煙草鉀離子通道基因NtTPK的克隆及表達分析
2017-04-08 03:00:03黃路平魯黎明李立芹
浙江農業學報 2017年3期
許 力,黃路平,魯黎明,李立芹
(四川農業大學 農學院,四川 成都 625014)
煙草鉀離子通道基因NtTPK的克隆及表達分析
許 力,黃路平,魯黎明,李立芹*
(四川農業大學 農學院,四川 成都 625014)
采用同源克隆的方法對煙草品種K326中的一個鉀離子通道基因NtTPK進行了研究,該基因序列全長1 317 bp,編碼438個氨基酸。實時熒光定量PCR結果表明,該基因在莖中的表達量最高;在低鉀處理24 h后該基因表達量最低;200 mmol·L-1NaCl處理24 h后該基因表達量最高;在煙草打頂7 d后,該基因在葉和根中的表達量達到最高。這些結果為進一步研究該基因的功能提供了一定理論基礎。
煙草;鉀離子通道;NtTPK;生物信息學;基因表達
鉀是植物生長發育所必需的大量元素之一,也是一些酶的激活劑,所以它在植物細胞光合作用、蛋白質合成和氧化代謝等一些生理過程中起重要作用[1-2]。K+是植物細胞中含量最豐富的陽離子,但不屬于有機結構的組分,在植物中含量很高,可占到植物總干質量的10%左右,新鮮植物組織細胞內的K+濃度高達100~200 mmol·L-1[3-4]。如果植物在生長過程中缺鉀則其生長就會受到限制,因為鉀缺乏會使依賴K+的酶活性降低,從而導致植物生理代謝紊亂[5]。由此可見,充足的K+對植物生長具有重要意義。煙草是一種需鉀量很大的作物,煙葉含鉀量是衡量煙葉品質的重要指標之一[6]。鉀除了作為重要的營養元素參與煙葉的生理生化反應外,它還可以增加煙葉的燃燒性并因此降低煙制品的焦油含量,從而提高其安全性。但在我國北方煙區的煙葉含鉀量大都不足2%[7],這直接影響到煙葉的產量和品質。
鉀轉運體/通道是植物生理功能上的重要參與者,包括細胞信號傳導、滲透調節、植物營養和金屬耐受性[8]。植物從土壤中吸收K+及在體內的轉運是通過鉀離子通道和鉀離子轉運體完成的[9]。根據鉀離子通道的結構和功能不同可將其分為三大類:Shaker家族通道、TPK/KCO通道和其他鉀離子通道[10]。模式植物擬南芥中TPK/KCO通道包括兩孔鉀離子通道TPK(也稱串聯孔通道)和一個Kir-型通道。TPK1和TPK4是擬南芥中兩孔鉀離子通道家族中研究較多的兩個成員,TPK1定位在液泡膜上,受Ca2+激活,是選擇性K+通道[11];而TPK4定位在質膜上,不具外向整流特點[12];信號分子(如細胞內的H+與Ca2+)和物理因素(如溫度與壓力)可以調節植物兩孔鉀通道活性[13]。目前煙草TPK/KCO家族中僅NtTPK1和NtTPK2有報道。因此,對鉀離子通道TPK/KCO家族成員的研究具有重要理論意義。本研究通過同源克隆的方法從煙草品種K326中成功克隆到NtTPK基因,并對其進行了生物信息學分析,以及分析其組織表達及在低鉀、高鹽處理下及打頂前后不同時期的表達情況,以期為今后NtTPK的功能研究及其鉀營養分子機制提供重要的理論基礎。
1 材料與方法
1.1 試驗材料與處理
植物材料為煙草品種K326,首先進行漂浮育苗,待其生長到十字期,將其移栽到花盆中,每盆種植一株。對打頂前與打頂后煙株的根、葉以及正常生長的根、莖、葉和花進行取樣,然后在液氮中速凍,置于-80 ℃冰箱,以供后續試驗使用。
將煙草品種K326種子表面消毒后布種于MS培養基上,培養15 d后,將長勢一致的幼苗分別轉移到低鉀(10 μmol·L-1)和高鹽(200 mmol·L-1NaCl)的培養基上,處理0、6、12、24 h,然后進行整株取樣。低鉀培養基是去掉MS培養基中的KNO3,并且用NH4H2PO4代替KH2PO4,其余成分相同。高鹽培養基是在MS培養基的基礎上添加200 mmol·L-1NaCl。
通用型DNA純化回收試劑盒購自天根公司,DH5α大腸埃希菌菌株購于Vazyme Biotech公司,cDNA合成試劑盒購于Thermo Scientific公司,Trizol試劑、高保真酶R045A、載體pMD19-T、Prime Script RT reagent Kit、SYBR Green Master mix等試劑均購自大連寶生物公司,引物合成與測序由上海生工完成。
1.2 試驗方法
1.2.1 煙草RNA的提取及cDNA的合成
按照TaKaRa公司的RNA提取用Trizol試劑盒的說明書提取煙草RNA。以RNA為模板,按照cDNA合成試劑盒的操作說明完成cDNA的合成。反轉錄條件為:42 ℃ 60 min,72 ℃10 min。
1.2.2NtTPK基因的克隆
根據絨毛狀煙草兩孔鉀離子通道基因序列,設計引物NtTPKF和NtTPKR(表1),擴增反應程序為:95 ℃預變性3 min;95 ℃變性30 s,55 ℃退火30 s,72 ℃延伸1.5 min,35個循環。 目的片段純化后與pMD19-T載體16 ℃連接過夜,連接產物轉化大腸埃希菌DH5α 感受態,隨后涂在含氨芐青霉素(Ampicillin)的LB 平板上進行篩選,采用菌落PCR 法檢測陽性克隆,然后送到生工生物公司進行測序。
1.2.3 目的基因的生物信息學分析
采用ExPASy ProtParam tool與 DNAMAN 軟件分析其編碼蛋白的分子量、等電點以及其他理化性質,并進行疏水性分析;運用BioEdit,clustalx-2.0.9及MEGA2.0軟件構建系統進化樹;運用TMHMM Server v. 2.0進行目的基因的跨膜區分析;采用PSORT、NetPhos3.1 Server在線工具、SWISS-MODEL等生物信息學工具對蛋白進行亞定位預測、磷酸化位點及高級結構等的分析。
1.2.4 目的基因的表達分析
根據基因序列,設計目的基因實時熒光定量PCR引物NtTPKqF和NtTPKqR(表1),采用煙草的18S rRNA作為內參進行基因表達差異研究。PCR擴增程序為:50 ℃反應2 min,95 ℃預變性3 min,然后對95 ℃變性10 s和65 ℃延伸45 s進行39個循環。所有的樣品都設置3個重復,反應結束后,基因相對表達水平用2-△△Ct方法進行計算。
2 結果與分析
2.1NtTPK基因的克隆
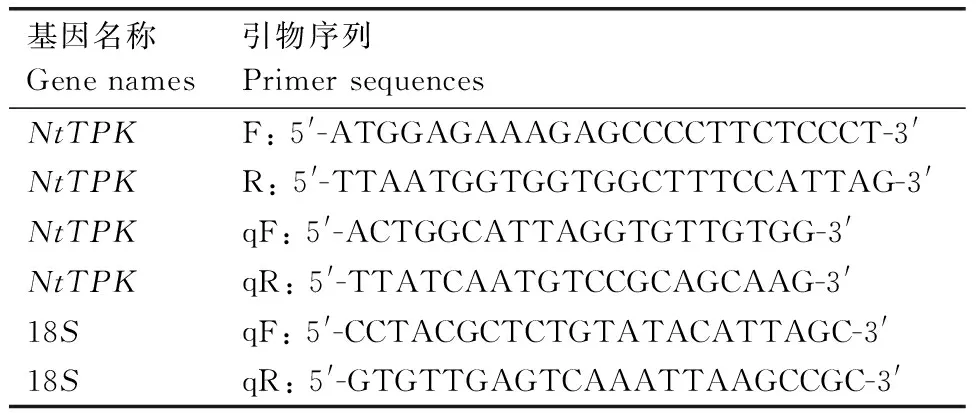
表1 引物序列
首先提取煙草根總RNA,然后反轉錄合成cDNA,以cDNA為模板進行PCR擴增,凝膠電泳結果顯示獲得一條約1 300 bp的特異性條帶(圖1)。采用凝膠回收試劑盒純化該條帶,然后進行連接并轉化大腸埃希菌DH5α,挑取陽性克隆進行測序。
2.2NtTPK基因的組織表達分析
為了檢測NtTPK基因在不同組織的表達水平,首先提取煙草根、莖、葉和花的總RNA,根、莖和葉的取樣是生長60 d的煙草,而花的取樣時期為盛花期,然后通過實時熒光定量PCR實驗分析NtTPK基因在根、莖、葉和花的表達量。結果表明,NtTPK基因根、莖、葉和花中均有表達,在莖中的表達量最高,其次是在根中的表達,在花中的表達量最低,在莖中表達量是在根中的3倍(圖2)。
2.3 目的基因的生物信息學分析
2.3.1NtTPK編碼蛋白的理化性質分析
運用ExPASy ProtParam tool軟件對目的基因編碼的蛋白進行理化性質在線分析。結果顯示,NtTPK蛋白由20種氨基酸構成(其中Leu最多為11.0%,而Trp最少為1.1%)。該預測蛋白的分子量為47.83 ku,分子式為C2188H3423N557O614S20,理論等電點pI為6.60;帶負電荷的氨基酸殘基(Asp+Glu)總數是42,帶正電荷的氨基酸殘基(Arg+Lys)總數是40,消光系數在280 nm時為51 715,NtTPK的脂肪系數為97.49,總的親水性平均系數為0.099。NtTPK蛋白的不穩定系數是34.26,推測它是一個穩定蛋白。
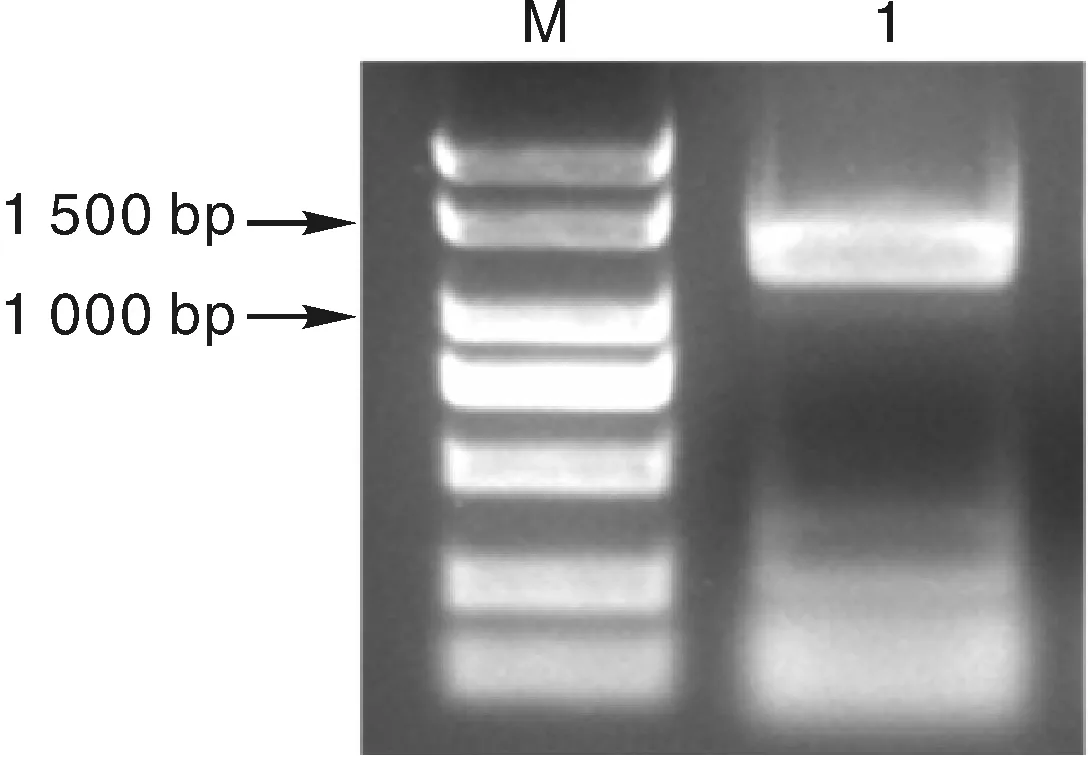
1,NtTPK基因;M,Marker1, NtTPK PCR product; M, Marker圖1 NtTPK基因PCR擴增電泳圖Fig.1 Electrophoresis analysis of NtTPK PCR product
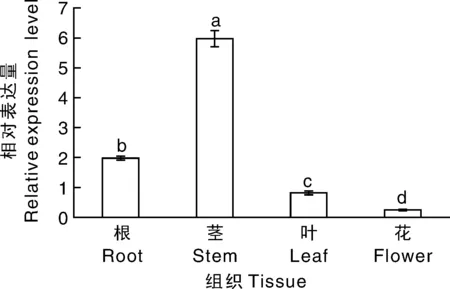
圖2 不同組織中NtTPK基因的相對表達量分析Fig.2 The relative expression analysis of NtTPK in different tissues
2.3.2NtTPK編碼蛋白的跨膜區及疏水性分析
運用TMHMM 2.0對NtTPK編碼蛋白的跨膜區進行預測分析,結果顯示,NtTPK所編碼的蛋白含有5個跨膜區(圖3-A),且在第3個和第4個跨膜區之間均有一個較長的胞質質環。為了進一步分析NtTPK的疏水性,運用DNAMAN軟件對其疏水區域進行預測及分析,結果表明,該蛋白含有較多的疏水區域(圖3-B)。結合NtTPK的疏水性、總的親水性平均系數(0.099)以及其他理化性質綜合分析,可以說明NtTPK是一個疏水性膜蛋白。
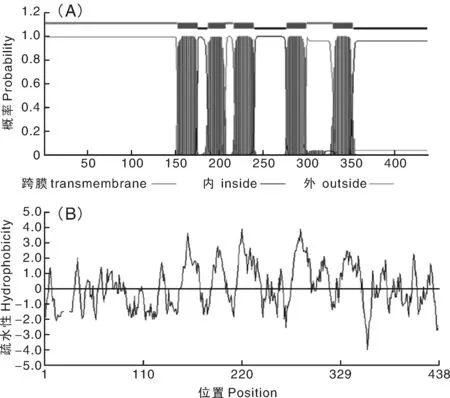
A, 蛋白質跨膜結構分析; B, 蛋白質疏水性分析A, Transmembrane domain analysis of target protein; B, Hydrophobicity analysis of target protein圖3 蛋白質跨膜結構與疏水性分析Fig.3 Transmembrane domain and hydrophobicity analyses of target protein
2.3.3 編碼蛋白的亞細胞定位及磷酸化位點分析
利用PSORT在線工具預測NtTPK的亞細胞定位,結果顯示,NtTPK主要定位在質膜和葉綠體類囊體膜上,在質膜上占0.6,葉綠體類囊體膜上占0.572,在高爾基體上占0.4,在內質網(膜)上占0.3,說明其可能定位在質膜與葉綠體類囊體膜上。利用NetPhos3.1 Server在線工具對NtTPK中的絲氨酸(Serine)激酶、蘇氨酸(Threonine)激酶和酪氨酸(Tyrosine)激酶的磷酸化位點進行分析,結果發現NtTPK中含有24個絲氨酸激酶磷酸化位點,9個蘇氨酸激酶磷酸化位點和3個酪氨酸激酶磷酸化位點(圖4),推測此蛋白能被激酶磷酸化,從而參與鹽脅迫生理過程的調控。
2.3.4 蛋白二級結構及三維結構預測分析
運用IBCP的在線工具SOPMA,對NtTPK的二級結構進行預測,結果顯示,α-螺旋為主要結構占46.42%,而β-折疊占7.92%,延伸鏈占22.64%以及無規則卷曲占23.02%(圖5)。運用SWISS-MODEL生物信息學工具對NtTPK蛋白的三維結構進行預測(圖6)。
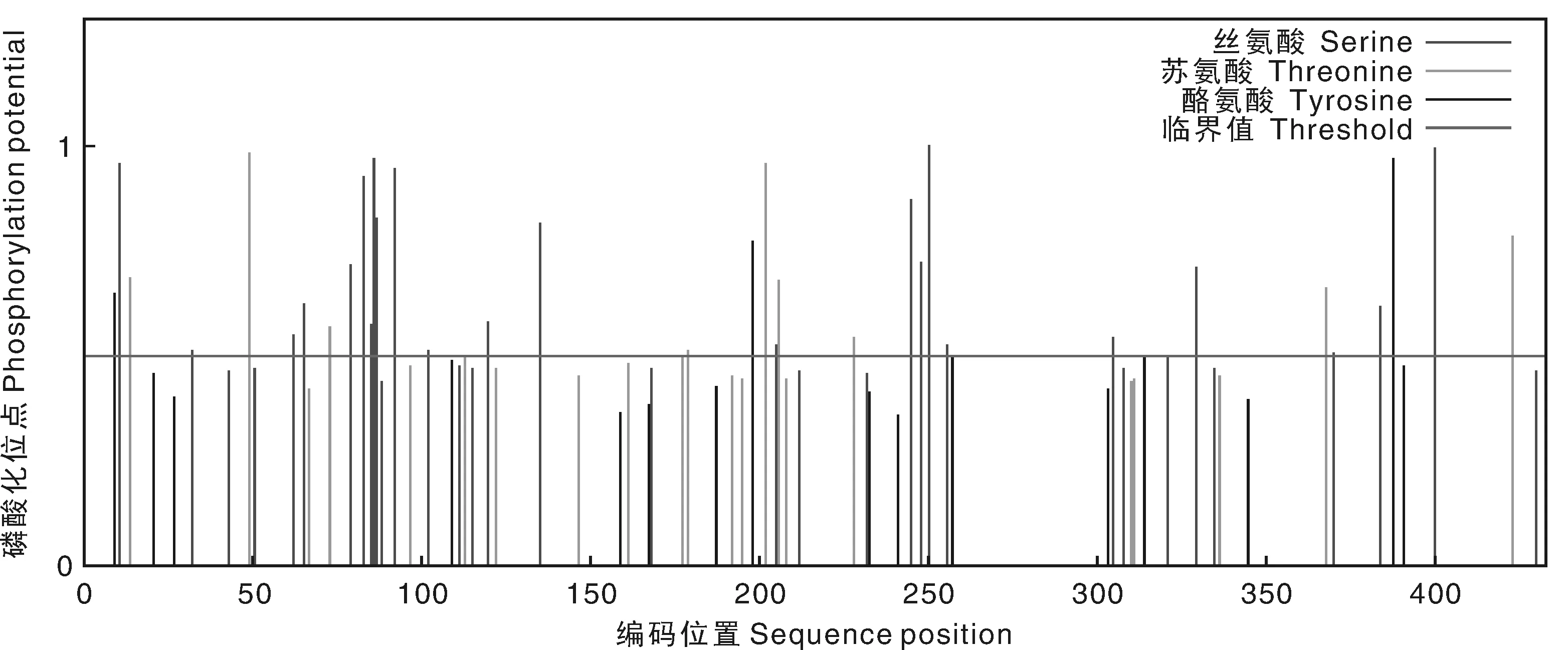
圖4 NtTPK中絲氨酸、蘇氨酸及酪氨酸激酶的磷酸化位點分析Fig.4 Predicted phosphorylation site of serine, threonine and tyrosine kinase in NtTPK
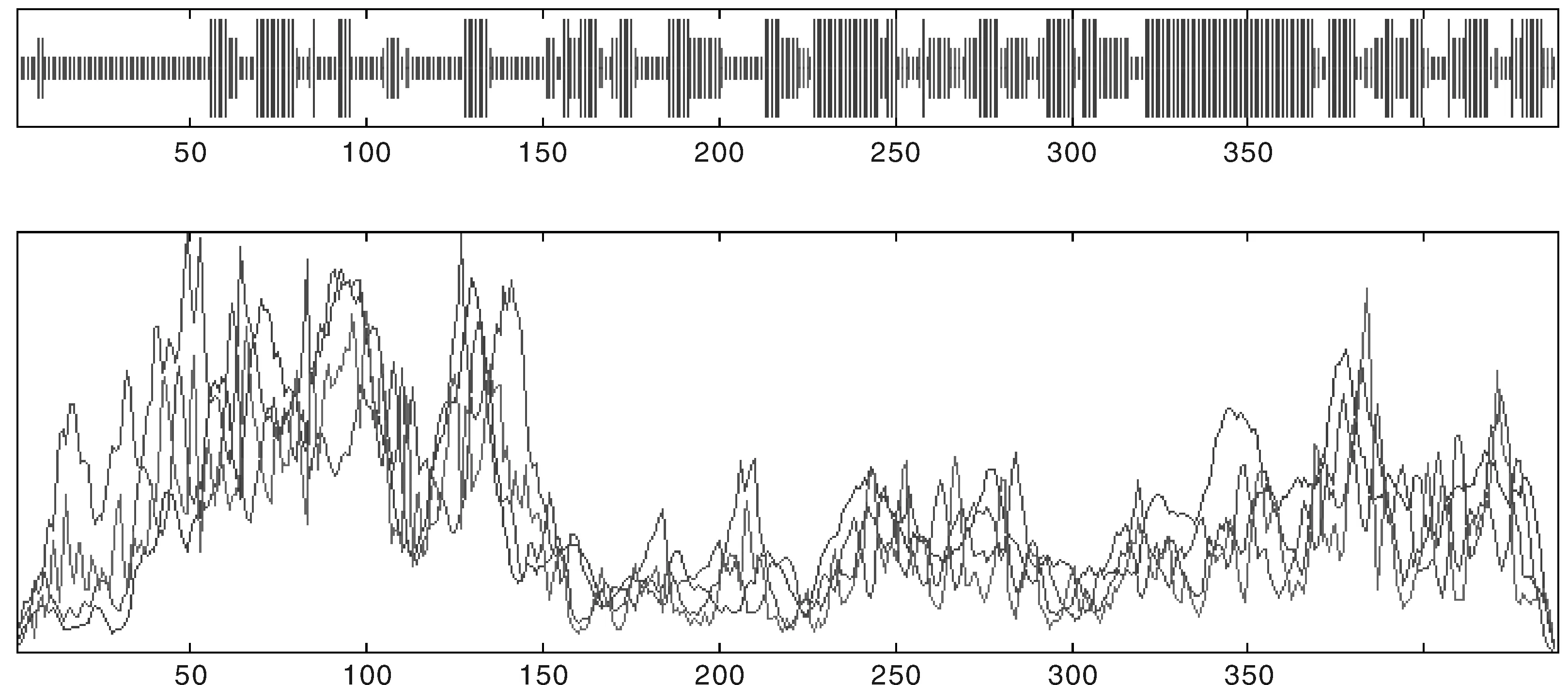
圖5 蛋白質的二級結構預測Fig.5 Predicted secondary structure of target protein
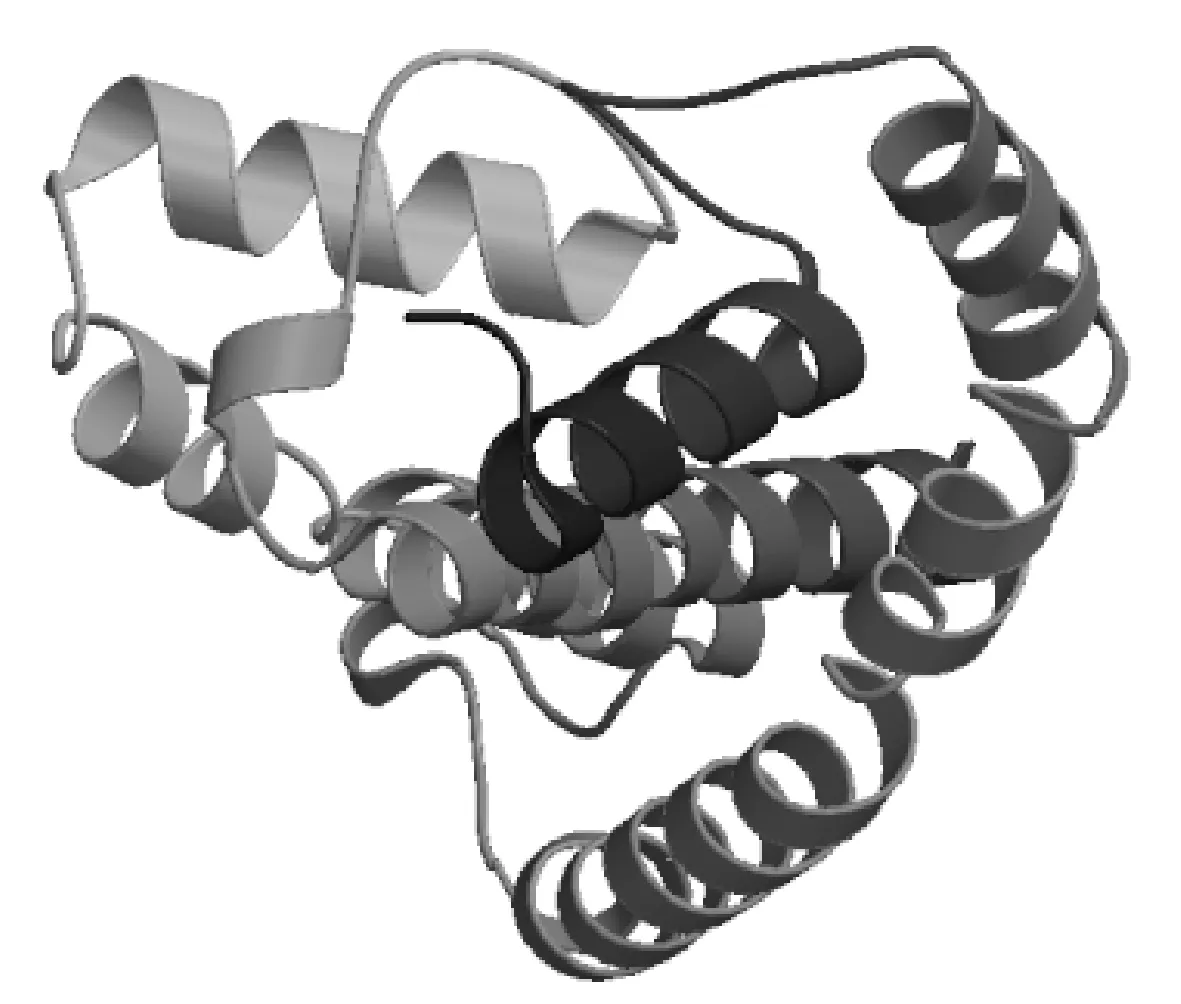
圖6 蛋白質的三維結構預測結果Fig.6 The predicted advanced structure of target protein
2.3.5 NtTPK的序列多重比對及同源性分析
將NtTPK的氨基酸序列在NCBI 數據庫中進行BLAST比對,按照相似性程度高低選擇19個序列,運用BioEdit,clustalx-2.0.9及MEGA2.0軟件構建系統進化樹,結果表明,NtTPK與絨毛狀煙草兩孔鉀離子通道TPK3、普通煙草兩孔鉀離子通道NtTPK3、普通煙草NtTPK1等具有較高的序列同源性(99%、99%、98%),與胡蘿卜兩孔鉀離子通道TPK3的同源性最低,為81%。根據比對的同源蛋白信息(圖7),將克隆的煙草基因命名為NtTPK。2.4 低鉀與高鹽處理下NtTPK基因表達模式分析
首先在超凈工作臺把生長15 d的煙草幼苗轉移到低鉀與高鹽的培養基上處理0、6、12、24 h,然后進行取樣提取RNA。通過實時熒光定量PCR實驗分析該基因在低鉀與高鹽處理后的表達模式,結果顯示,與對照相比,NtTPK基因隨低鉀處理時間的延長其表達量呈下降趨勢,在處理24 h時,其表達量最低,比對照降低5.1倍(圖8)。NtTPK基因隨高鹽處理時間的延長表達量呈上升趨勢,在處理24 h時,其表達量最高,是對照的5.3倍(圖8)。
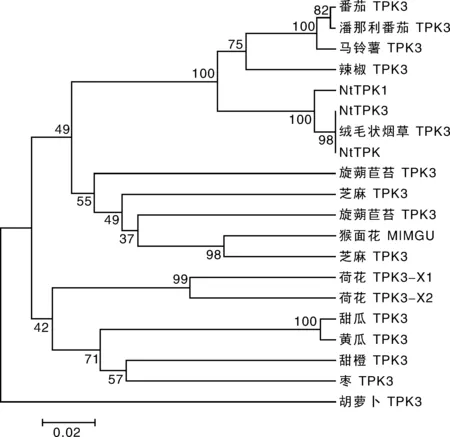
圖7 NtTPK與其他植物鉀離子通道蛋白的同源性分析Fig.7 Homology analysis of NtTPK and other plant potassium channel protein
2.5NtTPK基因在打頂前與打頂后不同時期的表達分析
本試驗通過實時熒光定量PCR實驗分析該基因在煙草打頂后(0、1、3、7 d)不同時期葉片和根中的表達量,煙草打頂的時期為現蕾期,結果顯示,在煙草打頂1、3和7 d后,葉中該基因表達量逐漸增加。7 d時,其表達量最高。在根中,與對照相比,該基因在打頂后1和3 d的表達量基本一致;但在打頂7 d時,NtTPK基因表達量達到最高水平,是對照的14.6倍(圖9)。
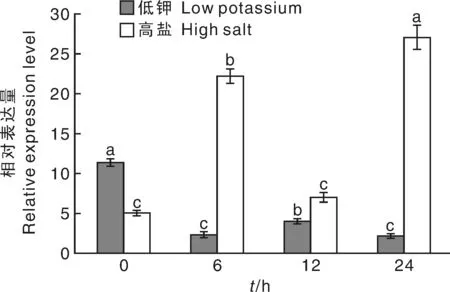
圖8 低鉀與高鹽處理下NtTPK基因表達模式分析Fig.8 Analysis of NtTPK gene expression under low potassiumor high salt treatments
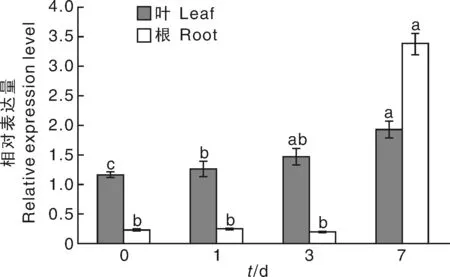
圖9 打頂前后不同時期葉與根中NtTPK基因的相對表達量分析Fig.9 The relative expression analysis of NtTPK in leaves and roots before and after topping stage
3 討論
眾所周知,鉀對煙草生長發育和品質具有重要的影響[14],在本研究中,首先采用同源克隆的方法獲得煙草品種K326中鉀離子通道基因NtTPK的全長CDS序列,長度為1 317 bp,編碼氨基酸438個,對其跨膜區域預測分析顯示,該蛋白具有5個跨膜結構域,這與煙草NtTPK2和胡楊PeTPK1跨膜結構域數目一致[15-16]。蛋白的理化性質及疏水性分析表明,NtTPK可能是疏水性的膜蛋白。亞細胞預測其可能定位在質膜與葉綠體類囊體膜上,這與擬南芥TPK3既定位在質膜也定位在類囊體膜上的雙重定位結果一致[17]。磷酸化位點分析說明該蛋白能被一些激酶磷酸化,這些絲氨酸或蘇氨酸殘基的磷酸化是與一些蛋白(如14-3-3蛋白)相互作用的關鍵[18]。實時熒光定量PCR實驗表明,NtTPK在根、莖、葉和花中均有表達,這與擬南芥AtTPK/KCO和煙草NtTPK2的表達模式是一樣的[15,19],其中該基因在莖中的表達量相對較高,這與NtTPK1基因的表達模式類似[20]。說明NtTPK可能在鉀的轉運方面具有重要的作用。蔡元保等[21]研究表明,在巴西橡膠樹中HbKCO1基因在鉀饑餓處理12和24 h時表達量明顯下降,在本研究中,NtTPK基因在低鉀處理12和24 h后表達量也呈下降趨勢,與他們的結果一致。NtTPK基因在高鹽處理6、12和24 h后,其表達量呈上升趨勢,這與煙草NtTPK1和HbKCO1表達模式類似,這兩個基因的表達也是受高鹽誘導[20-21]。
在生產上對煙草進行打頂是提高煙葉產量的一項重要栽培措施,但打頂會影響煙葉的K+含量[22]。韓錦峰等[23]研究認為,我國煙葉鉀含量低的重要原因是煙草通過根系將K+排到體外,而鉀又不能得到充分、適時、合理的補充所造成的。本試驗中,通過實時熒光定量PCR實驗分析煙草打頂前后不同時期葉和根中NtTPK的基因表達模式,結果表明,在打頂7 d后,NtTPK基因表達量在葉和根中達到最高。代曉燕等[24]研究了煙草品種紅花大金元中內流型NKT1與外流型NTORK1鉀離子通道基因在打頂后的表達模式,發現在打頂5和24 h后,NKT1基因和NTORK1基因在葉中表達量受到抑制,但是,在打頂1和5 h后這兩個基因在根中表達量增加。本研究中的NtTPK與這兩個基因在打頂后的表達模式不完全一致,原因可能是煙草品種不同、取樣時間不同以及鉀離子通道功能不同等。今后進一步研究工作可通過酵母雙雜交尋找NtTPK互作蛋白、利用電壓鉗及轉基因技術等明確NtTPK功能。綜上所述,本研究結果為下一步研究煙草鉀離子通道TPK功能以及了解煙草鉀營養的分子機制提供了一定理論基礎。
本研究從煙草品種K326中克隆到1個煙草鉀離子通道基因,命名為NtTPK。該基因序列全長1 317 bp,編碼438個氨基酸。預測其具有5個跨膜結構域,系統進化樹分析NtTPK與絨毛狀煙草TPK3相似性為99%。NtTPK基因在煙草莖中的表達量最高,低鉀處理抑制該基因的表達,而高鹽處理24 h后表達量明顯上升。在煙草植株打頂7 d后,該基因在葉與根中的表達量達到最高。
[1] MARSCHNER H. Mineral nutrition of higher plants[M]. London: Academic Press, 1995: 889.
[2] WHITE P J, KARLEY A J. Potassium[M]//HELL R, MENDEL R R. Plant cell monographs. Berlin: Springer, 2010: 199-224.
[3] 魯黎明,楊鐵釗.植物K+吸收轉運的分子機制研究進展[J].棉花學報, 2006, 18(6): 379-385. LU L M, YANG T Z. Advances in research of molecular mechnisms of potassium uptaking and transportation in plant[J].CottonScience, 2006, 18(6): 379-385.(in Chinese with English abstract)
[4] WANG Y, WU W H. Potassium transport and signaling in higher plants[J].AnnualReviewofPlantBiology, 2013, 64: 451-476.
[5] SYNSUKE K, REDA E, HANY A. Potassium deficiency affects water status and photosynthetic rate of the vegetative sink in green house tomato prior to its effects on source activity[J].PlantScience, 2011, 180(2): 368-374.
[6] 代曉燕,郭春燕,劉國順,等.鉀肥不同追施時期對烤煙品質和產量的影響[J].浙江農業學報, 2014, 26(2): 424-426. DAI X Y, GUO C Y, LIU G H, et al. Effects of different topdressing periods of potassium fertilization on quality and yields of flue cured tobacco[J].ActaAgriculturaeZhejiangensis, 2014, 26(2): 424-426.(in Chinese with English abstract)
[7] 曹志洪.優質烤煙生產的土壤與施肥[M].南京:江蘇科學技術出版社,1991: 17-28.
[8] PINTO E, FERREIRA I M. Cation transporters/channels in plants: Tools for nutrient biofortification[J].JournalofPlantPhysiology, 2015, 179: 64-82.
[9] HAN M, WU W, WU W H, et al. Potassium transporter KUP7 is involved in K+acquisition and translocation inArabidopsisroot under K+-limited conditions[J].MolecularPlant, 2016, 9: 437-446.
[10] 程鈺宏,趙瑞雪,董寬虎.植物鉀(K+)離子通道的研究[J].山西農業科學, 2008, 36(2): 3-7. CHENG Y H, ZHAO R X, DONG K H. Research on plasma K+channel protein[J].JournalofShanxiAgriculturalSciences, 2008, 36(2): 3-7. (in Chinese with English abstract)
[11] BIHLER H, EING C, HEBEISEN S, et al. TPK1 is a vacuolar ion channel different from the slow-vacuolar cation channel[J].PlantPhysiology, 2005, 139(1):417-424.
[12] MARCEL D, MüLLER T, HEDRICH R, et al. K+transport characteristics of the plasma membrane tandem-pore channel TPK4 and pore chimeras with its vacuolar homologs[J].FEBSLetters, 2010, 584(11): 2433-2439.
[14] 魯黎明.煙草鉀轉運體基因TPK1的電子克隆及生物信息學分析[J].中國農業科學, 2011, 44(1): 28-35. LU L M.Insilicocloning and bioinformatic analysis ofTPK1 gene in tobacco[J].ScientiaAgriculturaSinica, 2011, 44(1): 28-35. (in Chinese with English abstract)
[15] 閆亞飛,王根洪,程廷才,等.煙草鉀離子通道基因NtTPK2的克隆及序列分析[EB/OL]. (2012-03-07) http://www.paper.edu.cn/releasepaper/content/201203-247.
[16] WANG F F, DENG S R, DING M Q, et al. Overexpression of a poplar two-pore K+channel enhances salinity tolerance in tobacco cells[J].PlantCellTissueandOrganCulture, 2013, 112(1): 19-31.
[17] CARRARETTO L, FORMENTIN E, TEARDO E, et al. A thylakoid-located two-pore K+channel controls photosynthetic light utilization in plants[J].Science, 2013, 342(6154): 114-118.
[18] LATZ A, BECKER D, HEKMAN M, et al. TPK1,a Ca2+-regulatedArabidopsisvacuole two-pore K+channel is activated by 14-3-3 proteins[J].ThePlantJournal, 2007, 52(3): 449-459.
[19] VOELKER C, SCHMIDT D B, MUELLER-ROEBER B, et al. Members of the Arabidopsis AtTPK/KCO family form homomeric vacuolar channels in planta[J].ThePlantJournal, 2006, 48(2): 296-306.
[20] HAMAMOTO S, MARUI J, MATSUOKA K,et al. Characterization of a tobacco TPK-type K+channel as a novel tonoplast K+channel using yeast tonoplasts[J].TheJournalofBiologicalChemistry, 2007, 283(4): 1911-1920.
[21] 蔡元保,朱家紅,暢文軍,等.巴西橡膠樹K+通道蛋白基因HbKC01的克隆與表達分析[J]. 農業生物技術學報, 2010, 18(1): 30-36. CAI Y B, ZHU J H, CHANG W J, et al. Cloning and expression analysis ofHbKC01 fromHeveabrasiliensis[J].JournalofAgriculturalBiotechnology, 2010, 18(1): 30-36. (in Chinese with English abstract)
[22] JIANG F, LI C J, JESCHKE W D, et al. Effect of top excision and replacement by 1-naphthylacetic acid on partition and flow of potassium in tobacco plants[J].JournalofExperimentalBotany, 2001, 52(364): 2143-2150.
[23] 韓錦峰,朱大恒,劉華山,等.我國烤煙含鉀量低的原因及解決途徑[J].河南農業科學, 2010, 39(2): 18-23. HAN J F, ZHU D H, LIU H S, et al. Reasons and solutions for low potassium content of flue-cured tobacco in China[J].JournalofHenanAgriculturalSciences, 2010, 39(2):18-23. (in Chinese)
[24] 代曉燕,蘇以榮,魏文學,等.打頂對烤煙植株鉀素代謝和鉀離子通道基因表達的影響[J]. 中國農業科學, 2009, 42(3): 854-861. DAI X Y, SU Y R, WEI W X, et al. Effects of topping on potassium metabolism and expression of potassium channels in tobacco plants[J].ScientiaAgriculturaSinica, 2009, 42(3): 854-861. (in Chinese with English abstract)
(責任編輯 張 韻)
Cloning and expression analysis of potassium channelNtTPKgene in tobacco
XU Li, HUANG Luping, LU Liming, LI Liqin*
(CollegeofAgronomy,SichuanAgriculturalUniversity,Chengdu625014,China)
A new potassium channel geneNtTPKwas cloned from cultivar K326 using homologous cloning strategy. Sequences analysis showed this gene contained 1 317 bp and coded 438 amino acid residues. Quantitative real-time PCR results showed that the highest expression level of this gene was in the stem; and the lowest expression level emerged after 24 h under low potassium treatment; the gene expression reached its peak level after 24 h under 200 mmol·L-1NaCl treatment; and 7 d after topping, the gene expression showed the highest level in leaves and roots of tobacco. These results provide theoretical bases for further study of the function of potassium channelNtTPKgene.
tobacco; potassium channel;NtTPK; bioinformatics; gene expression
10.3969/j.issn.1004-1524.2017.03.03
2016-10-28
植物生理學與生物化學國家重點實驗室開放課題(SKLPPBKF 1505, SKLPPBKF 1506)
許力(1992—),女,四川中江人,碩士研究生,主要從事植物分子生物學研究。E-mail: 892334388@qq.com
*通信作者,李立芹,E-mail: liliqin88@163.com
S572
A
1004-1524(2017)03-0366-07
浙江農業學報ActaAgriculturaeZhejiangensis, 2017,29(3): 366-372
http://www.zjnyxb.cn
許力,黃路平,魯黎明,等. 煙草鉀離子通道基因NtTPK的克隆及表達分析[J].浙江農業學報,2017,29(3): 366-372.
猜你喜歡
奧秘(創新大賽)(2023年3期)2023-05-06 01:48:20
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
浙江中西醫結合雜志(2017年2期)2017-01-12 18:23:59
山東工業技術(2016年15期)2016-12-01 05:31:22
當代化工研究(2016年9期)2016-03-20 16:22:08
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
