GRP78基因沉默對人卵巢癌SKOV3細胞增殖與凋亡的影響及機制
2017-03-14 02:36:14李平潘銀鳳王艷偉焦燕
山東醫藥 2017年5期
李平,潘銀鳳,王艷偉,焦燕
(1山東能源新汶礦業集團萊蕪中心醫院,山東萊蕪271103;2長春中醫藥大學)
GRP78基因沉默對人卵巢癌SKOV3細胞增殖與凋亡的影響及機制
李平1,潘銀鳳2,王艷偉1,焦燕1
(1山東能源新汶礦業集團萊蕪中心醫院,山東萊蕪271103;2長春中醫藥大學)
目的 探討葡萄糖調節蛋白78(GRP78)基因沉默對人卵巢癌SKOV3細胞增殖與凋亡的影響及機制。方法 設計、合成針對GRP78基因的短發夾RNA干擾序列,構建GRP78載體質粒,將其轉染入293T細胞,包裝產生慢病毒(GRP78 shRNA)。將培養好的SKOV3細胞隨機分為對照組、空白載體組、實驗組,將空白質粒、GRP78 shRNA分別轉染至空白載體組、實驗組。用MTT法檢測各組細胞生長抑制率,用流式細胞術檢測細胞凋亡率,用Western blotting法檢測細胞內GRP78、半胱氨酸天冬氨酸蛋白水解酶3(Caspase-3)及增強子結合蛋白同源蛋白(CHOP)蛋白表達。結果 轉染24、48、72 h,與對照組、空白載體組比較,實驗組細胞生長抑制率升高(P均<0.05);且實驗組各時間點細胞生長抑制率比較差異有統計學意義(P均<0.05)。轉染72 h,與對照組、空白載體組比較,實驗組細胞凋亡率升高(P均<0.05)。轉染72 h,與對照組、空白載體組比較,實驗組GRP78蛋白相對表達量降低,CHOP、Caspase-3蛋白相對表達量升高(P均<0.05)。結論 沉默SKOV3細胞內GRP78基因后,激活內質網應激凋亡信號通路,從而抑制細胞增殖、促進其凋亡。
卵巢腫瘤;SKOV3細胞;葡萄糖調節蛋白78;細胞增殖;細胞凋亡;內質網應激
卵巢癌是婦科常見的惡性疾病,其發病率占婦科癌癥的20%。中、晚期卵巢癌患者5年生存率不足20%[1]。卵巢癌以手術治療及化療為主,但效果并不理想。葡萄糖調節蛋白78(GRP78)是內質網上重要的分子伴侶蛋白,與腫瘤細胞生長、轉移及死亡抵抗有密切關系[2]。內質網應激(ERS)是近年來新發現的凋亡機制,現階段研究認為ERS發生的主要原因是內質網折疊蛋白功能異常,導致錯誤蛋白在內質網內堆積[3],短時間的ERS是細胞自我修復的一種方法,但長期的ERS即可引起細胞凋亡。已有研究表明,沉默GRP78可直接影響卵巢癌細胞的侵襲力和遷移力,但具體機制尚不完全清楚[4]。2016年1~6月,本研究采用慢病毒轉染技術沉默卵巢癌SKOV3細胞GRP78的表達,探討GRP78基因沉默對人卵巢癌SKOV3細胞增殖與凋亡的影響及機制,為卵巢癌的靶向治療提供理論依據。
1 材料與方法
1.1 材料 shRNA慢病毒(沈陽萬類生物科技有限公司);人卵巢癌SKOV3細胞(美國ATCC公司),293T細胞(上海紀寧生物公司);GRP78、增強子結合蛋白同源蛋白(CHOP)及半胱氨酸天冬氨酸蛋白水解酶3(Caspase-3)抗體(Boster公司),GAPDH內參抗體(中國Wanleibio公司);Annexin V細胞凋亡試劑盒(美國SIGMA公司),RPMI1640(美國GIBCO公司),光學顯微鏡(日本Olympus公司),WD-9405B型水平搖床(中國六一公司),LSR-Ⅱ流式細胞儀(美國BD公司)。
1.2 慢病毒載體構建 用Ambion公司軟件,針對人GRP78基因mRNA(NM 005347.3)設計shRNA序列,構建質粒,轉染293T細胞。根據其對GRP78的抑制效率,確定有效RNAi模版序列:正義3′-GGAGCGCAUUGAUACUAGATT-5′、反義3′-UCUAGUAUCAAUGCGCUCCTT-5′。慢病毒骨架質粒pLVTHM經BamHⅠ和EcoRⅠ雙酶切消化后與雙鏈DNA進行反應,對其克隆進行菌種培養擴增,抽提質粒DNA。將對數生長期的293T細胞接種于25 mL細胞培養皿,細胞生長至70%時,將重組的慢病毒骨架質粒與包裝輔助質粒用磷酸鈣共轉染至病毒包裝細胞293T,培養12 h后棄去含有轉染混和物的培養液,加入新鮮的細胞培養液繼續培養。收集轉染72 h的293T細胞上清液,于4 ℃、4 000 r/min離心10 min去除細胞碎片,經0.45 μm濾器過濾后置于25 000 r/min超速離心10 min, 而后以PBS液重懸病毒沉淀,制備完成慢病毒載體GRP78 shRNA,置于-80 ℃冰箱保存備用。
1.3 SKOV3細胞培養、分組及轉染 SKOV3細胞加入10%滅活胎牛血清的DMEM培養液中進行培養,條件:37 ℃、5% CO2,飽和濕度,2~3 d傳代1次。將細胞懸液以5×104/孔接種于24孔板,細胞融合80%~90%,隨機分為對照組、空白載體組及實驗組。將空白質粒、GRP78 shRNA分別轉染至空白載體組、實驗組。培養基中加入8 μg/mL ploybrene以提高感染效率,混勻后于37 ℃、5% CO2培養箱中孵育過夜。孵育24 h后將培養基更換為正常培養液,在原有條件下繼續培養48 h[4]。采用Western blotting法檢測各組細胞GRP78表達,以驗證GRP78是否轉染成功。
1.4 SKOV3細胞生長抑制率測算 采用MTT法。分別于轉染24、48、72 h取各組細胞,調整細胞為4×107/L,取200 μL接種于96 孔板中(每組細胞5個復孔),培養72 h,每孔加入5 g/L MTT 20 μL,繼續培養4 h后去上清液,逐一加入DMSO 100 μL振蕩混勻,待藍色晶體完全溶解反應后,應用酶標儀測定波長570 nm 處的吸光度值(A值),并計算細胞生長抑制率。細胞生長抑制率=(1-實驗組A值/對照組A值)×100%[5]。
1.5 SKOV3細胞凋亡率檢測 采用流式細胞術。轉染72 h取各組細胞,經消化、PBS沖洗、懸浮、300目尼龍網過濾后加入Binding Buffer 懸濁細胞300 μL,加入Annexin V-FITC 5 μL混合,再加入Propidium Iodide 10 μL充分混勻;常溫下避光反應20 min;采用流式細胞儀檢測細胞凋亡情況,計算細胞凋亡率。
1.6 SKOV3細胞內GRP78、Caspase-3及CHOP蛋白檢測 采用Western blotting法。轉染72 h取各組細胞,加入RIPA裂解液及蛋白酶抑制劑PMSF,提取蛋白質,測定濃度。取樣15 μL以10%的SDS-PAGE電泳,常規轉膜后脫脂奶粉封閉,加一抗anti-GRP78(1∶500)、anti-Caspase-3(1∶500)及anti-CHOP(1∶200)4 ℃孵育過夜,TBST洗脫5 min×3次,辣根過氧化酶標抗二抗室溫孵育2 h,TBST洗脫10 min×3次,ECL化學發光顯像[6]。目的蛋白的相對表達量=目的蛋白灰度值/GAPDH灰度值。

2 結果
2.1 沉默GRP78對SKOV3細胞增殖及凋亡的影響 轉染24、48、72 h,與對照組、空白載體組比較,實驗組生長抑制率升高(P均<0.05);且實驗組各時間點生長抑制率比較差異有統計學意義(P均<0.05)。轉染72 h,與對照組、空白載體組比較,實驗組凋亡率升高(P均<0.05)。見表1。
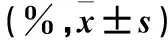
表1 各組細胞生長抑制率、凋亡率比較
注:與實驗組同時間點比較*P<0.05;與同組前一時間點比較,#P<0.05。
2.2 沉默GRP78對SKOV3細胞內GRP78、CHOP及Caspase-3蛋白表達的影響 轉染72 h,與對照組、空白載體組比較,實驗組GRP78蛋白相對表達量降低,CHOP、Caspase-3蛋白相對表達量升高(P均<0.05)。見表2。

表2 各組GRP78、CHOP和Caspase-3蛋白相對表達量比較
注:與實驗組比較,*P<0.05。
3 討論
癌細胞由于其自身瘋狂繁殖和易于轉移等特點,使傳統抗癌藥物難以將癌細胞殺死。現代科學技術對于細胞的靶向誘導逐漸成熟,誘導腫瘤細胞凋亡已成為治療腫瘤的希望和研究熱點[7,8]。大量研究證實,惡性腫瘤細胞瘋狂生長和繁殖,致使腫瘤細胞普遍缺氧并發生ERS,但發生ERS的腫瘤細胞并不走向凋亡,這可能與腫瘤細胞長期處于缺氧狀態已產生耐缺氧及ERS耐受有關[9~11]。
卵巢癌因發現時多為中晚期,預后極差,現階段手術治療效果欠佳,臨床治療多以化療為主[12]。但化療藥物具有不良反應大、靶向性不明確、易產生耐藥性等缺點,在臨床應用中受到了諸多限制。因此,尋找高效、靶向治療卵巢癌的方法是亟待解決的問題。GRP78又稱免疫球蛋白重鏈結合蛋白。Glembotski等[13]研究顯示,GRP78通過重新折疊內質網中的“錯誤折疊蛋白”,起到保護細胞不受損傷的作用。但近年來研究發現,腫瘤的微環境可誘導GRP78高表達,高表達的GRP78又通過內質網應激通路調節腫瘤細胞的生長及死亡抵抗[2],可見GRP78的高表達是腫瘤發病因素之一。Kaira等[9]研究發現,GRP78是預測腫瘤不良預后的獨立因素。通過抑制GRP78的表達可降低卵巢癌細胞的耐藥性,增強化療藥物對卵巢癌細胞的殺傷能力[14,15]。因此,降低腫瘤細胞中GRP78的表達,誘導卵巢癌細胞凋亡,可能成為靶向治療或輔助治療卵巢癌的新方向。
在眾多靶向沉默技術中慢病毒轉染沉默技術的穩定性和成功率最高[16]。本研究通過慢病毒轉染技術選擇性沉默GRP78基因表達。卵巢癌SKOV3細胞沉默GRP78基因后,細胞凋亡率升高,且細胞生長受到抑制,生長抑制率隨轉染時間的延長而增加,說明沉默GRP78可有效打破卵巢癌細胞ERS耐受,并減慢腫瘤細胞繁殖速度;且隨著時間延長,可加重未折疊蛋白在SKOV3細胞內質網中的堆積,激活ERS凋亡通路,促進腫胞凋亡。
CHOP又稱生長停滯及誘導DNA損傷的基因153,可直接參與調節下游凋亡基因的表達,通過直接調節核內的靶基因,增加細胞對ERS介導凋亡的敏感性[17]。Caspase家族至今已發現14位家庭成員,其中Caspase-12為ERS下游的凋亡因子,經Caspase-12→Caspase-8→Caspase-3“瀑布式”激活,凋亡執行核心蛋白Caspase-3可通過多種機制引發細胞凋亡[18]。本研究發現,當沉默GRP78基因表達后,SKOV3細胞中CHOP及Caspase-3表達升高,推測沉默GRP78后,SKOV3細胞內質網內堆積的錯誤折疊蛋白不能降解,可能激活ERS凋亡信號通路,通過PERK→eIF2a→ATF-4→CHOP、IRE1→CHOP、ATF-6→CHOP、Caspase-12→Caspase-8→Caspase-3等途徑促進細胞凋亡[19,20]。
總之,本研究結果表明,沉默GRP78基因可通過激活ERS凋亡信號通路,抑制卵巢癌SKOV3細胞增殖,促進細胞凋亡,此為靶向治療卵巢癌提供了理論依據。將沉默GRP78作為“生物導彈”,局部靶向給藥,可能成為臨床治療卵巢癌的新方法。本研究也有不足之處,ERS凋亡信號通路較多,本研究未逐一探明,下一步研究將證實SKOV3細胞沉默GRP78后,對ERS凋亡信號通路的影響。
[1] 張彤,沈丹華,陳云新,等. PTEN、PAX-2和β-catenin在Ⅰ型子宮內膜癌及其癌前病變中的表達及意義[J].現代婦產科進展,2014,23(2):85-88.
[2] 賈鵬宇,何苗,魏敏杰.GRP78在腫瘤發生發展及干性形成中的研究進展[J].生命的化學,2015,35(4):559-564.
[3] 王伊林,趙雅君,楊洋,等.磷酸肌酸鈉對慢病毒介導Calumenin蛋白沉默阿霉素損傷心肌細胞內質網應激信號通路的作用[J].臨床心血管病雜志,2015,31(10):1119-1122.
[4] 范麗梅,魏敏,楊淑麗,等.沉默GRP78基因對人卵巢癌SKOV3細胞侵襲力的抑制作用[J]. 中國老年學雜志,2010,30(23):3516-3519.
[5] Kim SH, Kim KY, Yu SN, et al. Silibinin induces mitochondrial NOX4-mediated endoplasmic reticulum stress response and its subsequent apoptosis[J]. BMC Cancer, 2016,16(1):452.
[6] 王伊林,單曉彤,柴花,等.丹參酮ⅡA磺酸鈉腹腔注射對阿霉素心肌病大鼠心功能及左室纖維化的影響[J].山東醫藥,2016,56(7):34-36.
[7] 胡明英,王曉莉,畢學輝,等.RNA干擾沉默AKT基因對卵巢癌SKOV3細胞增殖與凋亡影響研究[J]. 中華腫瘤防治雜志,2013,20(24):1869-1873.
[8] 劉曼,胡明英,王曉莉,等. AKT基因沉默對人卵巢癌細胞增殖及Fas、Fasl、Caspase-3蛋白表達的影響[J].山東醫藥,2016,56(7):29-30.
[9] Kaira K, Toyoda M, Shimizu A, et al. Expression of ER stress markers(GRP78/BiP and PERK) in patients with tongue cancer[J]. Neoplasma, 2016,63(4):588-594.
[10] 吳芳,安永康,朱艷琴.內質網應激與腫瘤細胞凋亡的研究進展[J].現代腫瘤醫學,2014,22(9): 2228-2231.
[11] Zhang B, Han H, Fu S, et al. Dehydroeffusol inhibits gastric cancer cell growth and tumorigenicity by selectively inducing tumor-suppressive endoplasmic reticulum stress and a moderate apoptosis[J]. Biochem Pharmacol, 2016,15(104):8-18.
[12] 高翔,劉永軍,李曉丹,等.細胞因子誘導殺傷細胞聯合多西他賽、卡鉑治療晚期卵巢癌的療效及安全性[J].山東醫藥,2015,55(5):82-84.
[13] Glembotski CC. The role of the unfolded protein response in the heart[J]. J Mol Cell Cardiol, 2008,44(3):453-459.
[14] Stope MB, Hettenbach D, Kaul A, et al. The Tumor Suppressor MicroRNA-1 exhibits restricted inhibition of proliferation of ovarian cancer cells[J]. Anticancer Res, 2016,36(7):3329-3334.
[15] 范麗梅,張立會,崔松花,等.特異性下調GRP78基因表達對卵巢癌細胞耐藥性的影響[J].吉林大學學報(醫學版),2011,37(2):255-259.
[16] Lei CJ, Yao C, Pan QY, et al. Lentivirus vectors construction of SiRNA targeting interference GPC3 gene and its biological effects on liver cancer cell lines Huh-7[J]. Asian Pac J Trop Med, 2014,7(10):780-786.
[17] 黃金燕,趙連友,林可,等.高血壓大鼠動脈血管平滑肌細胞中GRP78和CHOP表達的變化及對血管重構的影響[J].心臟雜志,2011,23(1):20-26.
[18] Zhang Q, Liu J, Chen S, et al. Caspase-12 is involved in stretch-induced apoptosis mediated endoplasmic reticulum stress[J]. Apoptosis, 2016,21(4):432-442.
[19] Ma J, Yang YR, Chen W, et al. Fluoxetine synergizes with temozolomide to induce the CHOP-dependent endoplasmic reticulum stress-related apoptosis pathway in glioma cells[J]. Oncol Rep, 2016,36(2):676-684.
[20] 趙彥超,顧耘.細胞凋亡通路研究進展[J].現代醫學,2013,41(4):285-288.
李平(E-mail:wylteam@163.com)
10.3969/j.issn.1002-266X.2017.05.012
R737.31
A
1002-266X(2017)05-0040-03
2016-07-15)
