝食的實驗魚麻醉灌喂1.5%體重的糊狀餌料(浮性顆粒飼料和水的混合比例為1∶1.5, 麻醉灌喂操作詳見參考文獻[16, 17])。灌喂后3.5h對實驗魚進行耗氧率測定(先前研究發現此時為實驗魚攝食代謝峰值出現時間[16]), 作為其運動前代謝率。隨后對禁食或攝食處理的實驗魚進行力竭性運動并測定其運動恢復過程中耗氧率反應。EPOC測定具體操作如下[8]: 運動前代謝率測定結束后, 將單尾實驗魚放入環形水道(水體體積約120 L, 水流速度約65 cm/s)中用手進行追趕使其達到力竭狀態(通常為失去平衡或觸碰魚體時不再有應激反應, 在追趕2—3min內達到力竭狀態), 隨后立即將實驗魚放回呼吸室(0.2 L)中并計時, 在力竭運動后1、2、3、4、5、6、7、8、9、10、15、20、25、30、35、40、45、50、55和60min分別測定其耗氧率。呼吸室的流速設定為0.85 L/min左右, 據相關公式[18]計算, 呼吸室水體置換99%在1min內即可完成, 能夠滿足測定實驗魚的最大運動代謝水平的要求。
1.3 指標測量與參數計算
為了考察有氧運動訓練對中華倒刺鲃幼魚心臟和鰓指數的影響, 將實驗魚放入裝有過量麻醉劑(MS-222, 200 mg/L)的容器中麻醉致死。待稱量體重后將其心臟和鰓完整取出, 在魚用生理鹽水中漂洗干凈后用吸水紙將其水分吸干, 用微量天平稱量(精確至0.0001 g)。心臟和鰓指數計算公式如下[7]:
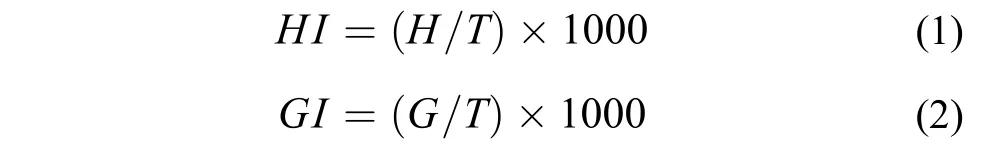
公式中HI表示心臟指數(‰), GI表示鰓指數(‰), H表示心臟重量(g), G表示鰓重量(g), T表示魚體重量(g)。
實驗魚耗氧率(作為其代謝率指標)的測定采用自主研制的魚類代謝儀[19]完成。實驗魚耗氧率計算公式如下:

公式中Rm表示單尾魚耗氧率(mg O2/h),ΔO2表示裝有實驗魚呼吸室和沒有實驗魚呼吸室(對照呼吸室)之間的溶氧差值(mg O2/L, 溶氧值用溶氧儀測量呼吸室出水口水中溶氧得到), v表示呼吸室的流速(L/h, 由呼吸室出水口1min內出水量稱重得到)。為了消除個別魚體重差異較大造成的影響, 用0.75作為體質量系數將單尾魚耗氧率標準化為1 kg體質量[16,19]。校正后耗氧率計算公式如下:
44 Prevalence and risk factors of chronic kidney disease in high-risk population in Minhang District of Shanghai

公式中Rs表示標準體重耗氧率[mg O2/(kg·h)],Rm同(3)式, m表示魚體體質量(kg)。
相關參數定義如下: (1)運動前代謝率[mg O2/ (kg·h)]是指實驗魚力竭運動前的代謝率; (2)運動代謝峰值[mg O2/(kg·h)]是指實驗魚在力竭運動后恢復過程中觀察到的最大代謝率; (3)代謝率增量[mg O2/ (kg·h)]是指力竭運動前代謝率和力竭運動后代謝峰值之間的差值; (4)過量氧耗(mg O2/kg)是指實驗魚在力竭運動后恢復過程中各個時間點高于運動前代謝率的差值在時間上的積分。
1.4 數據處理和統計分析
所有實驗數據用EXCELL (2007)進行常規計算, 用平均值±標準誤(Mean±SE)表達, 顯著性水平為P<0.05, 統計分析采用SPSS 17.0軟件進行。有氧運動訓練和攝食對EPOC參數的影響采用雙因素方差分析(ANOVA)。有氧運動訓練對所有參數的影響采用單因素方差分析, 若差異顯著則進行多重比較(LSD法)。攝食對EPOC參數的影響采用T-Test分析。
2 結果
2.1 體重、心臟指數和鰓指數
實驗魚的體重在對照組和各個訓練組之間沒有顯著性差異(表 1)。4 BL/s訓練組的心臟指數(0.99‰)分別比對照組、1和2 BL/s訓練組顯著提高了10.7%、11.5%和12.5% (分別為0.89‰、0.88‰和0.87‰)(P<0.05)(圖 1a)。4 BL/s訓練組的鰓指數(11.1‰)分別比對照組、1和2 BL/s訓練組顯著提高了7.0%、9.2%和7.5%(分別為10.3‰、10.1‰和10.3‰)(P<0.05)(圖 1b)。
2.2 力竭運動前代謝率
在禁食狀態下, 1、2和4 BL/s訓練組的運動前代謝率[分別為65.5、63.6和62.1 mg O2/(kg·h)]分別比對照組[52.1 mg O2/(kg·h)]顯著提高了25.7%、22.1%和19.2%; 在攝食狀態下, 1、2和4 BL/s訓練組的運動前代謝率[分別為103.9、105.9和111.8 mg O2/(kg·h)]分別比對照組[87.4 mg O2/(kg·h)]顯著提高了18.9%、21.2%和27.9%(P=0.001)(表 1); 攝食狀態下對照組、1、2和4 BL/s訓練組的運動前代謝率分別顯著高于禁食狀態下對照組、1、2和4 BL/s訓練組(P<0.001)(表 1)。攝食和運動對力竭運動前代謝率沒有交互作用(表 1)。
2.3 力竭運動后代謝率特征
在力竭運動后, 所有組的代謝率都迅速上升達到峰值, 隨后急劇下降并逐步恢復到運動前水平(圖 2)。在禁食狀態下, 4 BL/s訓練組的運動后代謝峰值[337.3 mg O2/(kg·h)]顯著高于對照組[290.2 mg O2/(kg·h)]; 在攝食狀態下, 1、2和4 BL/s訓練組的運動后代謝峰值[分別為339.1、343.6和374.9 mg O2/(kg·h)]顯著高于對照組[288.9 mg O2/(kg·h)] (P<0.001)(表 1); 攝食狀態下對照組、1、2和4 BL/s訓練組的運動后代謝峰值與禁食狀態下對照組、1、2和4 BL/s訓練組無顯著性差異(表 1)。攝食和運動對運動后代謝峰值沒有交互作用(表 1)。

表 1 有氧運動訓練和攝食對中華倒刺鲃幼魚力竭運動后代謝率的影響(平均值±標準誤)Tab. 1 The effect of aerobic exercise and feeding on excess post-exercise oxygen consumption (EPOC) response in juvenile Spinibarbus sinensis (Mean±SE)
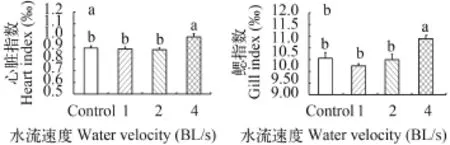
圖 1 有氧運動訓練對中華倒刺鲃幼魚心臟和鰓指數的影響(平均值±標準誤)Fig. 1 The effects of aerobic exercise on the heart and gill indexes in juvenile Spinibarbus sinensis (Mean±SE)
在禁食狀態下, 4 BL/s訓練組的代謝率增量[279.2 mg O2/(kg·h)]顯著高于對照組[236.7 mg O2/(kg·h)]; 在攝食狀態下, 1、2和4 BL/s訓練組的代謝率增量[分別為235.2、237.3和263.3 mg O2/ (kg·h)]顯著高于對照組[201.5 mg O2/(kg·h)](P= 0.002)(表 1); 在攝食狀態下對照組的代謝率增量顯著低于禁食狀態下對照組(P=0.022)(表 1)。攝食和運動對代謝率增量沒有交互作用(表 1)。
不論是禁食還是攝食狀態, 對照組、1、2和4 BL/s訓練組的過量耗氧之間都沒有顯著差異(表1); 在攝食狀態下對照組的過量耗氧(73.7 mg O2/kg)顯著降低于禁食狀態下對照組(102.2 mg O2/kg) (P=0.001), 但在攝食狀態下1、2和4 BL/s訓練組的過量耗氧分別與禁食狀態下1、2和4 BL/s訓練組無顯著性差異(表 1)。攝食和運動對過量耗氧沒有交互作用(表 1)。
3 討論
3.1 有氧運動訓練對中華倒刺鲃幼魚力竭運動后代謝特征的影響
在禁食狀態下的運動前代謝率主要反映動物維持體液循環、蛋白質合成和滲透壓調節等基本生命活動的最低能量需求, 在一些文獻中也稱為維持代謝率[20]。持續的有氧訓練通常導致哺乳動物禁食狀態下的運動前代謝率的顯著上升[21]。不過有氧運動訓練對魚類禁食狀態下的運動前代謝率的影響并沒有得到一致性的結論。比如虹鱒(Oncorhynchus mykiss)[22]、黃尾(Seriola lalandi)[23]和斑馬魚(Danio rerio)[24]有氧運動訓練后其在禁食狀態下的運動前代謝率分別表現為下降、上升和不變。本研究發現, 不同強度的有氧運動訓練都顯著提高了中華倒刺鲃幼魚的維持代謝耗能(表 1), 這與我們以往的研究結果相一致[16]。然而無氧運動訓練并沒有引起中華倒刺鲃幼魚禁食狀態下的運動前代謝率的顯著變化[25]。因此, 運動訓練對魚類禁食狀態下的運動前代謝率的影響可能不僅與其種類相關, 還受到訓練方式的影響。
本研究發現, 通過力竭運動誘導的中華倒刺鲃幼魚代謝峰值[大約290—330 mg O2/(kg·h)]與有氧運動(持續游泳)誘導的最大代謝率[大約300 mg O2/(kg·h)]相似[13], 因此可以作為其最大有氧代謝能力的評價指標。高強度有氧運動訓練(4 BL/s)導致中華倒刺鲃幼魚禁食狀態下運動代謝峰值顯著提高(表 1), 這種結果與虹鱒[26]和大鱗大馬哈魚(Oncorhynchus tshawytscha)[27]的研究結果相似。魚類的有氧代謝能力與其呼吸和循環系統功能密切相關,心臟和鰓功能的改善將有利于魚類對氧氣的吸收和轉運[27]。因此, 有氧運動訓練后中華倒刺鲃幼魚心臟和鰓功能的顯著提高可能是促進其有氧代謝能力增加的重要原因(圖 1a、1b)。這與運動訓練后瓦氏黃顙魚(Peltebagrus vachelli)的研究結果相似[7]。此外, 本研究還發現, 在攝食狀態下, 不同強度的有氧運動訓練都顯著提高了中華倒刺鲃幼魚的運動代謝峰值, 這可能與攝食導致運動前代謝率的顯著提高有關(表 1)。
一般認為, 持續的有氧運動訓練能夠提高魚類的有氧運動能力, 而無氧運動訓練則有利于改善其無氧運動能力[1]。有氧運動訓練能夠影響魚類磷酸肌酸的還原、兒茶酚胺和皮質醇等激素的釋放以及乳酸清除速率, 這些因素與EPOC大小密切相關,進而可能影響魚類無氧運動及其代謝能力[1,28]。不過有關有氧運動訓練對魚類無氧運動及其代謝能力的研究鮮有報道。本研究發現, 不同強度的有氧運動訓練都沒有對中華倒刺鲃幼魚的EPOC大小產生顯著影響, 表明其無氧代謝能力并沒有改善。目前為止, 無氧運動訓練對魚類無氧運動能力的影響也沒有得到一致的結論。比如無氧運動訓練促進代謝產物的轉化提高了錦鯽(Carassius auratus)的無氧運動能力[29], 但卻沒有對瓦氏黃顙魚和南方鲇(Silurus meridionalis)的EPOC的大小產生顯著影響[7,30]。由此表明, 運動訓練對魚類無氧運動能力的影響可能與其種類相關。更多有氧運動訓練對不同習性魚類EPOC大小的影響及其生理生化機制研究可能有利于揭示其內在原因。

圖 2 有氧運動訓練和攝食對中華倒刺鲃幼魚力竭運動后代謝反應的影響Fig. 2 The effect of aerobic exercise and feeding on excess post-exercise oxygen consumption response in juvenile Spinibarbus sinensis平均值±標準誤, a. 對照禁食組; b. 1 BL/s訓練禁食組; c. 2 BL/s訓練禁食組; d. 4 BL/s訓練禁食組; e. 對照攝食組; f. 1 BL/s訓練攝食組; g. 2 BL/s訓練攝食組; h. 4 BL/s訓練攝食組Mean±SE, a. control group in the fasting treatment; b. 1 BL/s exercise group in the fasting treatment; c. 2 BL/s exercise group in the fasting treatment; d. 4 BL/s exercise group in the fasting treatment; e. control group in the fed treatment; f. 1 BL/s exercise group in the fed treatment; g. 2 BL/s exercise group in the fed treatment; h. 4 BL/s exercise group in the fed treatment
3.2 攝食對中華倒刺鲃幼魚力竭運動后代謝特征的影響
由于消化、吸收和同化等生理過程的能量支出增加, 攝食通常會顯著提高動物的代謝率[31]。本研究發現, 攝食導致中華倒刺鲃幼魚運動前代謝率顯著提高(表 1), 這與以往關于中華倒刺鲃等許多魚類攝食后代謝的研究結果相似[9,16,19]。
從理論上來說, 由于循環和呼吸系統等的限制,每種動物都存在一個潛在的最大代謝率, 它是動物進行各種生理活動的有效代謝空間。在魚類的生活史中, 攝食和運動這兩種生理活動經常同時發生,因此, 不同魚類表現出不同的代謝分配模式[11,12]。以往的研究發現, 攝食和運動同時發生才能誘導出鯉魚(Cyprinus carpio)的最大代謝率, 表明其代謝空間能夠在一定程度上同時滿足攝食和運動兩種生理功能的代謝需求, 表現為代謝“添加模式”[12]。本研究發現, 單獨的運動誘導中華倒刺鲃幼魚的最大代謝率[禁食狀態下對照組的運動代謝峰值, 290.2 mg O2/(kg·h)]顯著高于單獨攝食誘導的最大代謝率[攝食狀態下對照組的運動前代謝率, 87.4 mg O2/(kg·h)],與攝食和運動同時誘導的最大代謝率[在攝食狀態下對照組的運動代謝峰值, 288.9 mg O2/(kg·h)]相當。由此表明, 當攝食和運動兩種生理功能同時進行時, 二者在能量代謝水平上可能表現出“競爭模式”。這與草魚(Ctenopharyngodon idellus)的研究結果相似, 可能與其鰓的攝氧能力以及循環系統的輸氧能力的限制有關[9]。
以往的研究發現, 攝食導致南方鲇和鲇(Silurus asotus)等擁有較高消化代謝能力魚類EPOC大小顯著降低, 而沒有對草魚和瓦氏黃顙魚等擁有較低消化代謝能力魚類的EPOC大小產生顯著影響,因此研究者認為攝食對魚類無氧代謝能力的影響可能與其消化代謝能力的高低有關[8,9]。本研究發現, 中華倒刺鲃幼魚最大攝食量(1.5%體重水平)和攝食代謝峰值[攝食狀態下對照組的運動前代謝率, 87.4 mg O2/(kg·h)]都較低, 暗示其消化代謝能力較低。不過, 攝食對其EPOC大小卻產生了負面影響(表 1)。由于EPOC的組成成分較為復雜, 不僅涉及到氧氣的攝入與轉運, 還與乳酸和糖原代謝以及滲透壓調節等因素相關[5], 因此, 攝食對中華倒刺鲃幼魚EPOC產生顯著影響的具體原因有待進一步研究。
[1]Davison W. The effects of exercise training on teleost fish, a review of recent literature [J]. Comparative Biochemistry and Physiology, 1997, 117(1): 67—75
[2]Pearson M P, Spriet L L, Stevens E D. Effect of sprint training on swim performance and white muscle metabolism during exercise and recovery in rainbow trout (Salmo gairdneri) [J]. Journal of Experimental Biology, 1990, 149(1): 45—60.
[3]B?rsheim E, Bahr R. Effect of exercise intensity, duration and mode on post-exercise oxygen consumption [J]. Sports Medicine, 2003, 33(14): 1037—1060
[4]Gaesser G A, Brooks G A. Metabolic bases of excess post-exercise oxygen consumption: a review [J]. Medicine and Science in Sports and Exercise, 1984, 16(1): 29—43
[5]Kieffer J D, Wakefield A M, Litvak M K. Juvenile sturgeon exhibit reduced physiology responses to exercise [J]. Journal of Experimental Biology, 2001, 204(24): 4281—4289
[6]Cutts C J, Metcalfe N B, Taylor A C. Juvenile Altantic salmon (Salmo salar) with relatively high standard metabolic rates have small metabolic scopes [J]. Functional Ecology, 2002, 16(1): 73—78
[7]Liu Y, Cao Z D, Fu S J, et al. The effect of exhaustive chasing training and detraining on swimming performance in juvenile darkbarbel catfish (Peltebagrus vachelli) [J]. Journal of Comparative Physilolgy B, 2009, 179(7): 847—855
[8]Li K G, Cao Z D, Fu S J. Effect of feeding on excess post-exercise oxygen consumption in juvenile Chinese catfish (Silurus asotus Linnaeus) [J]. Acta Hydrobiologica Sinica, 2012, 36(6): 1036—1040 [李可貴, 曹振東,付世建. 攝食對鲇魚幼魚力竭性運動后過量耗氧的影響. 水生生物學報, 2012, 36(6): 1036—1040]
[9]Fu S J, Zeng L Q, Li X M, et al. Effect of meal size on excess post-exercise oxygen consumption in fishes with different locomotive and digestive performance [J]. Journal of Comparative Physiology B, 2009, 179(4): 509—517
[10]Zeng L Q, Zhang Y G, Cao Z D, et al. Effect of temperature on excess post-exercise oxygen consumption in juvenile southern catfish (Silurus meridionalis Chen) following exhaustive exercise [J]. Fish Physiology and Biochemistry, 2010, 36(4):1243—1252
[11]Li X M, Cao Z D, Peng J L, et al. The effect of exercise training on the metabolic interaction between digestion and locomotion in juvenile darkbarbel catfish (Peltebagrus vachelli) [J]. Comparative Biochemistry and Physiology A, 2010, 156(1): 67—73
[12]Pang X, Cao Z D, Fu S J. The effects of temperature on metabolic interaction between digestion and locomotion in juveniles of three cyprinid fish (Carassius auratus, Cyprinus carpio and Spinibarbus sinensis) [J]. Comparative Biochemistry and Physiology A, 2011, 159(3): 253—260
[13]Pang X, Yuan X Z, Cao Z D, et al. The effects of temperature and exercise training on swimming performance in juvenile qingbo (Spinibarbus sinensis) [J]. Journal of Comparative Physiology B, 2013, 183(1): 99—108
[14]Cai Y Z, He C R, Cai Y Q, et al. A primary study of biology of Spinibarbus sinensis Bleeker [J]. Freshwater Fisheries, 2003, 33(3): 16—18 [蔡焰值, 何長仁, 蔡燁強,等. 中華倒刺鲃生物學初步研究. 淡水漁業, 2003, 33(3): 16—18]
[15]Cai Y Z, Cai Y Q, He C R. Experiments on artificial propagation of Spinibarbus sinensis [J]. Freshwater Fisheries, 2005, 35(1): 35—38 [蔡焰值, 蔡燁強, 何長仁. 中華倒刺鲃人工繁殖技術研究. 淡水漁業, 2005, 35(1): 35—38]
[16]Li X M, Yu L J, Wang C, et al. The effect of aerobic exercise training on growth performance, digestive enzyme activities and postprandial metabolic response in juvenile qingbo (Spinibarbus sinensis) [J]. Comparative Biochemistry and Physiology A, 2013, 166(1): 8—16
[17]Li X M , Yuan J M , Zeng L Q, et al. The effects of aerobic exercise on gut evacuation of juvenile qingbo (Spinibarbus sinensis) and the associated mathematical models [J]. Acta Hydrobiologica Sinica, 2015, 39(5): 1012—1018 [李秀明, 袁建明, 曾令清, 等. 有氧運動訓練對中華倒刺鲃幼魚腸道排空及其數學模型選擇的影響. 水生生物學報, 2015, 39(5): 1012—1018]
[18]Steffensen J F. Some errors in respirometry of aquatic breathers: how to avoid and correct for them [J]. Fish Physiology and Biochemistry, 1989, 6(1): 49—59
[19]Fu S J, Cao Z D, Peng J L. Effect of meal size on postprandial metabolic response in Chinese catfish (Silurus asotus Linnaeus) [J]. Journal of Comparative Physiology B, 2006, 176(5): 489—495
[20]Johnston I A. Growth and metabolism in Antarctic fish [J]. Antarctic Special Topic, 1993, 141—150
[21]Speakman J R, Selman C. Physical activity and resting metabolic rate [J]. Proceedings of the Nutrition Society, 2003, 62(3): 621—634
[22]Skov P V, Larsen B K, Frisk M, et al. Effects of rearing density and water current on the respiratory physiology and haematology in rainbow trout, Oncorhynchus mykiss at high temperature [J]. Aquaculture, 2011, 319(3—4): 446—452
[23]Brown E J, Bruce M, Pether S, et al. Do swimming fish always grow fast? Investigating the magnitude and physiological basis of exercise-induced growth in juvenile New Zealand yellowtail kingfish, Seriola lalandi [J]. Fish Physiology and Biochemistry, 2011, 37(2): 327—336
[24]Bagatto B, Pelster B, Burggren W W. Growth and metabolism of larval zebrafish: effects of swim training [J]. Journal of Experimental Biology, 2001, 204(24): 4335— 4343
[25]Li X M, Yu L J, Cao Z D, et al. The effects of exhaustive chasing training on the growth performance and postprandial metabolic response in juvenile qingbo (Spinibarbus sinensis) and rock carp (Procypris rabaudi) [J]. Freshwater Fisheries, 2013, 43(1): 63—68 [李秀明, 于麗娟,曹振東, 等. 力竭追趕訓練對兩種鯉科魚類生長和攝食代謝的影響. 淡水漁業, 2013, 43(1): 63—68]
[26]McKenzie D J, H?glund E, Dupont-Prinet A, et al. Effects of stocking density and sustained aerobic exercise on growth, energetics and welfare of rainbow trout [J]. Aquaculture, 2012, 338: 216—222.
[27]Gallaugher P E, Thorarensen H, Kiessling A, et al. Effects of high intensity exercise training on cardiovascular function, oxygen uptake, internal oxygen transport and osmotic balance in chinook salmon (Oncorhynchus tshawytscha) during critical speed swimming [J]. The Journal of Experimental Biology, 2001, 204(16): 2861—2872
[28]Young P S, Cech J J. Effects of exercise conditioning on stress responses and recovery in cultured and wild youngof-the-year striped bass, Morone saxatilis [J]. Canadian Journal of Fisheries and Aquatic Sciences, 1993, 50(10): 2094—2099
[29]Xia W, Fu S J, Peng J L, et al. The effect of exhaustive exercise training on the anaerobic metabolism for juvenile crucian carp [J]. Journal of Chongqing Normal University (Natural Science), 2011, 28(4): 16—22 [夏偉, 付世建, 彭姜嵐, 等. 力竭運動訓練對錦鯽幼魚無氧代謝能力的影響. 重慶師范大學學報(自然科學版), 2011, 28(4): 16—22]
[30]Cao Z D, Fu S J. The Effect of exhaustive exercise training and fasting on post-exercise oxygen consumption rate in southern catfish (Silurus meridionalis Chen) [J]. Acta Hydrobiologica Sinica, 2009, 33(5): 837—843
[31]McCue M D. Specific dynamic action: a century of investigation [J]. Comparative Biochemistry and Physiology A, 2006, 144(4): 381—394
THE EFFECT OF AEROBIC EXERCISE TRAINING AND FEEDING ON EXCESS POST-EXERCISE OXYGEN CONSUMPTION IN JUVENILE QINGBO (SPINIBARBUS SINENSIS)
LI Xiu-Ming1,2, YUAN Jian-Ming2,3, ZHANG Yao-Guang2and FU Shi-Jian1
(1. Laboratory of Evolutionary Physiology and Behavior, Chongqing Key Laboratory of Animal Biology, Chongqing Normal University, Chongqing 401331, China; 2. Key Laboratory of Freshwater Fish Reproduction and Development, Ministry of Education, Southwest University, Chongqing 400715, China; 3. Fisheries Technology Extension Station in Chongqing, Chongqing 400020, China)
To investigate the effects of aerobic exercise and feeding on excess post-exercise oxygen consumption (EPOC) in juvenile qingbo (Spinibarbus sinensis), 120 fish [(21.35±0.05) g, (10.21±0.03) cm] were exercised at 25℃under four water velocities, i.e., nearly still water (control), 1 body length (BL)/s, 2 BL/s and 4 BL/s for eight weeks to measure the heart index, gill index and EPOC. The results showed that the heart and gill indexes significantly increased in 4 BL/s training group compared to those in other three groups (P<0.05). Aerobic exercise resulted in a significant increase in the pre-exercise metabolic rate under both feeding and fasting status (P=0.001). There were no differences in the EPOC magnitude of S. sinensis between the exercise groups and control group under both feeding and fasting status. Peak post-exercise metabolic rate and increment of metabolic rate were significantly higher in the 4 BL/s training group than those of the control group under fasting status. Peak post-exercise metabolic rates and increment of metabolic rates were significantly higher in the three exercise groups than those of the control group under feeding status (P<0.005). Feeding resulted in a significantly higher pre-exercise metabolic rate compared to fasting fish under the same water velocity (P<0.001). Peak post-exercise metabolic rate is similar between the feeding and fasting treatment groups with the same water velocity. Feeding led to a significant decrease in the increment of metabolic rate and EPOC magnitude in the control group (P<0.05). Our results suggest that aerobic exercise significantly increased aerobic capacity, which may be related to the improvement of respiratory and circulatory system function, and that exhaustive exercise can induce maximum oxygen consumption rate, and that feeding reduced the anaerobic capacity in juvenile S. sinensis.
Aerobic exercise training; Feeding; Excess post-exercise oxygen consumption; Metabolic rate; Spinibarbus sinensis
Q493
A
1000-3207(2017)02-0277-08
10.7541/2017.34
2015-12-29;
2016-04-17
國家自然科學基金(31302160, 31172096); 高等學校學科創新引智計劃(111計劃)建設項目(B14037); 中國博士后科學基金(2014M562271); 重慶市科委重大項目(cstc2014yykfC80001); 重慶市基礎與前沿研究計劃項目(cstc2015jcyjA80007); 重慶市教育委員會科學技術研究項目(KJ1400506); 重慶師范大學基金項目(14XLB001)資助 [Supported by the National Natural Science Foundation of China (31302160, 31172096); the 111 Project (B14037); China Postdoctoral Science Foundation (2014M562271); the Major Projects of Chongqing Science and Technology Committee (cstc2014yykfC80001); the Basic and Frontier Research Project of CQ (cstc2015jcyjA80007); Science and Technology Research Project of Chongqing Education Committee (KJ1400506); the Foundation Project of Chongqing Normal University (14XLB001)]
李秀明(1979—), 男, 重慶市人; 博士; 研究方向為魚類生理生態和水產養殖。E-mail: lixiuming_441188@126.com
付世建, 教授; E-mail: shijianfu9@hotmail.com
