楊樹不同種質花粉萌發和致敏蛋白的差異*
2017-03-27 02:58:45章小鈴王留強吳立栓盧孟柱胡建軍
林業科學 2017年2期
關鍵詞:差異
章小鈴 王留強 孫 佩 吳立栓 樊 瑋 張 進 盧孟柱 胡建軍
(1. 林木遺傳育種國家重點實驗室 國家林業局林木培育重點實驗室 中國林業科學研究院林業研究所 北京 100091; 2. 中國科學院植物研究所光生物學重點實驗室 北京 100093)
?
楊樹不同種質花粉萌發和致敏蛋白的差異*
章小鈴1王留強1孫 佩1吳立栓2樊 瑋1張 進1盧孟柱1胡建軍1
(1. 林木遺傳育種國家重點實驗室 國家林業局林木培育重點實驗室 中國林業科學研究院林業研究所 北京 100091; 2. 中國科學院植物研究所光生物學重點實驗室 北京 100093)
【目的】 研究6份楊樹種質(36號楊、南楊、110楊、森海2號楊、小葉楊和鉆天楊)成熟花粉的萌發特性以及致敏蛋白差異,為低致敏楊樹品種選育及植物過敏性疾病控制等研究提供理論依據。【方法】 2015 年3月,在北京、河北和河南采集6份不同楊樹種質的花枝,經水培后收集花粉,采用液體培養法進行楊樹花粉萌發試驗,并通過雙向電泳、實時熒光定量PCR等研究不同楊樹種質的花粉致敏蛋白差異。【結果】 1) 在6份楊樹種質花粉中,36號楊、南楊和鉆天楊花粉活力較高,其次為110楊和小葉楊,森海2號楊活力最差; 24 h時花粉萌發率最大的種質是36號楊,高達74.42%(±2.36%),而三倍體森海2號楊花粉萌發率為0; 鉆天楊花粉管平均長度為456.00 μm(±2.05 μm),小葉楊花粉管最短,為276.44 μm(±11.08 μm),且不同種質的花粉萌發率和花粉管萌發長度差異極顯著(P<0.01),表明不同種質的花粉活力存在很大差異。2) 利用Imagemaster 2D platinum 6.0軟件分析南楊、110楊、森海2號楊、小葉楊和鉆天楊的2-DE圖譜,共發現有408個蛋白點相匹配,以36號楊鑒定的28個潛在致敏蛋白為參照,在5份種質花粉中有6個致敏蛋白均缺失表達,南楊和森海2號楊花粉有21個致敏蛋白,110楊次之,有19個,而小葉楊和鉆天楊的致敏蛋白數量最少,為18個。對其余22個致敏蛋白進行差異表達量分析發現,其中有12個致敏蛋白在5份楊樹種質中均低于36號楊,其相對表達量范圍為-3.99~-0.02,尤其是森海2號楊和小葉楊分別有10個和8個致敏蛋白表達量顯著(P<0.05)或極顯著(P<0.01)低于36號楊; 南楊和鉆天楊降低趨勢最小。有3個致敏蛋白在5份種質中表達均高于36號楊,其中南楊最高,達到極顯著水平(P<0.01),其次為鉆天楊、森海2號楊和110楊、小葉楊。組間差異方差分析發現南楊和鉆天楊的17個致敏蛋白表達量顯著或極顯著大于小葉楊和森海2號楊。3) qRT-PCR分析結果表明,9個致敏蛋白編碼基因中有5個在南楊和鉆天楊中表達量都較高,包括spots 12、51、200、216和161,其次是小葉楊,而森海2號楊和110楊均只有2個致敏基因表達高。【結論】 楊樹不同種質間花粉活力、致敏蛋白數量以及表達量存在明顯差異,可為今后楊樹低致敏新品種選育提供重要的參考,但是成熟花粉致敏蛋白表達量的高低與其致敏性強弱的相關性還有待進一步深入研究。
楊樹; 成熟花粉; 致敏蛋白; 雙向電泳; 實時熒光定量PCR
近年來,過敏性疾病已經成為許多國家和地區季節性的流行病(呂相征等, 2005),受到社會各界的廣泛關注,被世界衛生組織列為21世紀重點防治的三大疾病之一。在眾多的過敏原之中,花粉是自然界中最常見的吸入型過敏原,容易造成過敏患者出現支氣管哮喘、季節性鼻炎、枯草熱及皮炎等癥狀(陳紹寧等, 2008)。新的統計結果表明,幾乎所有工業化國家的花粉過敏病人口都在劇增,例如美國、日本、意大利、西班牙、奧地利等國家花粉過敏癥的人數占總人口的15%左右,而挪威、瑞典、比利時、英國和法國花粉過敏患者高達30%左右。在我國,花粉過敏的發病率為0.5%~1%,高發病區達5%,雖然沒有上述一些國家那么嚴重,但近年來發病率也呈現逐年增高趨勢(楊瓊梁等, 2015)。
木本植物花粉過敏現象在許多國家都有報道,花粉飄散具有明顯的季節性和地域性,不同國家和地區在不同季節有不同的致敏花粉。Lin等(2002)調查了引起花粉過敏癥主要的過敏原,結果發現引起過敏(患病率)的花粉樹種主要是美國白櫟(Quercusalba)(34.3%)、垂枝樺(Betulapendula)(32.9%)和梣葉槭(Acernegundo)(32.9%),其次為山毛櫸(Fagusgrandifolia)(29.6%)、美國白梣(Fraxinusamericana)(26.0%)、美國榆(Ulmusamericana)(24.6%)和美洲黑楊(Populusdeltoides)(20.6%)。根據土耳其錫夫里希薩爾氣傳致敏花粉濃度的調查結果顯示,松科(Pinaceae)、柏科(Cupressaceae)、白蠟樹屬(Fraxinus)、楊屬(Populus)是主要的氣傳致敏花粉(Erkara, 2008)。Mardones 等(2013)研究了塔卡爾2007年5月—2008年4月期間大氣花粉粒數,發現空氣中花粉數量最多的樹種是二球懸鈴木(Platanusacerifolia),其花粉濃度達到每周平均每日高達203粒·m-3; 其次是埃及榕(Acerpseudoplatanus)(116粒·m-3),楊屬(103粒·m-3)和木犀欖(Oleaeuropaea)(19粒·m-3)相對較少。在我國,由于南北地理、海拔、氣溫等差異大,花粉致敏種類不盡相同。我國南方花粉致敏樹種主要有懸鈴木屬(Platanus)、松屬(Pinus)、構屬(Broussonetia)等,而北方主要以懸鈴木屬、楊屬、柳屬(Salix)、樺木屬(Betula)為代表(辛嘉楠等, 2007; 楊瓊梁等, 2015)。花粉能引起人過敏是因為花粉中含有能剌激機體發生過敏反應的物質,其主要成分是蛋白質。迄今為止,關于木本植物花粉過敏原的研究報道主要有懸鈴木屬(Pla a 1和Pla a 2)(Asturiasetal., 2002; Ibarrolaetal., 2004)、樺木屬(Bet v 1,Bet v 2和Bet v 4等)(Ebneretal., 1995; Twardoszetal., 1997)、榆屬(Ulmus)(Car b 1)(Hiraharaetal., 2001)、日本柳杉(Cryptomeriajaponica)(Cry j 1,Cry j 2和Jun a 1)(Larsenetal., 1992; Midoroetal., 1999)等。
我國是世界上楊樹人工林面積最大的國家,面積已超666.7萬hm2(賈黎明等, 2013)。楊樹不僅是速生豐產林、防護林的主要造林樹種,也是我國城市綠化常用的樹種,楊樹的花期3—4月,雄株產生大量的花粉,具有過敏的潛能。為了解楊樹花粉致敏原,本實驗室建立了楊樹花粉蛋白提取和分離技術體系,并對美洲黑楊36號楊(P.deltoides‘2KEN8’)花粉進行了蛋白分離和致敏蛋白預測(Zhangetal., 2015),為研究其他楊樹種質花粉致敏性奠定了基礎。楊樹種類繁多,不同種質間的花粉致敏性可能存在差異。為了探索不同楊樹種質間花粉致敏蛋白的表達特性差異,本文選用目前生產上常見的6份楊樹種質為研究材料,開展了楊樹不同種質花粉萌發率、花粉管長度等基本特性研究; 并利用差異蛋白質組技術,以36號楊成熟花粉預測的28個致敏蛋白為基礎,對其他5份楊樹種質間花粉致敏蛋白的表達水平進行了比較,為探索楊樹不同種質的花粉致敏性差異、選育低致敏楊樹品種以及控制植物過敏性疾病等研究提供理論依據。
1 材料與方法
1.1 花粉收集與保存
6份楊樹種質[36號楊(Populusdeltoides‘2KEN8’)、南楊(P.deltoides‘Nanyang’)、110楊(P.deltoides×P.maximowiczii‘Eridano’)、森海2號楊(P.deltoides×P.cathayana‘Senhai 2’)、小葉楊(P.simonii)和鉆天楊(P.nigravar.italica)]花枝于2015年3月采自北京、河北和河南(表1)。將花枝適當修剪,在室內進行隔離水培,室溫(20±2) ℃。待雄花成熟散粉時,用培養皿收集花粉,室溫晾干后,裝于50 mL離心管中,在-20 ℃冰箱中保存。
1.2 花粉離體培養
采用液體培養法進行楊樹花粉萌發試驗(胡超, 2011),稍有改動。將花粉從-20 ℃冰箱中取出,每份楊樹種質稱取0.01 mg花粉,在4 ℃條件下充分吸水4 h。將花粉均勻撒入液體培養基(15%蔗糖 + 100 mg·L-1硼酸 + 300 mg·L-1CaCl2+ 200 mg·L-1MgSO4+ 100 mg·L-1KNO3,pH6.0)上,21 ℃暗培養2,4,8,12,24 h后,在光學顯微鏡下觀察花粉萌發情況,統計花粉萌發數并測量花粉管長度。每個處理隨機觀察5個視野,每個視野花粉粒不少于30個,以花粉管長度大于花粉粒直徑視為萌發。萌發率=已萌發花粉數/花粉總數 × 100%。每個樣品在不同培養時間隨機選取50個已萌發的花粉,采用顯微測微尺測量花粉管長度。每份種質設置3次生物學重復。
1.3 成熟花粉可溶性總蛋白的提取
采用三氯乙酸丙酮(TCA-丙酮)沉淀法提取5份楊樹種質成熟花粉可溶性總蛋白質(Zhangetal., 2015)。具體操作: 將花粉從-20 ℃冰箱中取出,稱取1 mg花粉,置于4 ℃條件下充分吸水4 h。加液氮充分研磨至粉末,置于100 mL預冷的TCA-丙酮溶液(10%TCA,0.07%β-巰基乙醇)中,震蕩混勻后在-20 ℃條件下靜置過夜,再14 000 r·min-1離心30 min,棄上清; 用冷丙酮(含0.07%β-巰基乙醇)沖洗沉淀1 h后,14 000 r·min-1離心30 min,重復上述步驟3~4次,直至沉淀物為白色; 真空凍干成干粉,-80 ℃保存備用。
1.4 成熟花粉可溶性總蛋白的雙向電泳
按照1 mg·mL-1的比例用蛋白質裂解緩沖液[7 mol·L-1尿素,65 mmol·L-1DTT,4%(W/V)CHAPS,0.01%PMSF,0.2%載體兩性電解質]溶解凍存的蛋白樣品,并用Bradford法測定蛋白濃度。取600 μg樣品與水化液(7 mol·L-1尿素,2 mol·L-1硫脲,4%CHAPS,65 mmol·L-1DTT,0.002 kg·L-1Bio-Lyte)充分混合后作為上樣樣品,總體積為350 μL。首先,使用非線性IPG固相膠條進行第1向等電聚膠電泳。電泳結束后,立即將IPG膠條依次于10 mL平衡液Ⅰ[6 mol·L-1尿素,0.02 kg·L-1SDS,0.375 mol·L-1Tris-HCl(pH8.8),30%甘油,0.02 kg·L-1DTT]和平衡液Ⅱ[6 mol·L-1尿素,0.021 kg·L-1SDS,0.375 mol·L-1Tris-HCl(pH8.8), 20%甘油,0.04 kg·L-1碘乙酰胺]中各平衡15 min。然后進行第2向電泳,待溴酚藍線至凝膠底部邊緣時停止電泳,將SDS-PAGE膠在固定液中(10%乙酸,40%甲醇,50%ddH2O)震蕩過夜,再轉移至考馬斯亮藍G-250染色液(0.1%G-250,25%異丙醇,10%冰醋酸)中震蕩染色13 h。 最后用ddH2O脫色至背景近無色。每個品種的雙向電泳試驗平行重復3次,以確保試驗結果的可靠性。
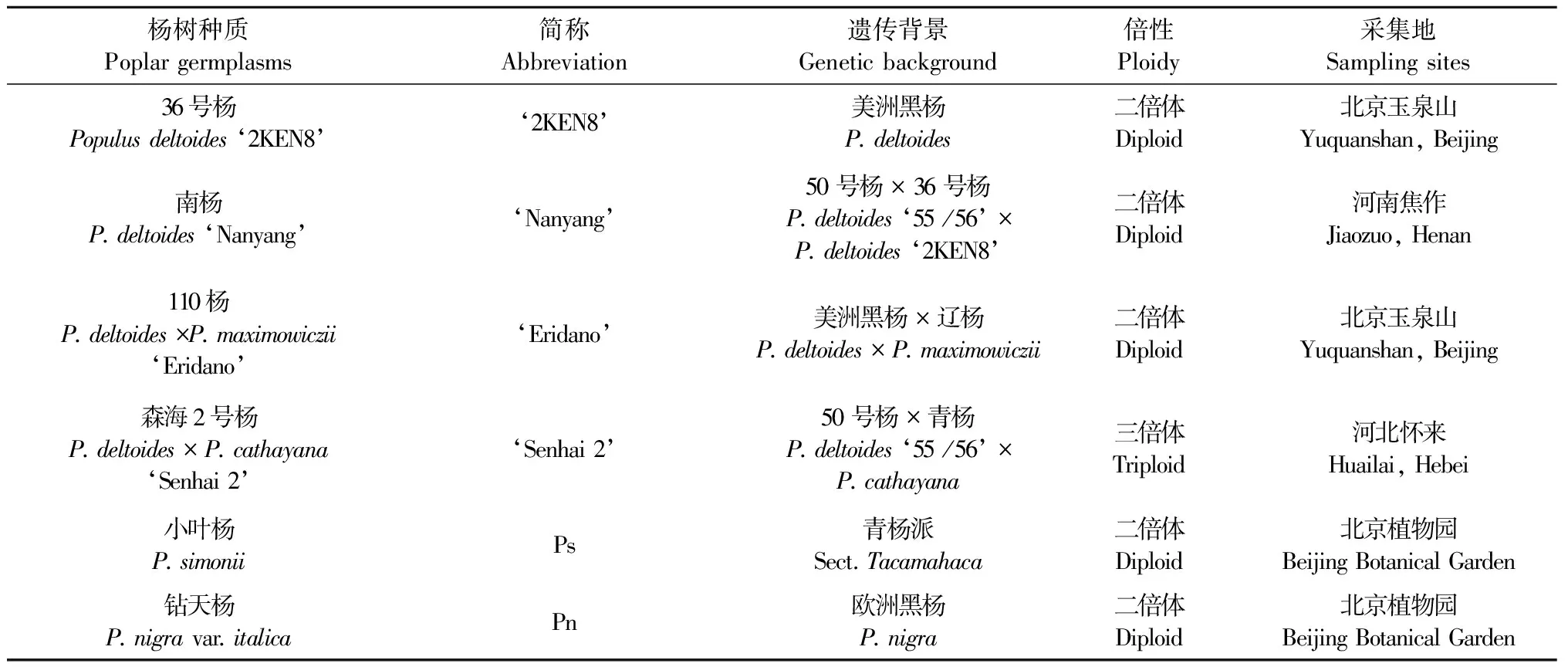
表1 試驗材料基本情況Tab. 1 The background of experimental materials
1.5 實時熒光定量PCR(qRT-PCR)
楊樹花粉總RNA提取方法參照植物總RNA提取試劑盒(RNeasy Plant Kit)使用說明書,將提取的RNA純化后進行反轉錄成cDNA(TaKaRa,China),以cDNA為模板進行qRT-PCR。利用在線設計軟件Primer3 Input 0.4.0設計實時定量PCR引物,其引物序列見表2。參照SYBR? Premix Ex TaqTMKit(TaKaRa,China)配制PCR 反應體系,反應在Roche480(Roche,USA)上進行,程序為: 95 ℃ 30 s; 95 ℃ 5 s,60 ℃ 30 s,40個循環; 60 ℃ 1 min; 50 ℃ 30 s。以PtActin作為內參對照,以36號楊的致敏相關基因作為對照組,qRT-PCR數據分析參照采用2-ΔΔCt方法(Pfaffl, 2001)。為了確保試驗結果的真實性和可靠性,每個試驗均設3次生物學重復和4次技術重復。

表2 用于qRT-PCR分析致敏蛋白基因的引物Tab. 2 Primers used in RT-PCR analysis of the allergenic proteins
1.6 分析方法
試驗數據采用SPSS 13.0和Originlab 9.0軟件進行方差分析、顯著性分析和圖表分析等。
2 結果與分析
2.1 花粉萌發率及花粉管長度差異分析
通過光學顯微鏡分別觀察6份楊樹種質成熟花粉在暗培養2,4,8,12和24 h后的萌發動態,統計萌發花粉的數量,并測量花粉管長度。當培養到2 h時,花粉粒已經吸水膨大,部分花粉粒的花粉管已經伸出,隨著培養時間的延長,花粉萌發率和花粉管長度逐漸提高(圖1A)。萌發試驗結果顯示,除森海2號楊花粉一直沒有萌發外,其他5份楊樹種質的花粉萌發率均隨著培養時間的延長逐漸提高。36號楊和南楊花粉萌發較快,在培養2 h時,萌發率分別為29.12%(±0.16%)和21.44%(±0.25%); 在培養4 h時,鉆天楊花粉萌發率迅速增加,達到36.67%(±0.02%); 在培養8 h時,110楊花粉仍未萌發; 在培養12 h時,小葉楊和110楊花粉萌發率迅速增加到41.74%(±1.51%)和38.83%(±0.16%); 在培養24 h時,36號楊花粉萌發率最大,高達74.42%(±2.36%),其他依次是南楊67.23%(±2.10%)、鉆天楊62.54%(±0.01%)、小葉楊56%(±0.04%)和110楊42.23%(±1.95%)。差異分析結果顯示,不同楊樹種質的花粉萌發率差異達到了極顯著水平(P<0.01)(圖1B)。由圖1C可看出,楊樹花粉管平均長度隨著培養時間的延長呈現逐漸增加趨勢。培養24 h時,鉆天楊花粉管平均長度為456.00 μm(±2.05 μm),其他依次為南楊410.10 μm(±11.31 μm),110楊390.06 μm(±12.59 μm),36號楊310.16 μm(±11.13 μm)和小葉楊276.44 μm(±11.08 μm),而且不同楊樹種質的花粉管萌發長度差異也達到了極顯著水平(P<0.01)。以上結果表明,36號楊、南楊和鉆天楊花粉活力較高,其次為110楊和小葉楊,森海2號楊活力最差,且不同楊樹種質的成熟花粉活力存在明顯差異。
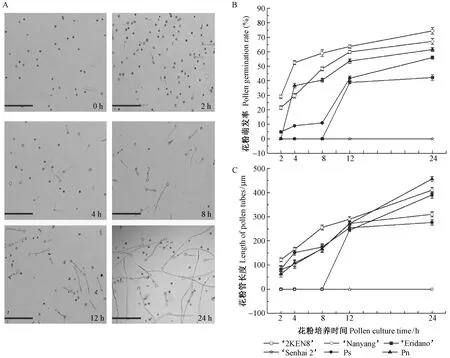
圖1 不同培養時間下6份楊樹種質花粉萌發動態、萌發率和花粉管長度比較(標尺:200 μm)Fig.1 Change of the pollen germination, tube length with time for culturing in six poplar germplasms(Bar:200 μm)A. 花粉萌發0,2,4,8,12,24 h的萌發狀態; B. 6份楊樹種質的花粉萌發率; C. 6份楊樹種質的花粉管長度隨培養時間的變化。A. Pollen germinating state at 0, 2, 4, 8, 12 and 24 h respectively; B. Pollen germination rate of six poplar germplasms; C. Change of pollen tube length with culturing time of poplar germplasms.
2.2 楊樹成熟花粉蛋白的雙向電泳圖譜分析
南楊、110楊、森海2號楊、小葉楊和鉆天楊成熟花粉蛋白的2-DE電泳圖譜(圖2)十分相似,表明該5份楊樹種質之間可以進行差異蛋白表達量的分析。利用Imagemaster 2D Platinum 6.0軟件對5份楊樹種質花粉的2-DE圖譜進行比較分析,在南楊、110楊、森海2號楊、小葉楊和鉆天楊的2-DE圖譜上分別檢測到449,442,428,419和422個蛋白質點,5份楊樹種質間共有408個蛋白點相匹配,這些蛋白質的分子質量范圍為5~63 kDa,等電點為4~7。但是,5份楊樹種質成熟花粉卻未發現特異蛋白。

圖2 5份楊樹種質成熟花粉2-DE圖譜Fig.2 2-DE profile of mature pollen of five poplar germplasms
2.3 致敏蛋白在不同楊樹種質的表達差異分析
以36號楊已鑒定的28個潛在致敏蛋白(Zhangetal., 2015)為參照,對其他5份楊樹種質成熟花粉的致敏蛋白相對表達量進行分析。從表3中可看出,南楊有22個致敏蛋白,森海2號楊有21個致敏蛋白,110楊有20個致敏蛋白,小葉楊有19個致敏蛋白,鉆天楊的致敏蛋白數量最少,為18個。與36號楊中的28個潛在致敏蛋白相比,有6個致敏蛋白點(spot 22,66,84,142,144,184)在該5份種質成熟花粉中均不存在,其他22個致敏蛋白點在電泳圖譜中的放大圖如圖3所示。隨后,對該22個致敏蛋白進行差異表達量分析,其中有12個致敏蛋白(spot 3,5,78,95,117,135,156,164,181, 197, 200, 203)在5份楊樹種質中表達量較低,其相對表達量范圍為-3.99~-0.02,其中森海2號楊和小葉楊表達量最低,其平均相對表達量分別為-1.83和-1.98。差異分析顯示,森海2號楊和小葉楊分別有10個和8個致敏蛋白顯著(P<0.05)或極顯著(P<0.01)低于36號楊。南楊和鉆天楊表達量稍低,分別為-0.83和-0.54。有3個致敏蛋白(spot 133, 208,216)在5份種質間表達量均較高。其中南楊最高,其平均相對表達量為2.88,均極顯著大于36號楊(P<0.01); 其次為鉆天楊(2.60)、森海2號楊(1.19)、110楊(0.83)和小葉楊(0.81)。組間差異方差分析發現,南楊和鉆天楊的17個致敏蛋白表達量均顯著(P<0.05)或極顯著(P<0.01)高于小葉楊和森海2號楊,例如肌球蛋白抑制蛋白4(spot 216)在南楊的表達量高達小葉楊的8倍之多。
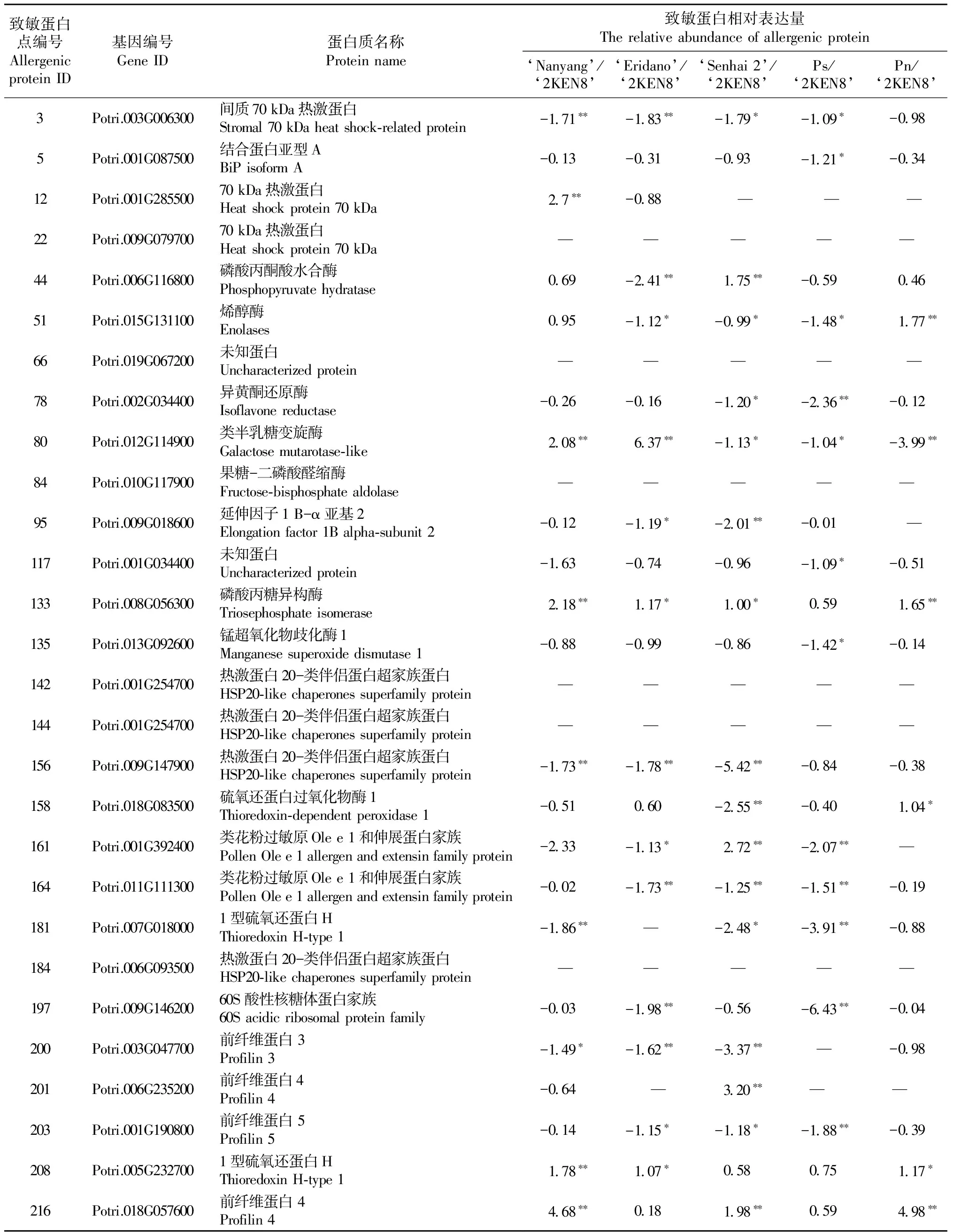
表3 致敏蛋白在不同楊樹種質間的表達量差異分析①Tab. 3 The different expression of allergic protein among different poplar germplasms
① “—”代表該種質花粉沒有這個致敏蛋白點; “*”,“**”分別代表對照組與試驗組之間差異顯著(P<0.05)和極顯著(P<0.01)。“—”represents the germplasm without this allergenic protein; “*”, “**” represent significant (P<0.05) and highly significant (P<0.01)difference between control and treatment, respectively.

圖3 5份楊樹種質的致敏蛋白點的放大Fig.2 Enlargement view of allergenic proteins in five poplar germplasms
2.4 致敏蛋白編碼基因在楊樹花粉中的轉錄水平
為進一步了解致敏蛋白編碼基因在6份楊樹種質間的轉錄水平,隨機選取9個致敏蛋白基因,通過qRT-PCR對這9個基因的轉錄水平進行分析(圖4)。結果顯示,spot 12,51, 200和216蛋白編碼基因在南楊和鉆天楊中的表達量顯著(P<0.05)或極顯著高于其他種質; spot 44和spot 197蛋白編碼基因在森海2號楊、小葉楊和鉆天楊中的表達量極顯著高于其他種質; spot 161蛋白編碼基因在南楊、森海2號楊、小葉楊和鉆天楊花粉中的表達量均是對照組36號楊的4 700倍以上,在鉆天楊花粉中的表達量竟高達8 453.06(±267.47)倍; spot 5蛋白編碼基因僅在小葉楊中表達相對較高,達到顯著水平; spot 80蛋白編碼基因在5份種質花粉中的表達差異不顯著。綜合以上分析表明,9個致敏蛋白編碼基因中,大多在南楊和鉆天楊中表達量較高,而在森海2號楊和小葉楊中表達量較低,這與前面致敏蛋白在5份種質間的表達模式相一致。因此,差異定量結果分析也進一步驗證了差異蛋白質組分析結果。
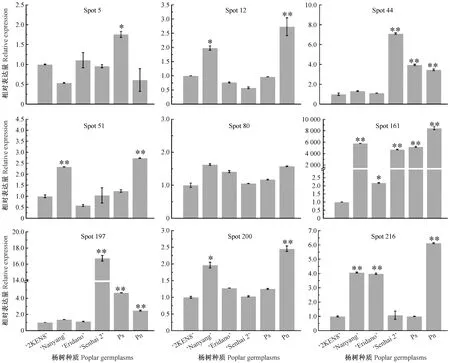
圖4 qRT-PCR分析致敏相關基因在楊樹成熟花粉中的表達情況Fig.4 Expression of related allergenic genes in poplar mature pollen by qRT-PCR “*”和“**”分別代表對照組與試驗組之間差異顯著(P<0.05)和極顯著(P<0.01)。 “*” and “**” represent significant (P<0.05) and highly significant (P<0.01) difference between control and treatment, respectively.
3 討論
過敏原作為花粉的激發物質,分析其在不同楊樹種質的差異對于低致敏品種選育具有重要意義。從本研究中的6份楊樹種質成熟花粉致敏蛋白數量、表達量及致敏蛋白編碼基因表達量來看,南楊和鉆天楊的致敏蛋白表達量顯著或極顯著大于其他楊樹種質。其致敏蛋白主要涉及蛋白質的合成與加工(spot 3,5,95,156, 197)、信號轉導(spot 200, 201和203)、糖酵解(spot 44,51和133)以及防御或脅迫反應(spot 78,135和208),這些蛋白為植物花粉的發育提供物質基礎和能量保障,促進花粉的萌發,與花粉萌發試驗結果相符。Ole e 1是木犀欖花粉主要過敏原,90%木犀欖花粉過敏患者的過敏原都是Ole e 1(Quiralteetal., 2002),其同源成員被鑒定為許多植物花粉的過敏原,如木樨科(Oleaceae)[如歐梣(Fraxinusexcelsior)、紫丁香(Syringaoblata)和歐洲女貞(Ligustrumvulgare)等]、平車前(Plantagodepressa)、藜(Chenopodiumalbum)等,但Ole e 1家族蛋白具體的功能和作用機制目前尚不清楚(Lombarderoetal., 2002)。在本研究中,Ole e 1家族蛋白(spot 161)在南楊、森海2號楊、小葉楊和鉆天楊花粉中的表達量均是對照組36號楊的4 700倍以上,特別是在鉆天楊花粉的表達量竟高達8 453.06(±267.47)倍,故推測Ole e 1可能是楊樹花粉的主要過敏原。此外,有6個致敏蛋白在5份種質花粉中均沒有檢測到表達情況,這可能是由于其表達量相對較低,雙向電泳技術很難檢測出來。
三倍體植物具有產量高和抗性強的雙重優勢,但是其花粉育性低(楊詩勤, 2014)。森海2號楊是通過人工雜交選育出的抗性強的優良三倍體新品種(賈會霞等, 2015)。在本研究中,森海2號楊的花粉沒有萌發,可能由于多倍體小孢子母細胞發育異常造成花粉活力差和育性低(曹媛等, 2012),而且其致敏蛋白表達水平也顯著低于其他種質。Kondo等(1997)發現三倍體日本柳杉(Cryptomeriajaponica)花粉過敏原Cry j 1和Cry j 2含量低于二倍體,認為三倍體植物可能具有低致敏性,可以考慮作為低致敏楊樹品種選育的一個方面。目前,對于三倍體花粉的低育性、低致敏性的機制還不清楚,有待于進一步深入研究。而且,本研究只是比較了楊樹不同種質的成熟花粉致敏蛋白差異,未對花粉萌發的不同階段進行差異蛋白質組分析。相關研究表明,花粉萌發伴隨著表達基因數量的降低及特異性轉錄本的增加,同時也會導致蛋白質組的變化(趙麗娟, 2011)。因此,花粉萌發可能會丟失或產生過敏蛋白,值得進一步研究。
本研究表明,不同楊樹種質間致敏蛋白表達量和致敏性存在差異,但問題是,致敏蛋白表達量的高低和致敏蛋白基因的表達量的多少與致敏性強弱有直接關聯嗎?Son等(1999)認為蘋果(Malus×domestica)不同品種間的致敏性差異主要與過敏原的表達量差異有關。Bolhaar等(2005)通過酶聯免疫測定金冠蘋果的Mald 1(蘋果主要過敏原)含量遠低于‘Jonathan’品種,但前者的Mald 1同患者血清中的抗體(sIgE)結合能力要強于后者。Zuidmeer等(2006)通過體外免疫測定方法,進一步研究了造成這種結果沖突的原因,認為可能是過敏原結構的不穩定,難以控制。本研究初步探索了5份楊樹種質成熟花粉致敏蛋白的表達特性,在今后的研究中,可以結合 IgE 檢測、組胺釋放試驗(HAT)、酶聯免疫吸附試驗(ELISA)等技術測定過敏原對患者血清中抗體結合能力的強弱來驗證致敏蛋白的表達量與致敏性的相關性以及確定低致敏性楊樹種質(Sichereretal., 2010)。同時,還可以借助現代分子生物技術如RNA干擾進行基因沉默,以達到消除致敏蛋白的目的(Galloetal., 2009)。
4 結論
本研究通過楊樹成熟花粉萌發試驗、差異蛋白質組、qRT-PCR技術對不同楊樹種質花粉中致敏蛋白表達量的差異進行了分析,結果表明楊樹不同種質的花粉活力、致敏蛋白表達水平存在顯著差異。其中36號楊、南楊和鉆天楊花粉活力較高; 南楊和森海2號楊的致敏蛋白數量最多,小葉楊和鉆天楊的致敏蛋白數量最少,且22個致敏蛋白在6份楊樹種質的表達存在很大差異,南楊和鉆天楊致敏蛋白表達量極顯著高于其他種質。該結果對今后篩選低致敏楊樹品種及控制植物過敏性疾病等研究具有重要參考價值。
曹 媛,康向陽,張志毅,等. 2012. 毛白楊花粉敗育過程中Ca2+-ATPase 的異常變化. 北京林業大學學報,34 (4): 10-17.
(Cao Y,Kang X Y,Zhang Z Y,etal. 2012. Anomalous changes in Ca2+-ATPase distribution during the process of pollen abortion inPopulustomentosaCarr. Journal of Beijing Forestry University,34(4): 10-17. [in Chinese])
陳紹寧,王 偉,葉永忠. 2008. I型花粉致敏原的分子特性及其生物學功能. 植物生理學通訊,44 (2): 341-344.
(Chen S N,Wang W,Ye Y Z. 2008. Molecular characteristics and biological function of group-1 pollen allergens. Plant Physiology Communications,44(2): 341-344. [in Chinese])
胡 超. 2011. 歐美黑楊與小葉楊雜交后代性狀分析及胚培養研究.武漢: 華中農業大學碩士學位論文.
(Hu C. 2011. Researches on offspring’s characters and embryo culture of hybrids betweenPopuluseuramericanaandPopulussimonii. Wuhan: MS thesis of Huazhong Agricultural University. [in Chinese])
賈會霞,姬慧娟,胡建軍,等. 2015. 楊樹新品種的SSR指紋圖譜構建和倍性檢測. 林業科學,51 (2): 70-79.
(Jia H X,Ji H J,Hu J J,etal. 2015. Fingerprints of SSR markers and ploidy detection for newPopulusvarieties. Scientia Silvae Sinicae,51(2): 70-79. [in Chinese])
賈黎明,劉詩琦,祝令輝,等. 2013. 我國楊樹林的碳儲量和碳密度. 南京林業大學學報:自然科學版,37 (2): 1-7.
(Jia L M,Liu S Q,Zhu L H,etal. 2013. Carbon storage and density of poplars in China. Journal of Nanjing Forestry University: Natural Sciences,37(2): 1-7. [in Chinese])
呂相征,劉秀梅,楊曉光. 2005. 健康人群食物過敏狀況的初步調查. 中國食品衛生雜志,17 (2): 119-121.
(Lü X Z,Liu X M,Yang X G. 2005. Preliminary survey on status of food allergy in young Chinese students. Chinese Journal of Food Hygiene,17(2): 119-121. [in Chinese])
辛嘉楠,歐陽志云,鄭 華,等. 2007. 城市中的花粉致敏植物及其影響因素. 生態學報,27 (9): 3820-3826.
(Xin J N,Ouyang Z Y,Zheng H,etal. 2007. Allergenic pollen plants and their influencing factors in urban area. Acta Ecologica Sinica,27(9): 3820-3826. [in Chinese])
楊瓊梁,歐陽婷,顏 紅,等. 2015. 花粉過敏的研究進展. 中國農學通報,31 (24): 163-164.
(Yang Q L, Ouyang T, Yan H,etal. 2015. Research progress of pollen allergy. Chinese Agricultural Science Bulletin,31(2): 163-164. [in Chinese])
楊詩勤. 2014. 水稻減數分裂相關基因OsPHS1的轉化與鑒定. 武漢: 湖北大學碩士學位論文.
(Yang S Q. 2014. Transformation and identification ofOsPHS1 gene related to rice meiosis. Wuhan: MS thesis of Hubei University. [in Chinese])
趙麗娟. 2011. 小黑楊(Populussimonii×P.nigra)花粉功能基因組和蛋白質組研究. 哈爾濱: 東北林業大學博士學位論文.
(Zhao L J. 2011. Functional genomics and proteomics ofPopulussimonii×P.nigrapollen. Harbin: PhD thesis of Northeast Forestry University. [in Chinese])
Asturias J,Ibarrola I,Bartolome B,etal. 2002. Purification and characterization of Pla a 1, a major allergen fromPlatanusacerifoliapollen. Allergy,57(3): 221-227.
Bolhaar S T H P,van de Weg W E,van Ree R,etal. 2005. In vivo assessment with prick-to-prick testing and double-blind, placebo-controlled food challenge of allergenicity of apple cultivars. Journal of Allergy and Clinical Immunology, 116 (5): 1080-1086.
Ebner C,Hirschwehr R,Bauer L,etal. 1995. Identification of allergens in fruits and vegetables: IgE cross-reactivities with the important birch pollen allergens Bet v 1 and Bet v 2 (birch profilin). Journal of Allergy and Clinical Immunology,95(5): 962-969.
Erkara I P. 2008. Concentrations of airborne pollen grains in Sivrihisar (Eskisehir), Turkey. Environmental Monitoring and Assessment,138 (1-3): 81-91.
Gallo M,Richard S. 2009. Removing allergens and reducing toxins from food crops. Current Opinion in Biotechnology, 20 (2): 191-196.
Hirahara K,Tatsuta T,Takatori T,etal. 2001. Preclinical evaluation of an immunotherapeutic peptide comprising 7 T-cell determinants of Cry j 1 and Cry j 2, the major Japanese cedar pollen allergens. Journal of Allergy and Clinical Immunology,108(1): 94-100.
Ibarrola I,Arilla M,Martínez A,etal. 2004. Identification of a polygalacturonase as a major allergen (Pla a 2) fromPlatanusacerifoliapollen. Journal of Allergy and Clinical Immunology,113 (6): 1185-1191.
Kondo Y,Ipsen H,L?wenstein H,etal. 1997. Comparison of concentrations of Cry j 1 and Cry j 2 in diploid and triploid Japanese cedar (Cryptomeriajaponica) pollen extracts. Allergy,52(4): 455-459.
Larsen J N,Str?man P,Ipsen H. 1992. PCR based cloning and sequencing of isogenes encoding the tree pollen major allergen Car b I fromCarpinusbetulus, hornbeam. Molecular Immunology,29(6): 703-711.
Lin R Y,Clauss A E,Bennett E S. 2002. Hypersensitivity to common tree pollens in New York City patients. Allergy and Asthma Proceedings,23 (4): 253-258.
Lombardero M,Obispo T,Calabozo B,etal. 2002. Cross-reactivity between olive and other species. Role of Ole e 1-related proteins. Allergy,57(s71): 29-34.
Mardones P,Grau M,Araya J,etal. 2013. First annual register of allergenic pollen in Talca, Chile. Allergologia Et Immunopathologia,41 (4): 233-238.
Midoro H T,Goldblum R M,Kurosky A,etal. 1999. Isolation and characterization of the mountain cedarJuniperusasheipollen major allergen, Jun a 1. Journal of Allergy and Clinical Immunology,104 (3): 608-612.
Pfaffl M W. 2001. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Research,29(9): 45.
Quiralte J,Florido F,Arias J,etal. 2002. Olive allergen-specific IgE responses in patients withOleaeuropaeapollinosis. Allergy,57(s71): 47-52.
Sicherer S H,Sampson H A. 2010. Food allergy. Journal of Allergy and Clinical Immunology,125 (2): 116-125.
Son D Y,Scheurer S,Hoffmann A,etal. 1999. Pollen-related food allergy: cloning and immunological analysis of isoforms and mutants of Mal d 1, the major apple allergen, and Bet v 1, the major birch pollen allergen. European Journal of Nutrition,38 (4): 201-215.
Twardosz A,Hayek B,Seiberler S,etal. 1997. Molecular characterization, expression inEscherichiacoli, and epitope analysis of a two EF-hand calcium-binding birch pollen allergen, Bet v 4. Biochemical and Biophysical Research Communications,239(1): 197-204.
Zhang J,Wu L S,Fan W,etal. 2015. Proteomic analysis and candidate allergenic proteins inPopulusdeltoidescl. “2KEN8” mature pollen. Frontiers in Plant Science,6: 1-22.
Zuidmeer L,van Leeuwen W A,Kleine B I,etal. 2006. Allergenicity assessment of apple cultivars: hurdles in quantifying labile fruit allergens. International Archives of Allergy and Immunology,41 (3): 230-240.
(責任編輯 徐 紅)
Differences in Pollen Germination and Allergenic Proteins among DifferentPopulusGermplasms
Zhang Xiaoling1Wang Liuqiang1Sun Pei1Wu Lishuan2Fan Wei1Zhang Jin1Lu Mengzhu1Hu Jianjun1
(1.StateKeyLaboratoryofTreeGeneticsandBreedingKeyLaboratoryofTreeBreedingandCultivationoftheStateForestryAdministrationResearchInstituteofForestry,CAFBeijing100091; 2.KeyLaboratoryofPhotobiology,InstituteofBotany,ChineseAcademyofSciencesBeijing100093)
【Objective】 The characteristics of pollen germination and the differences of allergenic proteins among six poplar germplasms (Populusdeltoides‘2KEN8’,P.deltoides‘Nanyang’,P.deltoides×P.maximowiczii‘Eridano’,P.deltoides×P.cathayana‘Senhai 2’,P.simoniiandP.nigravar.italica) were studied to provide a theoretical basis and scientific evidence for breeding hypoallergenic poplar varieties and for the control of plant allergy. 【Method】 Pollen was collected from flower branches of six poplar germplasms collected from Beijing, Hebei and Henan in March 2015 for a period of hydroponic culture, pollen germination was studied by liquid culture method, and pollen allergenic proteins of different poplar germplasms was used by two-dimensional electrophoresis and real-time quantitative PCR. 【Result】 1) The pollen viability ofP.deltoides‘2KEN8’,P.deltoides‘Nanyang’ andP.nigravar.italicawas higher, followed byP.simoniiandP.deltoides×P.maximowiczii‘Eridano’, however, the viability ofP.deltoides×P.cathayana‘Senhai 2’ was the worst; the largest pollen germination rate was inP.deltoides‘2KEN8’, up to 74.42% (±2.36%), while the pollen germination of triploidP.deltoides×P.cathayana‘Senhai 2’ was 0; the average length of pollen tube ofP.nigravar.italicawas 456 μm (±2.05 μm), the shortest was inP.simonii, 276.44 μm (±11.08 μm), and the difference of pollen germination rate and pollen tube length among different entities were highly significant (P<0.01), suggesting that the pollen viability was significantly different among different germplasms. 2) The 2-DE maps ofP.deltoides‘Nanyang’,P.deltoides×P.maximowiczii‘Eridano’,P.deltoides×P.cathayana‘Senhai 2’,P.simoniiandP.nigravar.italicamature pollen were analyzed by Imagemaster 2D platinum 6.0, a total of 408 protein spots were matched. Based on the 28 potentially allergenic proteins ofP.deltoides‘2KEN8’ identified in our previous study, 6 allergenic proteins were missing expression in the other five entities; bothP.nigravar.italicaandP.deltoides‘Nanyang’ had 21 allergenic proteins,P.deltoides×P.maximowiczii‘Eridano’ had 19; whileP.deltoides×P.cathayana‘Senhai 2’ andP.deltoides‘2KEN8’ had the least number of allergenic proteins, only 18. Morover, the analysis of difference expression indicated that 12 of the 22 remaining allergenic proteins were less expressed in the five entities than inP.deltoides‘2KEN8’, the range of relative expression level was -3.99--0.02. The expression level ofP.deltoides×P.cathayana‘Senhai 2’ andP.simoniiwas the least, especially they had 10 and 8 allergenic proteins which were significantly or highly significant lower than that ofP.deltoides‘2KEN8’, respectively. The expression of 3 allergenic proteins in the 5 entities was higher than inP.deltoides‘2KEN8’. The strongest expression was inP.deltoides‘Nanyang’, significantly greater than inP.deltoides‘2KEN8’, followed byP.nigravar.italica,P.deltoides×P.cathayana‘Senhai 2’,P.deltoides×P.maximowiczii‘Eridano’ andP.simonii. Variance analysis among groups suggested that the expression of 17 allergenic proteins ofP.nigravar.italicaandP.deltoides‘Nanyang’ were significantly (P<0.05) or highly significantly (P<0.01) greater than that ofP.deltoides×P.cathayana‘Senhai 2’ andP.simonii. 3) qRT-PCR results showed that there were 5 of 9 allergenic proteins encoding genes highly expressed inP.nigravar.italicaandP.deltoides‘Nanyang’, including spot 12, 51, 200, 216 and 161, followed byP.simonii, while both ofP.deltoides×P.maximowiczii‘Eridano’ andP.deltoides×P.cathayana‘Senhai 2’ had only 2 highly expressed allergenic protein coding genes.【Conclusion】 The pollen viability, the number and expression levels of allergenic proteins were significantly different among the 6 germplasm entities, providing an important basis for future breeding of new poplar hypoallergenic varieties. However, further study is needed to understand the correlation between expression levels and allergenicity of the allergenic proteins of mature pollen.
Populus; mature pollen; allergenic protein; two-dimensional electrophoresis; real-time quantitative PCR
10.11707/j.1001-7488.20170207
2016-04-06;
2016-04-19。
“十二五”林業行業公益類重大專項(201304103)。
S718.43
A
1001-7488(2017)02-0054-11
*胡建軍為通訊作者。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現代語文(2016年21期)2016-05-25 13:13:50
