鎘脅迫對海菜花種子萌發及幼苗初期生理生化的影響
2017-03-21 11:31:46陳祖擁范菲菲官小云
江蘇農業科學 2016年11期
陳祖擁+范菲菲+官小云
摘要:通過營養液培養法,研究Cd2+對海菜花種子萌發及幼苗初期葉綠素含量、抗氧化酶活性及可溶性蛋白質含量的影響。結果表明,各濃度(0.1~2.0 mg/L)Cd2+處理抑制海菜花種子的萌發,但與對照無顯著差異(P>0.05);海菜花種子的發芽勢、發芽指數、活力指數整體隨Cd2+處理濃度的增大而下降,當Cd2+處理濃度較高時,與對照差異顯著(P<0.05)。Cd2+脅迫明顯降低海菜花葉片中葉綠素a、葉綠素b、類胡蘿卜素含量,且與對照差異顯著(P<0.05)。隨著Cd2+濃度增大,超氧化物歧化酶(SOD)活性、過氧化物酶(POD)活性、可溶性蛋白質含量先升高后降低;過氧化氫酶(CAT)活性呈持續降低趨勢,且與對照差異顯著(P<0.05)。隨Cd2+脅迫濃度增大,其對海菜花的毒害作用增強。
關鍵詞:鎘脅迫;海菜花;萌發;葉綠素;抗氧化酶系統;可溶性蛋白質;毒害
中圖分類號: X522 文獻標志碼: A
文章編號:1002-1302(2016)11-0482-03
在云貴高原分布有我國特有的三級重點保護高原淡水植物海菜花(Ottelia acuminata var. acuminata),其對水質要求很高,對水生生態系統具有重要的生態指示作用。由于人類活動的增強,工業“三廢”排放,農業化肥、農藥等的大量使用和城市垃圾的肆意堆放,使生態系統中的重金屬污染日益加劇[1],導致海菜花數量急劇減少,而重金屬對海菜花種子發芽的毒害作用還鮮見報道。本研究選用毒性較強,且在貴州省具有較高土壤背景值[2]的Cd2+為代表,研究重金屬Cd2+對海菜花種子萌發和幼苗生長初期生理生化的影響,以期初步揭示Cd2+對海菜花的生態毒理效應。
1 材料與方法
1.1 材料
供試海菜花種子采自貴州省貴陽市花溪區花溪水庫下游花溪河段,挑選籽粒飽滿、大小均勻的種子用于試驗。CdCl2·2.5H2O 購自國藥集團上海化學試劑公司,各分析純、生化純試劑都用超純水配制。
1.2 試驗設計
用CdCl2·2.5H2O(分析純)和1/10濃度Hoagland營養液,分別配制0、0.1、0.2、0.5、1.0、2.0 mg/L Cd2+處理液,分別將等體積的該處理液置于鋪有2層濾紙、直徑9 cm的培養皿中,分別挑選飽滿的海菜花種子各20粒平鋪于濾紙上,蓋好玻璃培養皿,置于溫度(25±1) ℃、光照度2 000 lx、14 h/d、濕度80%的人工氣候培養箱中進行萌發試驗,每個處理3次重復。每天添加適量的超純水補償蒸發的水分,保持濾紙濕潤但不淹水。以胚根露出種皮時記錄為發芽種子[3],培養14 d后測定葉綠素、類胡蘿卜素、可溶性蛋白質含量,同時測定超氧化物歧化酶(SOD)、過氧化物酶(POD)、過氧化氫酶(CAT)活性。
1.3 分析方法
1.3.1 發芽指標計算方法 發芽指標計算方法如下:
發芽率(GR)=14 d內正常發芽的種子數/供試種子數×100%;
發芽勢(GP)=萌發前7 d發芽數之和/供試種子數×100%;
發芽指數(GI)=∑(Gt/Dt)。
式中:Gt為第t天的發芽數;Dt為相應發芽時間。
活力指數=GI×S。
式中:S為幼苗平均總長度[4]。
1.3.2 葉綠素、類胡蘿卜素含量測定 取各處理幼苗,用80%丙酮溶液提取,用分光光度計測定提取液在665、649、470 nm處的吸光度[5]。
1.3.3 SOD、POD、CAT活性和可溶性蛋白質含量的測定 取各處理幼苗,加入少量0.1 mol/L磷酸緩沖液(pH值7.8),置冰浴研磨勻漿,4 ℃、12 000 r/min離心15 min,上清液用于酶活性和可溶性蛋白含量的測定。SOD活性的測定采用NBT光化還原法[5],POD活性的測定采用愈創木酚法[5],CAT活性的測定采用南京建成科技有限公司CAT試劑盒測定,可溶性蛋白質含量的測定采用考馬斯亮藍G-250法[6]。
1.4 數據處理與分析
用SPSS 13.0軟件對試驗數據進行統計分析,顯著性水平設置為0.05水平;用Origin 2015軟件繪制變化趨勢圖。
2 結果與分析
2.1 鎘脅迫對海菜花種子發芽的影響
如表1所示,在試驗濃度范圍內,Cd2+脅迫對海菜花種子發芽率具有一定抑制作用,但與對照無顯著差異(P>0.05);不同Cd2+脅迫下海菜花種子發芽勢、發芽指數、活力指數的變化趨勢類似,整體表現出隨Cd2+處理濃度的增大呈下降趨勢,當Cd2+處理濃度較高時,其發芽勢、發芽指數、活力指數與對照差異顯著(P<0.05)。
2.2 鎘脅迫對海菜花幼苗生長的影響
從表2可知,隨著Cd2+濃度增大,海菜花幼苗長度、根長逐漸減小,且與對照差異顯著(P<0.05);當Cd2+濃度達到2.0 mg/L時,海菜花平均苗長僅為對照的7.4%,根的生長被完全抑制,沒有明顯的根系生成。Cd2+濃度≤0.5 mg/L時,Cd2+對幼苗葉片數無顯著影響;當Cd2+濃度≥1.0 mg/L時,海菜花幼苗葉片數隨Cd2+離子濃度增大而顯著減少(P<0.05)。海菜花平均根數隨Cd2+濃度增大呈減少趨勢,當Cd2+為0.1 mg/L時,海菜花平均根數與對照無顯著差異;當Cd2+≥0.2 mg/L時,海菜花平均根數均與對照差異顯著(P<0.05)。由此說明,隨著Cd2+脅迫濃度增大,Cd2+對海菜花幼苗生長的抑制作用增強。
2.3 鎘脅迫對海菜花光合色素含量的影響
葉綠素含量在很大程度上反映了植物的光合作用能力,葉綠素與光合速率、營養狀況關系密切,逆境條件下,葉綠素含量是表征植物受傷害程度的指標之一。類胡蘿卜素是植物細胞中重要的輔助色素,在光合作用過程中能夠激發活性氧的清除,保護細胞器免受傷害[4]。如表3所示,在Cd2+濃度為0.1~2.0 mg/L時,海菜花幼苗中葉綠素a、葉綠素b、類胡蘿卜素含量隨Cd2+處理濃度增大而降低;當Cd2+濃度為 2.0 mg/L 時,葉綠素a、葉綠素b、類胡蘿卜素含量分別僅為對照的1.7%、6.6%、2.7%,可見高濃度Cd2+顯著抑制了葉綠素、類胡蘿卜素的合成。
2.4 鎘脅迫對海菜花幼苗中抗氧化酶系統的影響
SOD、POD、CAT等3種抗氧化酶活性與植物抵御外界脅迫能力密切相關。由表4可知,隨著Cd2+濃度增大,海菜花幼苗中SOD活性整體表現出先升高后降低的趨勢,Cd2+脅迫下的海菜花幼苗SOD活性高于對照(P<0.05)。當Cd2+濃度為0.5 mg/L時,SOD活性最大,為對照的8.78倍;當Cd2+濃度為2.0 mg/L時,SOD活性仍為對照的2.46倍。海菜花幼苗中POD活性變化規律與SOD相似,隨著Cd2+濃度增大,POD活性也呈現先升高后降低的趨勢,當Cd2+濃度為 0.5 mg/L 時,POD活性最大,為對照的2.29倍,與對照差異顯著(P<0.05);隨著Cd2+濃度的繼續增大,POD活性開始下降,當Cd2+濃度為2.0 mg/L時,其活性與對照無顯著差異。由此可見,SOD的耐受性比POD強。海菜花幼苗中CAT活性的變化規律與SOD、POD活性不同,隨Cd2+濃度增大而逐漸降低,且均與對照差異顯著(P<0.05)。當Cd2+濃度為2.0 mg/L時,海菜花幼苗的CAT活性被抑制了85.9%,由此可見,CAT對Cd2+脅迫的耐受性較差。
2.5 鎘脅迫對海菜花幼苗中可溶性蛋白質含量的影響
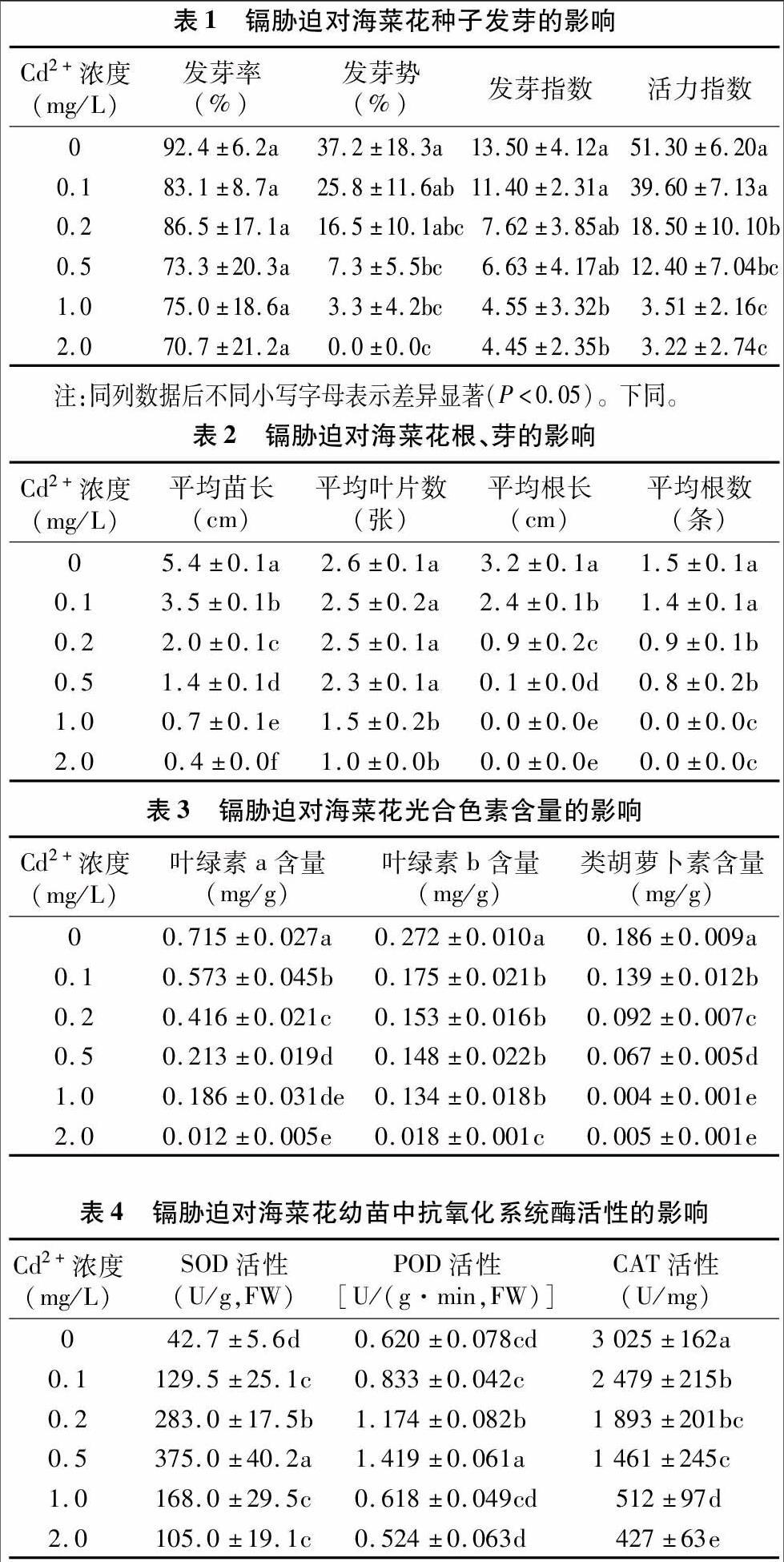
植物體內的可溶性蛋白質主要是參與各種代謝的酶類,其含量是植物的重要生理指標之一。由圖1可見,隨著Cd2+處理濃度增大,海菜花幼苗中可溶性蛋白質含量表現出先升高后降低的趨勢,在Cd2+處理濃度為0.5 mg/L時,海菜花幼苗中可溶性蛋白質含量最大,且與對照差異顯著(P<0.05);隨著Cd2+處理濃度的繼續增大,可溶性蛋白質含量開始下降,當Cd2+處理濃度為2.0 mg/L時,可溶性蛋白質含量與對照無顯著差異。由此表明,海菜花在受Cd2+脅迫時,體內能產生一些抗性蛋白抵御外界脅迫;但處于高濃度Cd2+脅迫時,這種抵御機制受到破壞,可溶性蛋白質含量降低。隨著鎘脅迫濃度增大,SOD、POD活性先升高后降低的變化趨勢也證明了可溶性蛋白質含量的這種變化規律。
3 結論與討論
本研究表明,各濃度Cd2+脅迫對海菜花種子的發芽率整體表現出抑制作用,這與Cd2+抑制紫羊茅[7]、白三葉[7]、亞麻[8]種子萌發的報道一致;而與低濃度Cd2+誘導、高濃度Cd2+抑制堿蓬[9]、黑麥草[10]等種子萌發的報道不同。種子萌發前期階段主要利用種子自身儲存的營養物質,外部污染物質須要通過種皮進入其內部才能參與萌發的前期過程。可能由于Cd2+在短期內較難通過海菜花種皮,所以對其發芽率抑制作用不顯著,這說明海菜花種子種皮的自我防御能力較強。海菜花種子的發芽勢、發芽指數、活力指數隨著外源Cd2+濃度的增大呈逐漸降低趨勢,在高濃度Cd2+處理時與對照差異顯著,這與Cd2+對紫羊茅[7]、白三葉[7]、黑麥草[10]種子萌發試驗結果一致。這說明高濃度Cd2+對海菜花種子萌發具有抑制作用,可能是高濃度Cd2+能部分通過種皮進入種子內部,抑制海菜花種子萌發。
植物幼苗生長和種子萌發過程中營養物質利用途徑不同,因此幼苗生長過程的Cd2+脅迫與發芽階段的Cd2+脅迫影響不同。Cd2+脅迫對海菜花幼苗的芽長、根長、葉片數、根數均有不同程度的影響,高濃度Cd2+脅迫對海菜花幼苗的芽長、根長、葉片數、根數有顯著的抑制作用,這與Cd2+對早熟禾[7]、紫羊茅[7]、白三葉[7]、亞麻[8]、黑麥草[10]影響的相關研究結果一致。由于重金屬對植物的傷害首先表現在對幼苗根部的傷害[11],所以Cd2+對根的抑制作用大于對芽的抑制作用。同時,由于胚根受到毒害,導致其對營養物質的吸收能力降低甚至消失,最終又影響芽的生長。另一方面,Cd2+可能使海菜花幼苗體內加速產生大量的活性氧,抑制各種酶活性,從而也影響到海菜花幼苗和根系的生長[12]。
重金屬破壞葉綠體結構,使植物葉片葉綠素含量降低,降低光合強度,這是重金屬對植物毒害的普遍現象[13]。本研究中,海菜花種子在受到Cd2+脅迫時,葉綠素a、葉綠素b、類胡蘿卜素含量均下降,這與Cd2+脅迫對黑麥草[10]、烤煙[14]綠葉素的影響規律一致。其主要原因是鎘抑制了葉綠素合成,降低海菜花幼苗中的葉綠素含量,減弱其光合作用,抑制其生長[10,13-14]。
SOD、POD、CAT是植物體內酶促防御系統的3個重要保護酶,它們發揮協同作用,消除活性氧,保護植物膜系統。本研究表明,隨著Cd2+處理濃度的增大,SOD、POD活性都是先升后降,這與Cd2+對黑麥草[10],Cu2+對海菜花幼苗[15]葉片中SOD、POD酶活性的影響規律一致。由此表明,低濃度Cd2+對POD、SOD活性具有激活作用,隨著Cd2+濃度逐漸增大,又轉為抑制作用。CAT活性則隨著Cd2+濃度增大而逐漸降低。由此可見,海菜花幼苗受Cd2+脅迫后抗氧化酶系統反饋機制不同,CAT、POD、SOD活性呈現不同的變化規律。陳國祥等認為,低濃度的重金屬逆境脅迫會抑制植物根系對養分、水分的吸收,同時引發活性氧(ROS)積累,增強脂質過氧化作用,使得SOD、POD活性增強[16]。本研究中,在低濃度Cd2+脅迫下,海菜花為有效緩解Cd2+脅迫引起的氧化脅迫,及時清除ROS的積累,SOD、POD活性增強也充分證實了此觀點。但隨著Cd2+濃度的進一步增大,高濃度的Cd2+導致海菜花幼苗中活性氧等自由基的積累超過植物本身的耐受極限,轉而開始抑制SOD、POD的活性。隨著Cd2+濃度增大,CAT活性逐漸降低,這可能是海菜花種子CAT對Cd2+敏感,不能抵御外來污染物脅迫,這也可能是其耐受性較差的原因之一。
海菜花幼苗中的可溶性蛋白是植物細胞內未與膜系統特異結合的酶,其含量越高,說明其體內生理生化反應與代謝活性越旺盛,有利于植物抵御逆境脅迫[12]。本研究中,隨著Cd2+濃度增大,可溶性蛋白質含量先升高后降低的變化趨勢,與Cd2+對谷子幼苗[12]、Cu2+對海菜花幼苗[15]葉片中可溶性蛋白質的影響結果一致。究其原因,一方面可能是適量的Cd2+可以誘導合成一些新蛋白質,參與抗逆性生理活動,促進可溶性蛋白質含量的增加;另一方面,高濃度Cd2+也誘導了蛋白水解酶活性增強,加速了蛋白質水解[12],同時高濃度Cd2+脅迫導致海菜花抗氧化酶系統受損,抗性酶減少,從而導致可溶性蛋白質含量減少。
參考文獻:
[1]王 磊,張 菲,王從彥,等. Cu、Cd污染背景下加拿大一枝黃花葉水提液對萵苣種子萌發和生長的生態毒理效應[J]. 生態毒理學報,2015,10(4):203-209.
[2]趙志鵬,邢 丹,劉鴻雁,等. 典型黃壤和石灰土對Cd的吸附解吸特性[J]. 貴州農業科學,2015,43(6):83-86.
[3]馬海鴿,蔣 齊,王占軍,等. 溫度和光照對不同預處理野生甘草種子萌發和幼苗生長的影響[J]. 水土保持研究,2014,21(5):225-229,235.
[4]王秀英. 模擬酸雨對小麥種子萌發和幼苗生長的影響[J]. 水土保持研究,2014,21(6):265-270.
[5]李合生. 植物生理生化試驗原理和技術[M]. 北京:高等教育出版社,2000.
[6]張志良,瞿偉菁. 植物生理學實驗指導[M]. 3版.臺北:藝軒圖書出版社,2009.
[7]韓寶賀,朱 宏,張藝馨. 鎘對三種草坪草種子萌發與生長的影響[J]. 中國草地學報,2014,36(3):98-103.
[8]郭 媛,邱財生,龍松華,等. 鎘脅迫對不同地區亞麻主栽品種種子萌發的影響[J]. 作物雜志,2015(4):146-151.
[9]王新新,吳 亮,朱生鳳,等. 鎘脅迫對堿蓬種子萌發及幼苗生長的影響[J]. 農業環境科學學報,2013,32(2):238-243.
[10]孫園園,張光蘭,楊文杰,等. 鎘脅迫對1年生黑麥草種子萌發及幼苗初期生理生化特性的影響[J]. 種子,2014,33(8):25-29.
[11]楊居榮,黃 翌. 植物對重金屬的耐性機理[J]. 生態學雜志,1994,13(6):20-26.
[12]肖志華,張義賢,張喜文,等. 外源鉛、銅脅迫對不同基因型谷子幼苗生理生態特性的影響[J]. 生態學報,2012,32(3):889-897.
[13]宇克莉,孟慶敏,鄒金華. 鎘對玉米幼苗生長、葉綠素含量及細胞超微結構的影響[J]. 華北農學報,2010,25(3):118-123.
[14]賀 遠,王樹聲,劉海偉,等. 鎘濃度對烤煙幼苗鎘含量及生長和生理指標的影響[J]. 中國煙草科學,2014,35(2):37-42.
[15]朱 健,張志紅,范菲菲,等. 銅脅迫對海菜花幼苗生理特征的影響[J]. 江蘇農業學報,2015,31(4):883-886.
[16]陳國祥,施國新,何 兵,等. Hg、Cd對莼菜越冬芽光合膜光化學活性及多肽組分的影響[J]. 環境科學學報,1999,19(5):521-525.
