氮磷添加對亞熱帶常綠闊葉林土壤微生物群落特征的影響
2017-03-09 08:28:14王晶晶許崇華王澤夫徐小牛
生態學報 2017年24期
關鍵詞:利用
王晶晶, 樊 偉, 崔 珺, 許崇華, 王澤夫, 徐小牛
安徽農業大學林學與園林學院,合肥 230036
土壤微生物是土壤中的活力成分,是土壤養分循環的推動力,土壤中一系列的過程都是以碳、氮循環為中心,其物質的流動和能量的循環都是在不同微生物的協同下完成,土壤微生物一直在挖掘有機物中C-N鍵的能量,調控土壤中有機化合物的轉化和養分的釋放[1]。微生物量是土壤養分的源和庫,也是指示土壤肥力的敏感指標[2]。長期不同的施肥措施不僅影響著土壤的理化性質,也影響著適宜各種微生物生長發育所需要的養分、水分、空氣、酸堿度、滲透壓和溫度等條件,因此土壤微生物的特性也隨之發生改變[3]。土壤微生物群落功能多樣性反映了土壤微生物群落的生態特征,近幾年來,土壤微生物生態的研究已經成為生態學研究的熱點之一[4]。
氮、磷均是植物生長所必需的大量營養元素,在植物生長、發育和繁殖等過程中有著重要的作用,自然狀態下,很多生態系統生產力都表現為氮限制、磷限制或者氮磷共限制[5]。自然條件下,氮進入陸地生態系統的主要途徑是固氮生物對大氣中氮氣的固定,而磷則主要通過原生礦物的風化作用進入生態系統,因此一般認為生態系統在演替早期主要表現為氮限制,在演替后期則主要為磷限制;控制氮可利用性的反應主要為生物反應,而控制磷可利用性的反應則主要是地球化學反應[6]。氮素也是土壤中較為活躍的營養元素,以有機氮和無機氮兩種形態存在于土壤中,氮素的轉化主要有氨化作用、硝化作用、反硝化作用和生物固定,其中硝化和氨化作用是氮素有效化的過程,有利于植物的吸收。近些年來,人類大量施用無機肥料來增加土壤中的有效養分,促進植物的生長,氮、磷、鉀作為“肥料三要素”,被植物真正吸收利用的僅僅是其中的小部分,大部分不能被植物所吸收利用的肥料,在土壤中會發生揮發、淋溶和固定等[7],人類大量使用氮肥來增加土壤可利用性氮素的水平,促進植物的生長,但是氮素的過量使用會對森林生態系統的結構和功能產生重要影響,土壤微生物對氮增加的響應有多樣性的特點[8],因此合理的添加氮磷對森林生態系統可持續發展具有重要實踐和理論意義。
亞熱帶常綠闊葉林生物多樣性豐富、群落結構復雜、生物量及生產力高、植物群落和生態系統的穩定性強,生態效益良好,在我國分布廣泛,具有涵養水源、保持水土和維持生物多樣性等重要作用[9]。氮素添加對常綠闊葉林土壤生態系統氮循環的影響有一些相關報道,而氮、磷添加對不同土層土壤微生物的影響還鮮為報道。為此,本研究在黃山西脈仙寓山風景區境內設置樣地,連續五年開展了氮、磷添加試驗,探究不同施肥梯度下土壤微生物量碳氮的變化,采用Biolog-ECO微平板法分析不同施肥措施對土壤微生物群落功能多樣性的影響,可為森林土壤中碳氮轉化、碳庫存儲、土壤肥力監測以及建立提高森林生產力的施肥模式提供有力支撐。
1 研究地區與研究方法
1.1 研究地區概況
試驗樣地設在安徽石臺縣仙寓山景區,地理位置30°1′47.44″ N、117°21′23.33″ E,屬于亞熱帶濕潤氣候,春季溫涼多雨,年平均氣溫為16℃,海拔為375 m,年降水量為1620 mm以上。土壤類型為黃棕壤,質地輕粘,呈酸性反應。調查區植被豐富,森林覆蓋率達85%,地帶性森林植被類型在海拔600 m以下多為亞熱帶常綠闊葉林,也分布著大面積的杉木(Cunninghamialanceolata)人工林,在海拔600 m以上分布有常綠落葉闊葉混交林、落葉闊葉林及馬尾松(Pinusmassoniana)林等。根據王氏家譜記載試驗區森林為風水林,面積近20 hm2,近100多年來沒有人為經營活動,如間伐、采伐等。試驗林為甜櫧(Castanopsiseyrei)老齡林分,其林齡在200年生左右,該林分以甜櫧為優勢種群,樹種豐富度較高,基于本底調查結果,試驗樣地林分組成結構相似,立木密度590—680株/hm2(DBH ≥ 5.0 cm),共出現喬灌木樹種52種,主要伴生喬木有青岡(Cyclobalanopsisglauca)、香桂(Cinnamomumsubavenium)、山杜英(Elaeocarpussylvestris)、楓香(Liquidambarformosana)、藍果樹(Nyssasinensis)、樹參(Dendropanaxdentiger)等;主要伴生灌木有山礬(Symplocossumuntia)、冬青(Ilexspp.)、老鼠矢(Symplocosstellaris)、豹皮樟(Litseacoreanavar.sinensis)、尖連蕊茶(Camelliacuspidata)等;主要伴生草本有油點草(Tricyrtismacropoda)、蛇根草(Ophiorrhizajaponica)、蕨類等,豐富度較高。
1.2 試驗設計
本試驗設計4個處理:不施肥(CK,0 kg N/hm2)、低氮(LN,50 kg N/hm2)、高氮(HN,100 kg N/hm2)、高氮+磷(HN+P,100 kg N/hm2+50 kg P/hm2)。氮肥為硝酸銨(NH4NO3)、磷肥為過磷酸鈣(Ca(H2PO4)2),其中每種施肥處理各設有個重復。施肥方式為噴霧器濕灑法,將肥料溶于20 L水中充分溶解,每月噴施一次,自2011年已連續施肥5年。
1.3 樣品采集與分析
1.3.1 土樣采集
試驗區設置12個面積均為20 m×20 m的固定樣地(其中3、7、8號為CK,1、6、10號為LN,0、5、11號為HN,2、4、9號為HN+P樣地),每個樣地之間設有8—10 m的隔離帶,以防不同處理間相互干擾影響。2015年12月在各樣地分別取0—10、10—20、20—30 cm 3個土層,X型布點各采集5鉆,過篩去除凋落物、細根及石礫等,混合均勻后放入4℃下冷藏保存。其中一部分用于測定土壤理化性質,另一部分過2 mm篩后裝入無菌袋,用于土壤微生物群落功能多樣性的測定。
1.3.2 土壤理化性質及土壤微生物量的測定
土壤含水率采用烘干法測定;土壤溶液pH按水土比2.5∶1的充分混合,稍加靜置后使用pH計測定;全磷和速效磷使用流動注射分析儀(FIAstar 5000, FOSS, Denmark)測定,可溶性有機碳使用TOC分析儀(Multi C/N 3100, Jena Analytik, Germany)測定,全碳和全氮使用元素分析儀(EA 3000, Vector, Italy)測定。微生物生物量碳(MBC)、氮(MBN)則使用氯仿熏蒸法,TOC分析儀測定。林地土壤本底情況如(表1)所示。
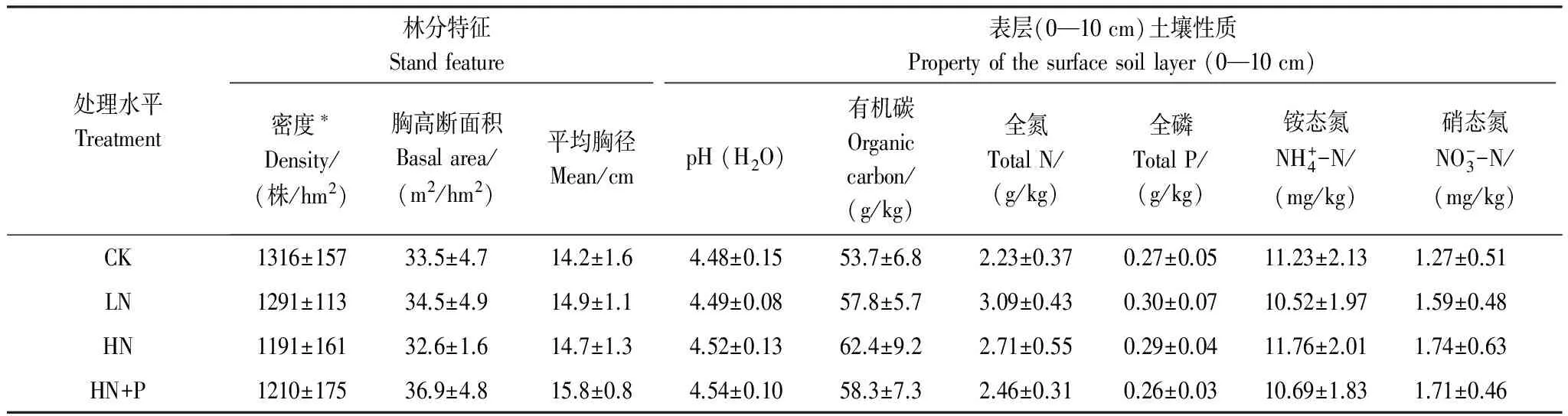
表1 試驗林分結構及表層土壤特性
*DBH≥5 cm; CK:對照Control; LN:低氮處理Low N treatment; HN:高氮處理High N treatment; P:磷Phosphorus
1.3.3 土壤微生物群落功能多樣性的測定
采用Biolog-ECO微平板法: 稱約10 g新鮮土樣放入250 mL的三角瓶中,加入100 mL已滅菌0.85% NaCl溶液,搖床震蕩30 min (轉速170 r/min), 將溶液稀釋103倍,稀釋后的溶液放置4℃冰箱內靜置2—3 min, 取上清液倒入無菌培養皿,使用八通道移液槍(每孔150 μL)移入ECO微平板內,最后將已加溶液的ECO板放入28℃生化培養箱中培養7 d,每隔24 h用Biolog自動讀取儀讀取一次數據。
1.4 數據處理
Biolog-ECO平板測定的每孔顏色平均變化率(average well color development,AWCD)值,用來表示微生物群落利用單一碳源的能力即微生物的整體代謝活性[10]:
AWCD=∑(Ci-R)/31
式中:Ci為第i個孔590—750 nm的吸光度值,R為對照孔吸光值,若Ci-R小于0,其吸光度值均計為0。
土壤微生物功能多樣性指數[11]:
Shannon指數:
H=-∑PilnPi
Simpson指數:
D=1-∑Pi2
式中:Pi=(Ci-R)/∑(Ci-R)表示第i孔的相對光密度值與整個微板相對光密度值總和的比值,ni表示第i孔的相對吸光度(Ci-R)。
數據采用SPSS 17.0軟件進行單因素方差分析,應用R 3.3.1進行雙因素、聚類分析和主成分分析。
2 結果與分析
2.1 不同施肥處理對不同土層土壤理化性質的影響
長期施加氮磷對土壤性質,如pH、全氮、全磷、速效磷、銨態氮和硝態氮都有著顯著的影響。在不同土層間pH、全氮、全磷、銨態氮均表現出了極顯著的差異(表2)。
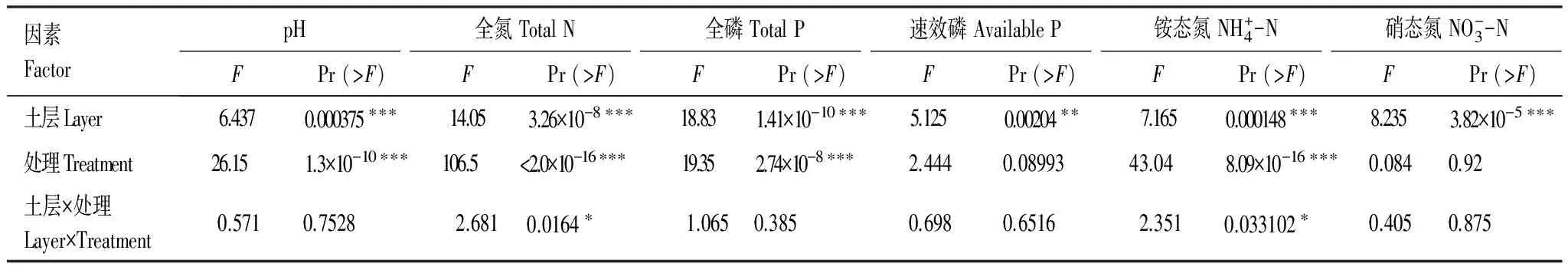
表2 不同土層和不同施肥處理土壤理化性質的雙因素方差分析結果
顯著水平/Significant levels: ***,P< 0.001; **,P< 0.01; *,P< 0.05
HN+P處理的土壤pH值在0—20 cm顯著低于CK與HN,20—30 cm土層,LN和HN+P處理的土壤pH值也表現出低于CK與HN(表3)。全氮、全磷的含量隨施肥處理方式的不同存在著顯著的差異,0—10 cm土層,全氮含量表現為,CK明顯低于各種處理,全磷含量表現為CK與HN+P處理高于其他處理。10—30 cm土壤中,CK的全磷含量顯著高于氮磷添加處理。0—20 cm土層中HN+P處理的速效磷明顯高于HN。不同土層中銨態氮與硝態氮的含量,CK都顯著低于各種處理,而LN處理與HN處理之間并沒有明顯的差異,HN+P處理的銨態氮、硝態氮含量在各土層中低于LN、HN處理。
2.2 不同處理對土壤微生物量碳、微生物量氮的影響
土壤微生物量碳、氮與微生物量碳氮比,在不同土層之間均表現為極顯著性差異,土壤微生物量碳在不同施肥處理間的差異性也極其顯著,而土壤微生物量氮差異性卻不明顯,土層和處理間交互作用并沒有差異(表4)。
MBC、MBN含量均隨土層的加深而降低,不同施肥處理下MBC的變化(圖1),在表層0—10 cm為HN>CK>HN+P>LN, 10—20 cm為HN>CK>LN>HN+P,20—30 cm為HN>CK>HN+P>LN;而MBN的變化則表現為:表層0—10 cm為HN>LN>CK>HN+P,10—20 cm為HN>LN>HN+P>CK,20—30 cm為HN>LN>HN+P>CK。
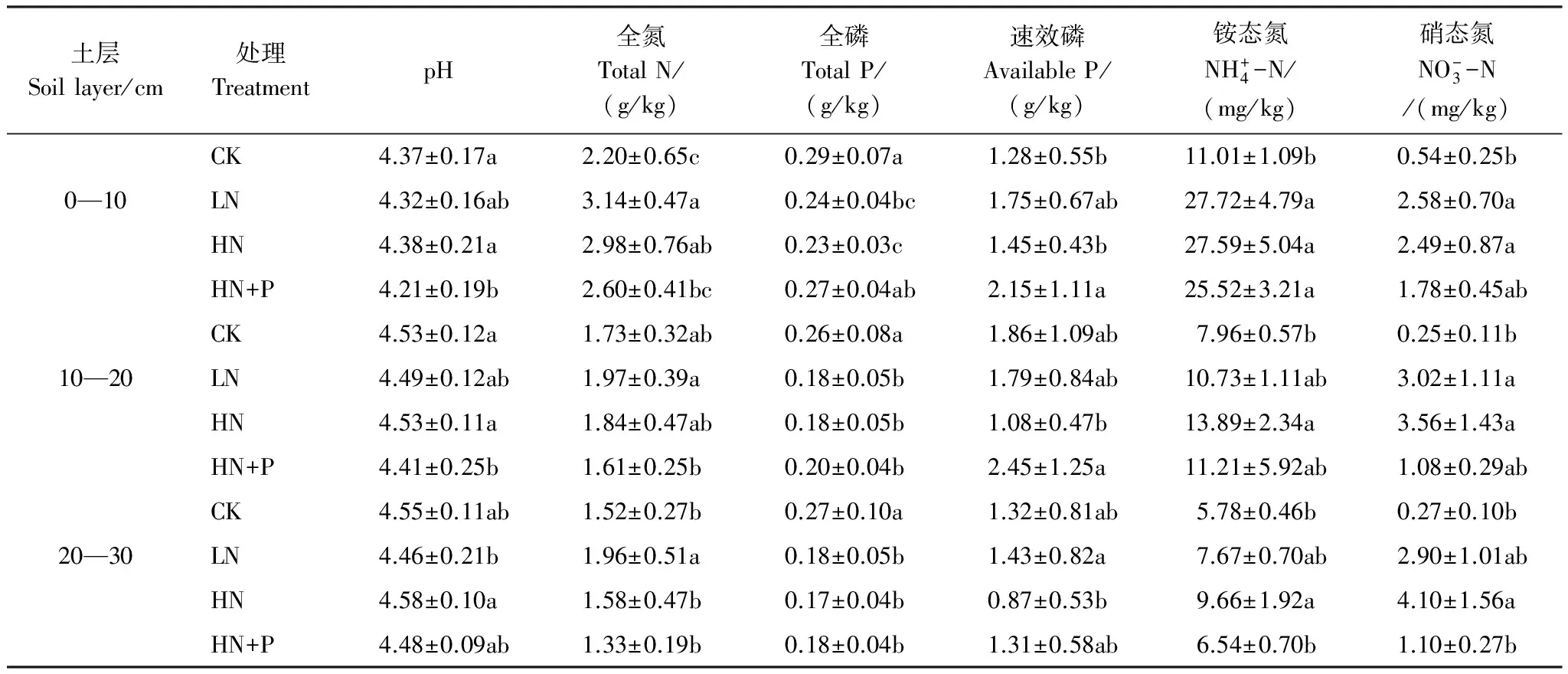
表3 不同的氮磷添加條件下不同土層的土壤理化性質
同列不同字母表示差異顯著(P<0.05)
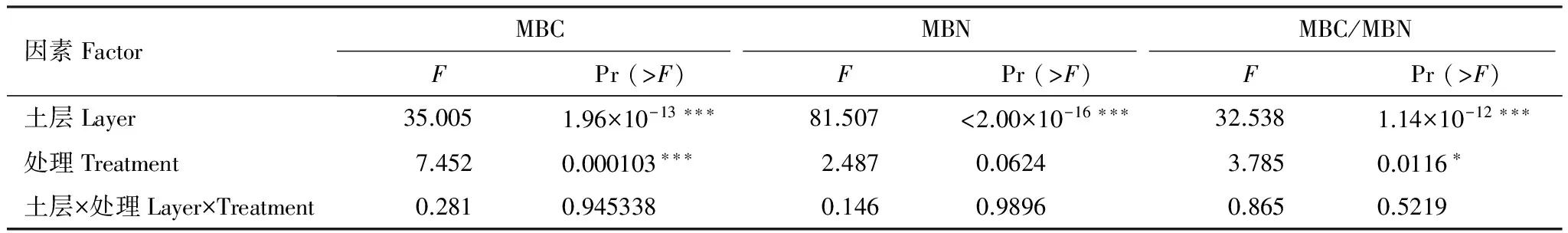
表4 不同土層和不同施肥處理土壤微生物量碳氮的雙因素方差分析結果
顯著水平/Significant levels: ***,P< 0.001; **,P< 0.01; *,P< 0.05;MBC:微生物量碳Microbial biomass carbon; MBN:微生物量氮Microbial biomass nitrogen
不同土層間的MBC,HN和LN處理均呈現了顯著性差異,在10—30 cm土層,HN與HN+P的差異也比較明顯;MBN在不同施肥處理下均沒有表現出差異性;0—10 cm和20—30 cm土層土壤微生物量碳氮比(MBC/MBN)在HN和LN處理上存在著差異。
CK處理的MBC、MBN和MBC/MBN在不同土層間均表現出顯著性差異,LN與HN處理的MBC含量0—10 cm明顯高于20—30 cm土層,HN與HN+P施肥水平下,表層土的MBC含量顯著高于底層土。MBN含量在不同施肥條件下,均表現出表層土0—10 cm顯著高于10—30 cm。
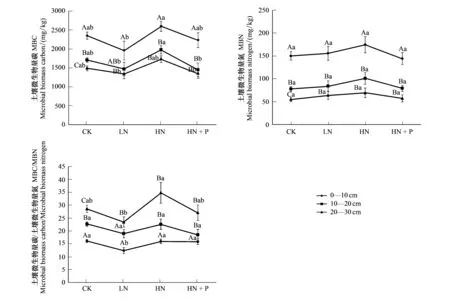
圖1 不同水平氮磷添加條件下不同土層土壤微生物量C、N的變化Fig.1 Changes of soil microbial biomass carbon and nitrogen in different soil layers under different nitrogen and phosphorus additions
2.3 不同施肥處理對土壤微生物群落碳源利用的動力學特征的影響
隨著培養時間的延長,土壤微生物群落AWCD值逐漸增加,從培養24 h開始進入對數增長期,AWCD值快速增長到72 h時變化速率最快,此時微生物代謝活性最為旺盛,大量碳源被利用,隨后緩慢增長直至趨向平穩(圖2)。
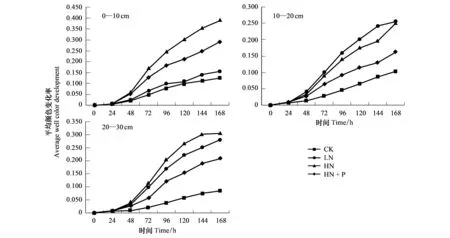
圖2 不同水平氮磷添加條件下不同土層間AWCD值的變化Fig.2 Changes of AWCD of soil microorganism in different soil layers under different nitrogen and phosphorus additionsAWCD:平均顏色變化率Average well color development
表層土的整體吸光度值都大于其他土層,不同施肥處理土壤中微生物對單一碳源利用程度存在著差異,差異性并不是很顯著(表5)。表層0—10 cm的AWCD值大小為HN>HN+P>LN>CK, 10—20 cm的AWCD值為LN>HN>HN+P>CK, 20—30 cm的AWCD值為HN>LN>HN+P>CK。
由(圖2)所示的AWCD值可知,不同土層的土壤微生物代謝活性,CK表現為:0—10 cm>10—20 cm>20—30 cm;LN處理表現為:10—30 cm>0—10 cm;HN與HN+P處理表現為:0—10 cm>20—30 cm>10—20 cm。

表5 不同土層和不同施肥處理AWCD值的雙因素方差分析結果
顯著水平/Significant levels: ***,P< 0.001; **,P< 0.01; *,P< 0.05
2.4 不同施肥處理對土壤微生物群落多樣性指數的影響
對土壤微生物培養72 h的AWCD值進行McIntosh、Shannon和Simpson多樣性指數分析,可在土壤微生物對不同碳源利用的數量特征上,反應土壤微生物群落物種的組成和分布情況。由表6可知,0—10 cm土層中,不同施肥處理的土壤微生物3個多樣性指數的差異性均達到了顯著水平,其中McIntosh指數,HN、HN+P與LN、CK呈現出顯著性差異,HN處理的土壤微生物Shannon指數與其他處理呈現出顯著差異,CK的土壤微生物Simpson指數與HN、HN+P也表現出顯著差異性;10—30 cm土層中,CK的土壤微生物3個多樣性指數與其他處理的均呈現顯著差異性。

表6 土壤微生物群落多樣性指數
同列不同字母表示差異顯著(P<0.05)
由三個土層不同施肥處理的土壤微生物功能多樣性指數聚類分析(圖3)可知,0—10 cm土層HN與HN+P聚為一類,CK與LN聚為一類;10—20 cm土層CK為一類,HN、LN和HN+P聚為一類,20—30 cm土層則HN與LN聚為一類,CK與HN+P聚為一類。
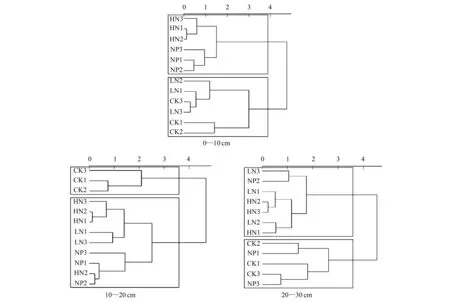
圖3 不同氮磷添加條件下不同土層微生物群落多樣性指數的聚類分析Fig.3 Cluster analysis of diversity index of microbial communities in different soil layers under different nitrogen and phosphorus additions
2.5 不同施肥處理對土壤微生物群落碳源利用率的影響
Biolog-ECO板31種碳源可以分為六大類,其中糖類/碳水類(10種)、氨基酸類(6種)、羧酸類(7種)、多聚物(4種)、多酚類(2種)、多胺類(2種)[12]。不同土層土壤微生物的碳源利用模式基本相同(圖4),0—10 cm土層中,CK、LN、HN處理的土壤微生物對碳源利用率大小順序均為:羧酸類>碳水類>氨基酸類>多聚物類>多酚類>多胺類,而HN+P處理,碳水類利用率高于羧酸類;10—20 cm土層中,CK的土壤微生物碳源利用率大小順序為:羧酸類>碳水類>氨基酸類>多酚類>多聚物類>多胺類,而LN、HN與HN+P處理表現為多聚物類>多酚類, HN+P處理的碳水類>羧酸類;20—30 cm土層中,CK的土壤微生物碳源利用率大小順序為羧酸類>碳水類>氨基酸類>多聚物類>多酚類>多胺類,而LN與HN為碳水類>羧酸類,HN+P為胺類>多酚類。
CK的土壤微生物在各土層上都表現出對羧酸類碳源的偏好,且與其他處理相比,羧酸類碳源利用率最高。LN處理的土壤微生物對多聚物類和羧酸類的利用率在土層上的變化比較明顯,HN與HN+P表層土壤微生物對碳源利用率并沒有明顯區別,而對多酚類碳源的利用率隨土層加深而降低。土壤微生物整體表現出對羧酸類、碳水類和氨基酸類的利用率較高,其次是多聚物類,對多胺類和多酚類碳源利用率最低。
2.6 土壤微生物群落對碳源利用的主成分分析
對培養72 h的各土層不同處理的AWCD值,進行群落功能多樣性主成分分析(PCA),0—10 cm的第一主成分(PC1)和第二主成分(PC2)的方差貢獻率分別是76.47%和6.65%,累計方差貢獻率83.12%;10—20 cm的第一主成分(PC1)和第二主成分(PC2)的方差貢獻率分別是44.54%和7.71%,累計方差貢獻率52.25%;20—30 cm的第一主成分(PC1)和第二主成分(PC2)的方差貢獻率分別是54.92%和10.86%,累計方差貢獻率65.87%。PC1與PC2是微生物群落碳源利用變異的主要成分,可以解釋變異的大部分信息。不同土層間施肥處理的土壤微生物群落在碳源利用上有明顯的空間變異性(圖5),0—10 cm土層,CK與LN處理均分布在PC1軸的負方向上,HN處理分布在PC1軸的正方向,HN+P處理分布在PC2軸的正方向。10—20 cm土層,CK分布在PC1軸的負方向,LN分布在PC2軸的負方向,HN分布在PC1軸的正方向,HN+P分布在PC2軸的正方向。20—30 cm土層,CK處理分布在PC1軸的負方向,LN與HN+P分布在PC2軸的正方向,HN分布在PC2軸的負方向。整體來看,不同N、P添加對0—10 cm土層的土壤微生物群落穩定性影響較大,空間分異性明顯,而10—30 cm土層的土壤微生物群落在碳源利用上的變異性并不是很明顯。
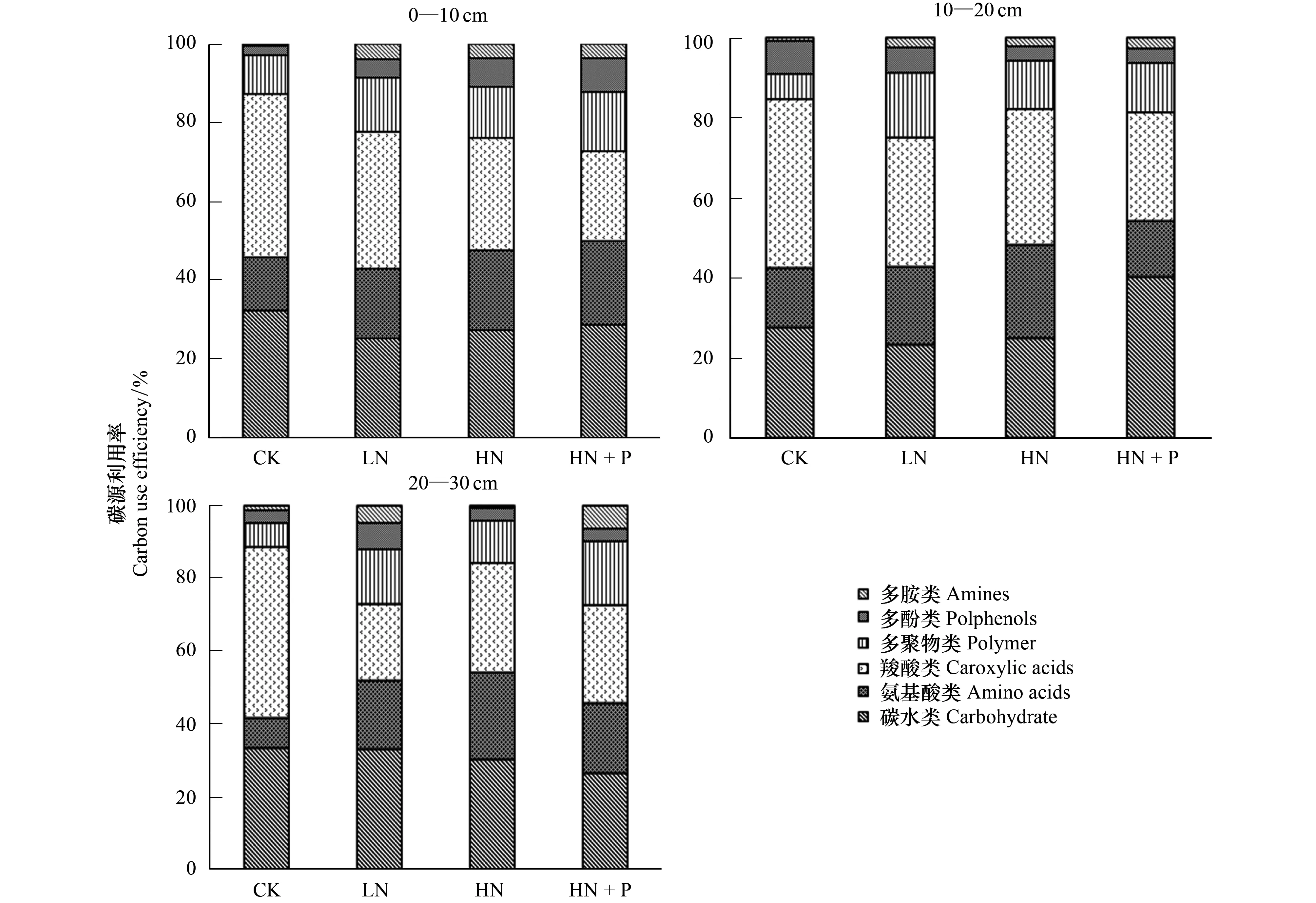
圖4 不同土層土壤微生物群落對不同碳源的利用率Fig.4 Use efficiency of carbon sources by soil microbial communities in different soil layers
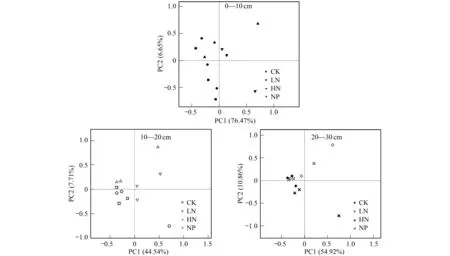
圖5 不同土層土壤微生物群落功能多樣性的主成分分析Fig.5 Principal component analysis of soil microbial community functional diversity in different soil layers
不同土層的31種碳源在2個主成分上的載荷值(表7),反映主成分與碳源利用的相關系數,其值越高表示該碳源對主成分的影響越大。0—10 cm土層中,氨基酸類、碳水類、羧酸類與PC1的相關性較高,羧酸類、碳水類、多聚物類與PC2的相關性較高,因此,0—10 cm土層的土壤微生物群落代謝特征,引起分異作用的主要碳源是碳水類與羧酸類,尤其碳水類碳源影響最大。10—20 cm土層中,羧酸類、碳水類、氨基酸類與PC1相關性較高,羧酸類、碳水類、氨基酸類與PC2相關性較高;20—30 cm土層中,碳水類、羧酸類、氨基酸類與PC1相關性較高,羧酸類、碳水類、多胺類與PC2相關性較高。綜上所述,不同土層土壤微生物功能多樣性在不同氮磷添加處理上的差異,主要體現在碳水類和羧酸類碳源的利用上。
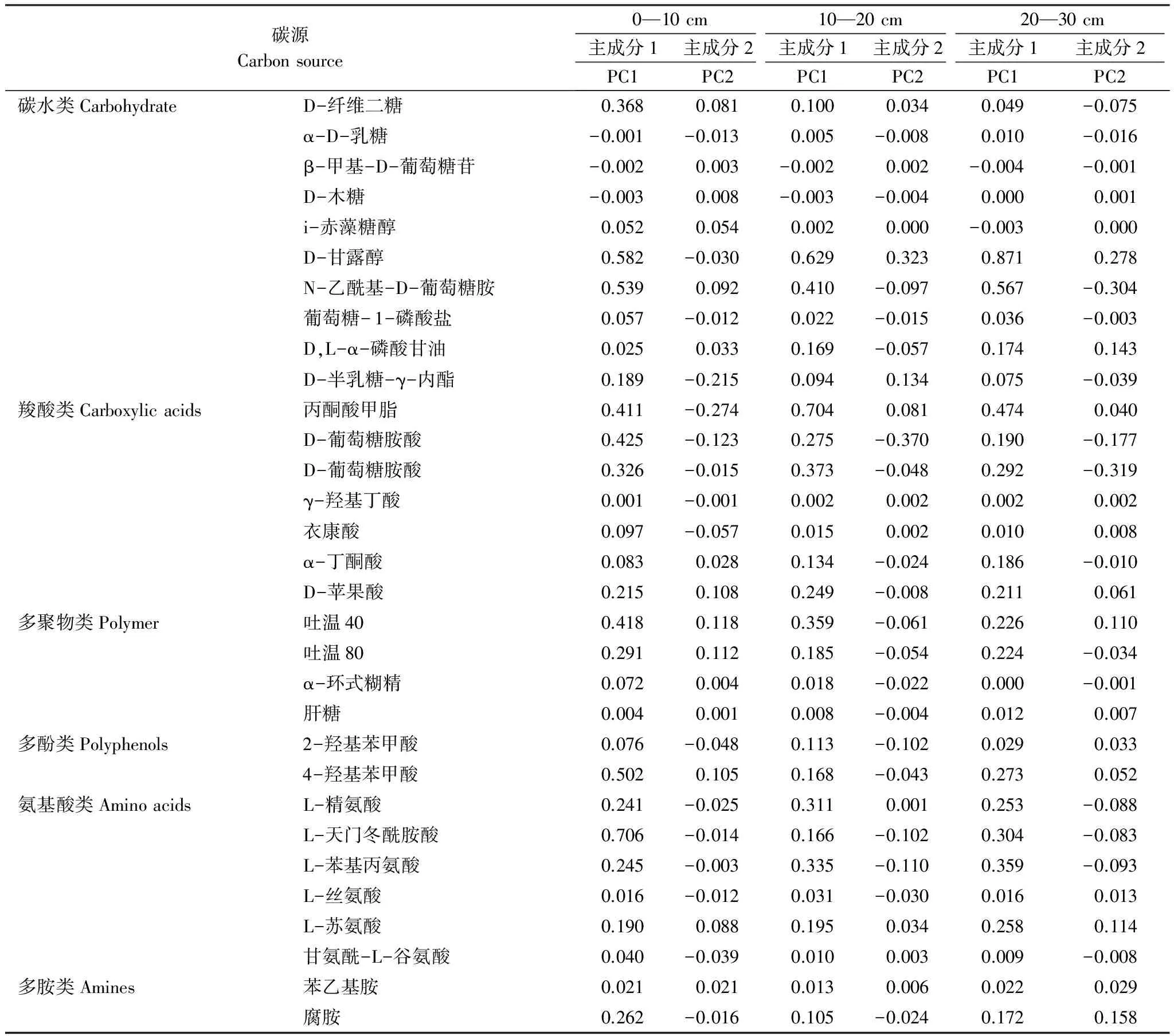
表7 31種碳源與主成分1、2的相關系數
2.7 土壤微生物多樣性指數與土壤理化性質的相關性
從(表8)可以看出,全氮、銨態氮與土壤微生物群落多樣性指數呈正相關,土壤微生物群落多樣性指數之間均呈顯著正相關,速效磷與土壤微生物群落多樣性指數呈顯著負相關。土壤pH與土壤理化性質的其他指標及土壤微生物群落多樣性均呈現負相關關系。
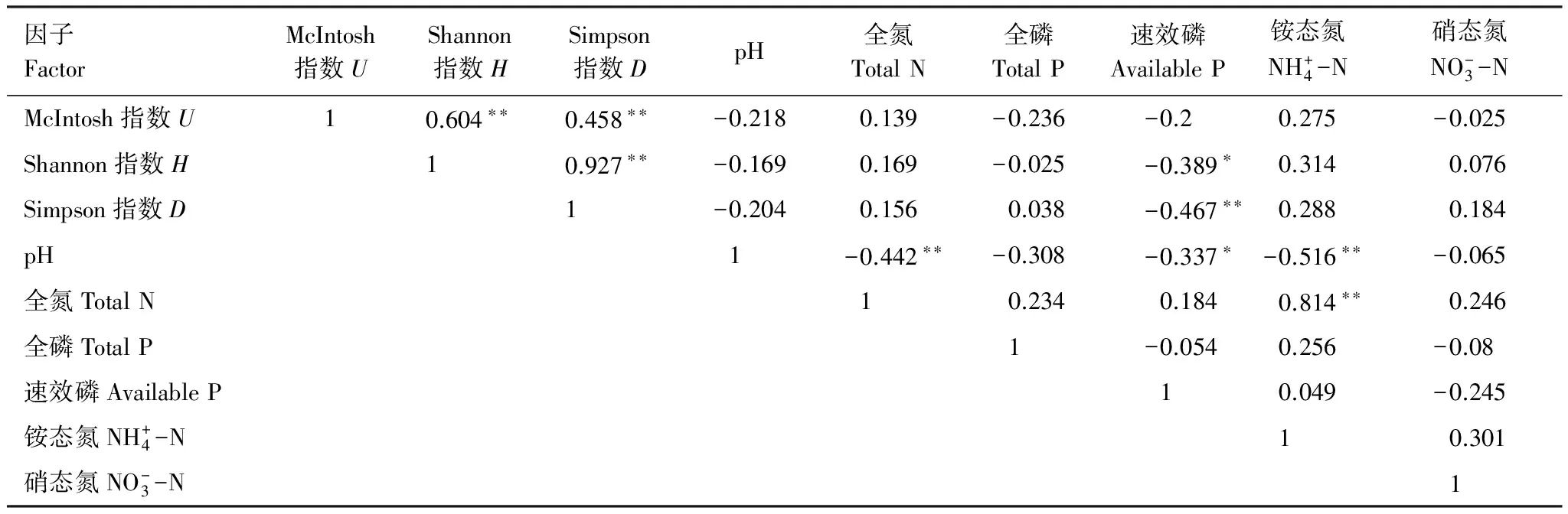
表8 土壤微生物群落多樣性指數與土壤化學性質的相關系數
*P<0.05; **P<0.01
3 討論
土壤微生物生物量碳、氮雖然在土壤中的含量較低,由于土壤微生物的繁殖速度快、周轉時間短且速率快,因此MBC、MBN在森林生態系統中很大程度上控制著養分的循環和能量的轉化[13]。本研究表明,MBC、MBN隨土層的加深而降低,而且表土層的AWCD值明顯高于其他土層,表明表土層的微生物群落的代謝活性最為活躍。有研究表明MBC含量與土壤有機質含量、土壤有機質活性、易礦化程度有關[14]。與對照相比,HN處理的土壤MBC、MBN明顯高于其他處理水平,可能是這一水平的氮添加,并沒有破壞土壤團聚體減弱微生物的活性,反而為微生物提供足夠的氮源,利于微生物的生長繁殖,從而促進土壤微生物量N的合成,同時高氮添加也增加了植物生物量和凋落物產量,增加了微生物可利用碳源的量。張彥東等[15]研究施氮肥對金沙江干熱河谷退化草地土壤微生物的影響,發現高氮施肥使微生物的數量和生物量有明顯的增加,這與我們的研究結果一致。土壤MBC/MBN比可反映微生物群落結構信息[16],HN處理MBC/MBN變化顯著,預示著高氮處理的土壤微生物群落結構變化是其微生物生物量較高的主要原因。
本研究表明,隨著土壤深度的增加土壤微生物對碳源底物的利用不斷降低。微生物對不同碳源的利用能力很大程度上取決于微生物的種類和固有性質,利用微平板法測定微生物對不同單一碳源的利用能力,可以比較分析不同微生物群落,得出其群落水平多樣性[17]。McIntosh多樣性指數反映土壤微生物種類數的不同,并能區分不同利用程度;Shannon多樣性指數反映土壤微生物群落利用碳源程度的變化度和差異度,Simpson多樣性指數可反映土壤微生物群落對常見碳源的利用程度[18]。不同氮、磷添加的表層土壤微生物群落多樣性指數均表現出顯著性差異,說明表層土壤微生物群落在利用常見碳源的程度、碳源種類的均勻度、豐富度及不同種碳源的利用程度上,氮、磷添加對其影響明顯。CK與LN處理的表層0—10 cm土壤多樣性指數聚為一類,CK在10—20 cm的土層多樣性指數單為一類,CK與HN+P處理的20—30 cm聚為一類,表明不同氮、磷添加,不同土層多樣性指數差異性較為一致。
Bach等[19]認為,凋落物和根系分泌物是土壤微生物的主要碳源,天然闊葉林群落物種多樣,結構更為復雜,具有較高的根系生物量和根系分泌物,為土壤微生物提供了豐富的碳源,其土壤微生物的結構和功能也相應增加[20]。劉秉儒等指出地上植物多樣性會通過凋落物的差異對土壤微生物功能代謝產生重要影響[21]。不同氮、磷添加,影響著表土層微生物對碳底物的利用能力,隨著氮添加劑量的增加,土壤微生物對酚類碳源的利用能力明顯提高(表7和圖4)。酚類物質影響土壤微生物的自然平衡,降低土壤中酚類物質的積累,可在一定程度上抑制有害微生物的繁殖。因此,高氮處理有助于土壤中酚類物質的消耗。氨基酸是土壤有機氮的重要組成部分,微生物在生長代謝過程中可利用氨基酸作為其氮源合成植物生長調節劑,促進其生長[22]。氨基酸類物質對土壤中有機質有正激發效應[23],HN與HN+P處理土壤微生物對氨基酸類碳源利用率提高,說明高劑量的氮添加可促進土壤有機質的分解。
主成分空間分布的不同反映了土壤微生物利用碳源的種類和程度的差異[24],不同處理對表土層碳源利用的離散程度較大,而10—30 cm土層的分布相對集中,因此表土層土壤微生物群落功能穩定性較差,即不同水平的氮、磷添加對表層土壤微生物碳源利用的空間分布有著顯著影響。土壤微生物功能多樣性在主成分分離中起主要貢獻作用的是碳水類和羧酸類碳源。
總之,天然闊葉林的表層土富集著大量的凋落物,養分充足,且具有較高的根系活動和細根周轉,利于提高土壤微生物量與微生物活性,下層土壤生境條件逐漸變差,土壤微生物代謝活性變弱[25]。土壤微生物群落多樣性也隨土層深度的增加而遞減[26]。不同氮磷添加,微生物生物量碳、氮和功能多樣的變化取決于土壤深度和施肥量[27]。較高劑量的氮、磷添加可改善土壤的理化性質,增加植物生物量和凋落物的產量,并且促進土壤微生物的代謝活性,提高土壤微生物生物量碳、氮含量,對土壤微生物功能多樣性及土壤微生物的物種多樣性都有顯著影響。
[1] 孫鳳霞, 張偉華, 徐明崗, 張文菊, 李兆強, 張敬業. 長期施肥對紅壤微生物生物量碳氮和微生物碳源利用的影響. 應用生態學報, 2010, 21(11): 2792- 2798.
[2] 逄好勝, 張會慧, 李鑫, 丁偉鵬, 胡舉偉, 林晗婧, 敖紅, 孫廣玉. 大興安嶺林區森林退化對土壤微生物群落功能的影響. 森林工程, 2015, 31(3): 25- 30.
[3] 田雅楠, 王紅旗. Biolog法在環境微生物功能多樣性研究中的應用. 環境科學與技術, 2011, 34(3): 50- 57.
[4] 張海芳, 李剛, 宋曉龍, 劉紅梅, 張靜妮, 楊殿林, 趙樹蘭, 多立安. 內蒙古貝加爾針茅草原不同利用方式土壤微生物功能多樣性. 生態學雜志, 2012, 31(5): 1143- 1149.
[5] Elser J J, Bracken M E S, Cleland E E, Gruner D S, Harpole W S, Hillebrand H, Ngai J T, Seabloom E W, Shurin J B, Smith J E. Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems. Ecology Letters, 2007, 10(12): 1135- 1142.
[6] 楊曉霞, 任飛, 周華坤, 賀金生. 青藏高原高寒草甸植物群落生物量對氮、磷添加的響應. 植物生態學報, 2014, 38(2): 159- 166.
[7] 王志勇. 測土配方施肥理論與實踐. 鄭州: 黃河水利出版社, 2010: 5- 10.
[8] 郁培義, 朱凡, 宿少鋒, 王志勇, 閆文德. 氮素添加對樟樹林紅壤微生物的影響. 環境科學, 2013, 34(8): 3231- 3237.
[9] 陳超凡, 吳水榮, 覃林, 范垚城, 譚玲, 郭文福. 南亞熱帶典型鄉土闊葉人工林土壤微生物碳源利用及功能多樣性. 生態學雜志, 2016, 35(5): 1132- 1139.
[10] Gomez E, Ferreras L, Toresani S. Soil bacterial functional diversity as influenced by organic amendment application. Bioresource Technology, 2006, 97(13): 1484- 1489.
[11] 楊永華, 姚健, 華曉梅. 農藥污染對土壤微生物群落功能多樣性的影響. 微生物學雜志, 2000, 20(2): 23- 25.
[12] 韓冬雪, 王寧, 王楠楠, 孫雪, 馮富娟. 不同海拔紅松林土壤微生物功能多樣性. 應用生態學報, 2015, 26(12): 3649- 3656.
[13] 吳家森, 錢進芳, 童志鵬, 黃堅欽, 趙科理. 山核桃林集約經營過程中土壤有機碳和微生物功能多樣性的變化. 應用生態學報, 2014, 25(9): 2486- 2492.
[14] 隋躍宇, 焦曉光, 高崇生, 程偉, 張興義, 劉曉冰. 土壤有機質含量與土壤微生物量及土壤酶活性關系的研究. 土壤通報, 2009, 40(5): 1036- 1039.
[15] 張彥東, 孫志虎, 沈有信. 施肥對金沙江干熱河谷退化草地土壤微生物的影響. 水土保持學報, 2005, 19(2): 88- 91.
[16] 李娟, 趙秉強, 李秀英, Bo H. B . 長期有機無機肥料配施對土壤微生物學特性及土壤肥力的影響. 中國農業科學, 2008, 41(1): 144- 152.
[17] 魯順保, 張艷杰, 陳成榕, 徐志紅, 郭曉敏. 基于BIOLOG指紋解析三種不同森林類型土壤細菌群落功能差異. 土壤學報, 2013, 50(3): 618- 623.
[18] 陳志芳. 戴云山不同森林類型土壤微生物群落多樣性特征的研究[D]. 福州: 福建農林大學, 2014.
[19] Bach L H, Grytnes J A, Halvorsen R, Ohlson M. Tree influence on soil microbial community structure. Soil Biology and Biochemistry, 2010, 42(11): 1934- 1943.
[20] 韓世忠, 高人, 李愛萍, 馬紅亮, 尹云鋒, 司友濤, 陳仕東, 鄭群瑞. 中亞熱帶地區兩種森林植被類型土壤微生物群落結構. 應用生態學報, 2015, 26(7): 2151- 2158.
[21] 劉秉儒, 張秀珍, 胡天華, 李文金. 賀蘭山不同海拔典型植被帶土壤微生物多樣性. 生態學報, 2013, 33(22): 7211- 7220.
[22] 宋賢沖, 楊中寧, 曹繼釗, 張照遠, 項東云. 萌芽更新對桉樹根際土壤微生物群落功能多樣性的影響. 桉樹科技, 2014, 31(3): 36- 40.
[23] 宋春雨, 張興義, 劉曉冰, 高崇升. 土壤有機質對土壤肥力與作物生產力的影響. 土壤與作物, 2008, 24(3): 357- 362.
[24] 李娟, 趙秉強, 李秀英, 姜瑞波, Bo H. B. 長期不同施肥制度下幾種土壤微生物學特征變化. 植物生態學報, 2008, 32(4): 891- 899.
[25] 吳則焰, 林文雄, 陳志芳, 方長旬, 張志興, 吳林坤, 周明明, 陳婷. 中亞熱帶森林土壤微生物群落多樣性隨海拔梯度的變化. 植物生態學報, 2013, 37(5): 397- 406.
[26] 董立國, 蔣齊, 蔡進軍, 張源潤, 許浩, 李生寶. 基于Biolog-ECO技術不同退耕年限苜蓿地土壤微生物功能多樣性分析. 干旱區研究, 2011, 28(4): 630- 637.
[27] Li F L, Liu M, Li Z P, Jiang C Y, Han F X, Che Y P. Changes in soil microbial biomass and functional diversity with a nitrogen gradient in soil columns. Applied Soil Ecology, 2013, 64: 1- 6.
猜你喜歡
中等數學(2022年2期)2022-06-05 07:10:50
中學生數理化·七年級數學人教版(2021年11期)2021-12-06 05:38:48
中學生數理化(高中版.高考數學)(2021年6期)2021-07-28 06:19:08
小學生學習指導(低年級)(2020年6期)2020-07-25 02:31:36
小學生學習指導(低年級)(2019年11期)2019-11-25 07:31:44
小學生學習指導(低年級)(2018年9期)2018-09-26 05:59:44
瘋狂英語·新讀寫(2018年2期)2018-09-07 09:32:10
數學小靈通·3-4年級(2017年6期)2017-06-22 11:28:50
工業設計(2016年5期)2016-05-04 04:00:33
河北遙感(2015年4期)2015-07-18 11:05:06
