外源調節劑包衣對低溫脅迫下棉花種子萌發及幼苗耐寒性的影響
2017-03-02 08:43:37李防洲冶軍侯振安
干旱地區農業研究 2017年1期
李防洲,冶軍,侯振安
(新疆生產建設兵團綠洲生態農業重點實驗室石河子大學農學院,新疆石河子832003)
外源調節劑包衣對低溫脅迫下棉花種子萌發及幼苗耐寒性的影響
李防洲,冶軍,侯振安
(新疆生產建設兵團綠洲生態農業重點實驗室石河子大學農學院,新疆石河子832003)
為探討外源調節劑包衣對低溫脅迫棉花幼苗耐寒性的調控效應,通過室內砂培試驗,研究了外源調節劑包衣處理對低溫下棉花種子萌發的影響,分析了5℃低溫脅迫下以及恢復常溫后外源調節劑包衣對棉花幼苗耐寒性的生理響應。結果表明:低溫脅迫下棉花種子的萌發和幼苗生長受到抑制,水楊酸以及外源調節劑復配包衣處理均能顯著提高低溫脅迫下棉花種子的發芽勢、發芽率、發芽指數和活力指數,其中發芽指數和活力指數達到極顯著水平,增幅分別為41.2%~44.4%和51.2%~63.9%;外源調節劑包衣處理葉片的相對電導率(REC)和MDA含量顯著降低,可溶性蛋白和可溶性糖等滲透調節物質的含量顯著提高,SOD、POD和CAT等抗氧化酶的活性也顯著增強;5mmol·L-1水楊酸+45mmol·L-1氯化鈣+60mg·L-1亞硒酸鈉復配包衣處理的葉片REC、MDA含量顯著低于其它包衣處理,可溶性蛋白含量及其增幅顯著高于其它包衣處理。說明水楊酸、氯化鈣和亞硒酸鈉三元復配包衣在緩解棉花幼苗低溫逆境中具有協同效應,可以緩解低溫脅迫對棉花種子萌發及幼苗生長的抑制作用,提高植物的耐寒性。
棉花;種子萌發;外源調節劑包衣;低溫脅迫;幼苗耐寒性
低溫是影響作物產量的重要環境因子之一,也是限制農業生產發展的主要自然災害之一[1],全球每年因低溫危害造成的農作物損失高達數千億元[2]。近些年來,隨著全球氣溫的升高,極端氣候頻現,尤其在棉花(Gossypium hirsutum)等喜溫作物的播種和幼苗生長期間經常遭遇連雨雪、寒潮等低溫氣象災害[3-5],從而導致棉花爛種、爛芽、弱苗甚至死苗,嚴重影響植株生長發育和產量品質形成[6],最終造成減產減收。種子包衣以種子為載體,以包衣設備為手段,將包衣劑按照一定比例均勻地包敷到種子表面的處理技術,在作物增產等方面具有很大的作用。阿里甫·艾爾西等[7]兩年的試驗研究表明,抗低溫種衣劑對低溫造成的爛種、爛芽具有明顯防治效果,在棉花生產中使用棉花抗低溫種衣劑具有明顯的增產效果。
目前關于植物耐寒性機理的研究表明,植物在低溫脅迫下會發生一系列生理生化紊亂,首先細胞內活性氧水平升高,誘發脂膜過氧化鏈式反應,導致細胞膜完整性破壞、透性增加,繼而葉片青枯死亡[8]。Suzuki N等[9]研究表明,低溫脅迫下,MDA含量增加,而SOD、POD等酶活性增強,以清除逆境引起的活性氧,降低其對膜蛋白的影響。已有研究表明一些外源調節劑(水楊酸、氯化鈣、亞硒酸鈉等)參與調節植物的許多生理生化過程。水楊酸(Salicylic acid,SA)作為誘導植物對非生物逆境反應的抗逆信號分子[10],通過減輕膜脂過氧化、提高滲透物質含量、提高抗氧化酶活性等途徑來提高玉米[11]、黃瓜[12]、大豆[13]等的耐寒性。鈣不僅是一種植物必需的大量元素,而且作為偶聯胞內信號與胞外生理生化的第二信使,在植物逆境生理生態中發揮著增強植物生理功能的作用[14]。Rickauer等[15]研究表明鈣能維持細胞壁、細胞膜及膜結合蛋白的穩定性。由繼紅等[16]、廖金柯等[17]研究表明鈣處理能顯著緩解植物幼苗的低溫傷害。硒作為植物生長發育的有益元素,現有研究表明,適量硒有清除過量自由基的功能,在一定程度上硒能緩解脅迫對植物的損傷[18]。
有關外源調節物在提高植物耐寒性方面的研究已有較多的研究且多集中于浸種或葉面噴施,而通過種子包衣處理技術,探討能夠調節植物生理機制的調節劑,在改善植物生理過程與低溫脅迫的關系、提高抗低溫脅迫能力方面的生理機制報道較少。為此,本文通過前期預備試驗,優選水楊酸、氯化鈣和亞硒酸鈉三種外源調節劑及其復配包衣處理研究外源調節劑包衣處理對棉花種子萌發及幼苗耐寒性的調控效應,為農業生產上使用簡單的化學調控和包衣技術相結合提高棉花對低溫冷害的耐受性以及在生產中的應用提供理論依據。
1 材料與方法
1.1 試驗材料
供試棉花品種為生產上大面積推廣的新陸早48號,由石河子棉花研究所提供;水楊酸、氯化鈣由天津市盛奧化學試劑有限公司生產;亞硒酸鈉由天津市光復精細化工研究所生產;基礎種衣劑為“錦華種衣劑”,由新疆錦華農藥有限公司生產。
1.2 試驗方法
1.2.1 種子包衣處理試驗共設5個處理,(1)基礎種衣劑包衣處理;(2)5 mmol·L-1水楊酸包衣處理;(3)5 mmol·L-1水楊酸+45 mmol·L-1氯化鈣包衣處理;(4)5 mmol·L-1水楊酸+60 mg·L-1亞硒酸鈉包衣處理;(5)5 mmol·L-1水楊酸+45 mmol·L-1氯化鈣+60 mg·L-1亞硒酸鈉包衣處理,依次記為CK、T1、T2、T3、T4。挑選均勻一致的棉花種子,將其外源調節劑添加到基礎種衣劑中,配置成以上不同濃度的外源包衣劑,以基礎種衣劑包衣處理作為對照(CK),按藥種比1∶40進行種子包衣處理,將包好的棉花種子放在通風干燥處陰干進行萌發和早期幼苗生長試驗。
1.2.2 種子萌發試驗模擬中國北方春季氣溫變化,設置5℃~20℃棉花種子萌發條件。采用20 cm ×14 cm×5 cm的發芽盒,用洗凈經高壓蒸汽滅菌消毒后的濕潤河沙做發芽床。試驗選擇以上包衣處理的種子,每個處理3次重復,每盒40粒種子,種子整齊置床后上覆約1.5 cm的濕沙,擺放于溫度為5℃培養箱處理24 h,然后于晝夜溫度20℃/10℃、光暗周期為12 h/12 h的培養箱繼續培養。萌發過程中,以胚芽露土為出苗,逐日觀察記錄萌發種子數。第7天測定發芽勢,第14天測定發芽率。
1.2.3 幼苗培養及低溫處理采用盆栽試驗,基質為洗凈的河沙,將洗凈的濕沙裝入盆缽中,每個處理播種5盆,每盆播種20粒,播種后上覆約2 cm的濕沙。在RXZ型智能人工氣候箱進行培養,光暗周期為14 h/10 h,晝/夜溫度25℃/25℃。待子葉展平后定期澆灌定量Hoagland營養液,一葉一心期定苗,每盆留苗15株,培養至兩葉一心時,挑選生長一致的幼苗進行低溫脅迫試驗。先在常溫下培養1 d,然后在5℃低溫下脅迫處理2 d,之后恢復常溫培養1 d(R1d)。分別于低溫脅迫0 d(即常溫下)、1 d、2 d以及恢復常溫1 d(R1d)時采樣,測定相關生理生化指標。
1.3 測定項目及方法
1.3.1 種子萌發參數測定發芽勢=第7天的發芽種子數/供試種子數×100%,發芽率=第14天的發芽種子數/供試種子數×100%。發芽指數(GI)=∑(Gt/Dt),式中Gt為t日內的發芽數,Dt為相應的發芽天數。活力指數(VI)=GI×S,式中GI為發芽指數,S為單株幼苗干物質質量(g),用稱量法測定干物質量。
1.3.2 生理指標的測定相對電導率(REC)采用DDS-307型電導率儀測定[19];丙二醛(MDA)含量的測定參照高俊鳳的方法用硫代巴比妥酸(TBA)法[19]測定;可溶性蛋白含量采用考馬斯亮藍G-250法[20]測定;可溶性糖含量采用蒽酮比色法[20]測定;抗氧化酶活性測定采用陳建勛[21]的方法稍加修正,酶液提取:準確稱取棉花幼苗葉片樣品0.5 g于預冷的研缽中,加1 ml 0.05 mol·L-1、pH=7.8的磷酸緩沖液在冰浴上研磨成漿,加緩沖液使終體積為5 ml。將提取液于10 000 r·min-1冷凍(4℃)離心20 min,上清液即為粗提液,然后將上清液倒入試管中0~4℃下保存,用于測定超氧化物歧化酶(SOD)、過氧化物酶(POD)以及過氧化氫酶(CAT)活性。
1.4 數據分析
采用Excel 2003進行數據處理及作圖,運用SPSS 19.0進行單因素方差分析及差異顯著性檢驗。圖表中數據為平均值±標準差。
2 結果與分析
2.1 外源調節劑包衣處理對低溫脅迫下棉花種子萌發的影響
由表1可知,在低溫條件下,外源調節劑包衣處理棉花種子的發芽勢、發芽率與CK差異顯著,外源調節劑包衣處理間差異不顯著。T2、T3和T4處理對棉花種子的發芽指數、活力指數較CK差異達到顯著水平(P<0.05),其發芽指數分別提高了41.2%、41.7%和44.4%,活力指數分別提高了62.4%、51.2%和63.9%。說明外源調節劑包衣對低溫脅迫下棉花種子的萌發起到了促進作用。

表1 外源調節劑包衣處理對棉花種子萌發的影響Table 1 Effectof seed film coating with exogenous regulating substances on seed germination of cotton
2.2 外源調節劑包衣處理對低溫脅迫下棉花幼苗細胞膜系統的影響
圖1表明,低溫脅迫前,與CK相比,外源調節劑包衣處理不同程度地降低了REC和MDA含量。隨低溫脅迫時間的延長REC和MDA含量不斷升高,外源調節劑包衣處理與CK相比顯著降低葉片的REC和MDA含量(P<0.05),低溫脅迫1 d,降幅分別為15.8%~27.2%和10.8%~24.5%。低溫脅迫2 d,REC和MDA含量達到峰值,T4處理的葉片REC和MDA與CK相比分別降低25.5%和29.2%,差異達到極顯著。恢復常溫后,T3和T4處理間棉花葉片REC和MDA含量差異不顯著且均呈現下降趨勢,各處理的REC和MDA含量均高于低溫脅迫前。說明低溫脅迫使幼苗葉片細胞膜透性增大,膜脂過氧化程度加劇,而外源調節物質包衣處理葉片中REC和MDA含量較CK明顯降低,從而緩解低溫對棉花幼苗的傷害。
2.3 外源調節劑包衣處理對低溫脅迫下棉花幼苗滲透調節系統的影響
由圖2可以看出,低溫脅迫前后,可溶性蛋白和可溶性糖含量呈先增后減的趨勢,其中可溶性蛋白含量在低溫脅迫2 d達到最大值,T2和T4包衣處理的可溶性糖含量在脅迫1 d達到峰值。說明外源調節劑包衣處理對調節低溫脅迫下葉片可溶性蛋白和可溶性糖可能有不同的作用機制。T1~T4處理均能顯著提高低溫脅迫后葉片中可溶性蛋白和可溶性糖含量(P<0.05),T4處理與CK相比,葉片中可溶性蛋白和可溶性糖含量在低溫脅迫1 d時提高35.8%和48.2%,脅迫2 d時提高27.8%和28.1%,恢復常溫后提高20.7%和23.9%。表明外源調節劑包衣能顯著增加可溶性有機物的積累,增強了低溫脅迫下的滲透調節能力,且以T4包衣處理效果最好。

圖1 外源調節劑包衣處理對低溫脅迫下棉花幼苗細胞膜系統的影響Fig.1 Effects of seed film coatingwith exogenous regulating substances onmembrane systems of cotton seedling under low temperature stress
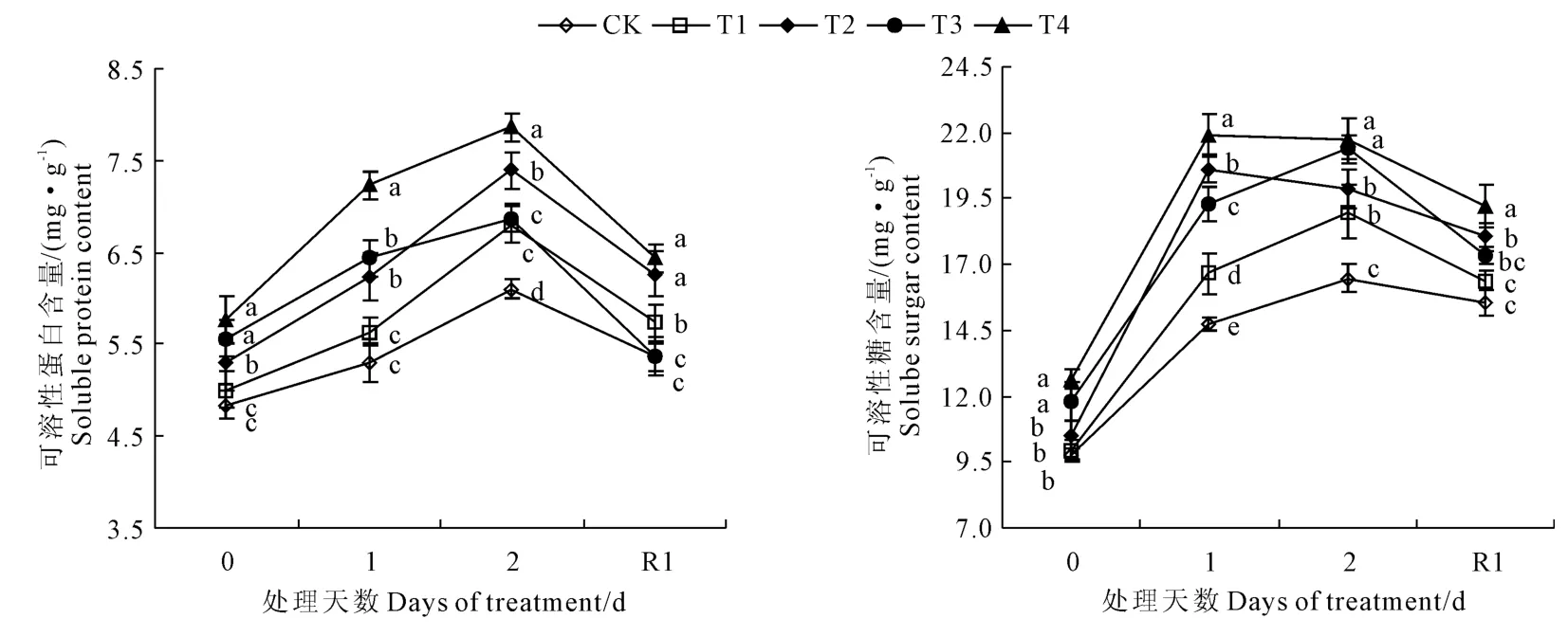
圖2 外源調節劑包衣處理對低溫脅迫下棉花幼苗滲透調節物質含量的影響Fig.2 Effects of seed film coating with exogenous regulating substances on osmotic regulation substances of cotton seedling under low temperature stress
2.4 外源調節劑包衣處理對低溫脅迫下棉花幼苗抗氧化酶系統的影響
從圖3可以看出,低溫脅迫下,與CK相比,T1~T4處理均能顯著提高棉花幼苗葉片中SOD、POD和CAT活性。POD和CAT活性分別在低溫脅迫2 d和1 d時出現最大值,其中T3和T4與CK相比,差異均達極顯著水平,但T3和T4處理間差異不顯著。低溫脅迫1 d時與CK相比,T4處理的棉花葉片SOD活性提高16.6%,T3處理的棉花葉片CAT活性提高28.3%。低溫脅迫2 d時,T3和T4處理的棉花葉片SOD活性與CK相比分別提高10.2%和11.6%,POD活性分別提高12.7%和14.0%。恢復常溫處理,POD和CAT活性在T1~T4處理之間差異不顯著。說明外源調節劑包衣處理可以提高棉花幼苗葉片抗氧化酶活性,增強植株的抗寒性。
3 討論
3.1 外源調節劑包衣處理對低溫脅迫下棉花種子萌發的影響
植物整個生長周期中種子萌發作為生理代謝最旺盛的階段,容易受到逆境刺激。棉花生產上,為防止或降低由于播期提前和早春低溫,造成的爛種、爛芽問題,衛秀英等[22]使用抗低溫種衣劑包衣的棉花種子,結果棉花種子在低溫條件下也保持較高的活力,有效降低低溫冷害對出苗帶來的危害。本研究表明,在低溫條件下,棉種萌發期各項指標都有不同程度降低,說明低溫對棉花種子萌發有抑制效應,而用生理調節劑包衣處理的棉種與基礎包衣相比,不同程度地顯著提高了棉種的發芽勢、發芽率,發芽指數和活力指數增幅分別為33.1%~44.4%和51.5%~63.9%。

圖3 外源調節劑包衣處理對低溫脅迫下棉花幼苗抗氧化酶活性的影響Fig.3 Effects of seed film coatingwith exogenous regulating substances on antioxidant enzyme activities of cotton seedling under low temperature stress
3.2 低溫脅迫對棉花幼苗抗寒性的作用機制
植物在低溫等逆境條件下細胞內自由基代謝平衡被破壞而使自由基不斷增加,引發或加劇膜脂過氧化,導致細胞膜結構和功能遭到破壞,膜脂過氧化產物MDA含量增加[23],進而引起一系列生理生化代謝紊亂,導致傷害發生[24]。為了避免或減輕自由基對細胞造成傷害,植物體內可溶性蛋白質和可溶性糖等滲透調節物質的含量增加,從而降低細胞滲透勢,增強植物自身適應逆境的能力[20],同時植物細胞形成相應的酶促保護系統(SOD、POD和CAT)[25]。本研究為證明低溫脅迫下外源調節劑包衣降低REC是否由于減少了膜系統的氧化傷害水平,對葉片抗氧化物質含量進行測定表明,低溫脅迫使棉花幼苗細胞膜的透性發生改變,進而使REC升高,同時產生過多的自由基,導致活性氧增加,MDA含量明顯增加,使抗氧化酶活性下降,細胞的膜結構和功能受到影響,從而使棉花幼苗代謝途經發生紊亂。
3.3 外源調節劑包衣處理對低溫脅迫下棉花幼苗抗寒性的影響
逆境不僅提高細胞活性氧水平,同時也誘發植物防御體系的建立,從而避免或減輕活性氧對植物的傷害。多數研究認為,外源調節劑在緩解植物逆境脅迫中具有重要作用。劉忠國等[26]對黃瓜幼苗耐熱性的研究發現水楊酸和氯化鈣的復配制劑具有拮抗性。而劉雪琴等[27]研究表明水楊酸和硝酸鈣復配處理對鹽脅迫下玉米幼苗具有緩解作用。辛慧慧等[28]研究表明水楊酸、殼聚糖和硝酸鈣在誘導棉花幼苗耐寒性中具有協同效應且水楊酸、殼聚糖、硝酸鈣三元復配的效果比水楊酸和殼聚糖兩元復配的效果好。本試驗研究表明,將種子包衣技術和化控技術相結合,通過研究低溫脅迫對棉花幼苗生理生化特性的影響,5 mmol·L-1水楊酸包衣處理棉花幼苗葉片的REC和MDA含量及SOD、POD、CAT活性與CK差異不顯著;低溫脅迫后,外源調節劑包衣處理棉花幼苗葉片的REC和MDA含量顯著降低,可溶性蛋白和可溶性糖含量以及抗氧化酶SOD、POD、CAT活性均顯著高于CK;恢復常溫后,REC和MDA含量,可溶性蛋白和可溶性糖含量以及抗氧化酶SOD、POD、CAT活性均有所下降,但外源調節劑包衣處理幼苗葉片的REC和MDA含量始終低于對照,而抗氧化酶SOD、POD、CAT活性均顯著高于對照,其中以5 mmol·L-1水楊酸+45 mmol·L-1氯化鈣+60 mg·L-1亞硒酸鈉包衣處理對緩解幼苗耐寒性具有顯著效果。
4 結論
1)外源調節劑包衣處理是一種安全、有效、經濟、以抗低溫制劑和種衣劑為一體的、可促進棉花生長,增強種子抗逆能力的一種包衣劑,在種子萌發過程中,各種外源調節劑以及營養成分緩慢釋放,通過誘導細胞內部酶的活性,促進萌發期種子內營養物質的轉化與合成,同時還誘導抗寒物的合成,從而激活棉種活力,增強了棉花萌發期的抗寒性。
2)低溫脅迫使棉花幼苗細胞膜的透性發生改變,進而使REC升高,同時產生過多的自由基,導致活性氧增加,MDA含量明顯增加,使抗氧化酶活性下降,細胞的膜結構和功能受到影響,從而使棉花幼苗代謝途徑發生紊亂。
3)低溫脅迫下,水楊酸、氯化鈣和亞硒酸鈉三元復配包衣和二元包衣處理的可溶性糖含量和SOD出現不同的峰值,說明不同外源調節劑對調控棉花幼苗逆境脅迫可能有不同的機制,也揭示了外源調節劑在調節棉花幼苗體內滲透調節物質含量和抗氧化酶活性水平等方面的復雜性。外源調節劑包衣通過激活棉花種子生物酶活性,從而提高細胞膜結構和功能的穩定性和細胞質濃度,從根本上可提高棉花對低溫冷害等不利環境的適應與抵抗能力,從而表明水楊酸、氯化鈣和亞硒酸鈉在緩解棉花幼苗耐寒性中具有協同效應。然而,有關外源調節劑緩解棉花幼苗低溫脅迫是一個復雜的調控機制,還待做進一步研究。
[1]劉祖祺,張石城.植物抗寒分子生物學研究進展[J].南京農業大學學報,1993,16(1):113-120.
[2]Guyic C L.Cold accelimation and freezing stress tolerance:Role of proteinmetabolism[J].Annual Review of Plant Physiology and Plant Molecular Biology,1990,41:187-223.
[3]胡汝驥,樊自立,王亞俊,等.近50年新疆氣候變化對環境影響評估[J].干旱區地理,2001,24(2):97-103.
[4]賴先齊,劉月蘭,徐臘梅,等.北疆棉區棉花低溫冷害的初步分析及對策探討[J].新疆農業科學,2008,45(5):782-786.
[5]賈宏濤,趙成義,巴特爾·巴克,等.新疆氣候變化影響的觀測事實及其對農牧業生產的影響[J].干旱區資源與環境,2009,23(11):72-76.
[6]趙強,徐臘梅,王磊,等.2006年5月上旬新疆石河子棉區低溫危害分析[J].新疆農業科學,2007,44(1):23-26.
[7]阿里甫·艾爾西,孫良斌,張少民,等.棉花抗低溫種衣劑田間篩選及效果分析[J].種子,2002,31(3):90-92.
[8]Boyer JS.Plant productivity and environment[J].Science,1982,218:443-448.
[9]Suzuki N,Mittler R.Reactive oxygen species and temperature stresses:a delicate balance between signaling and destruction[J].Physiol Plant,2006,126:45-51.
[10]Agarwal S,Sairam R K,Srivasa GC,et al.Changes in antioxidant enzymes activity and oxidative stress by abscisci acid and salicylic acid in wheat genotypes[J].Biologia Plantarum,2005,49(4):541-550.
[11]張富平,張蕊.低溫下外源水楊酸對玉米幼苗保護酶活性的影響[J].玉米科學,2007,15(4):83-85.
[12]劉偉,艾希珍,梁文娟,等.低溫弱光下水楊酸對黃瓜幼苗光合作用及抗氧化酶活性的影響[J].應用生態學報,2009,20(2):441-445.
[13]常云霞,徐克東,陳璨,等.水楊酸對低溫脅迫下大豆幼苗生長抑制的緩解效應[J].大豆科學,2012,31(6):927-931.
[14]Batistic O,Kudla J.Calcium:Not just another ion[C]//Hell R,Mendel R R.Cell Biology of Metals and Nutrients.Berlin,Heidelberg:Springer-Verlag,2010:17-54.
[15]Rickauer M,TanerW.Effects of Ca2+on amino acid transport and accumulation in roots of phaseolus vularis[J].Plant Physiology,1986,82(1):41-46.
[16]由繼紅,陸靜梅,楊文杰.鈣對低溫脅迫下小麥幼苗光合作用及相關生理指標的影響[J].作物學報,2002,28(9):693-696.
[17]廖金柯,朱新霞,胡小燕,等.低溫脅迫下棉花幼苗對外源鈣的生理響應[J].西北農業學報,2013,22(2):60-64.
[18]Feng R,Wei C,Tu S.The roles of selenium in protecting plants against abiotic stresses[J].Environmental and Experimental Botany,2012,58(3):105-110.
[19]高俊鳳.植物生理學實驗指導[M].北京:高等教育出版社,2006.
[20]張治安,陳展宇.植物生理學實驗技術[M].吉林:吉林大學出版社,2008.
[21]陳建勛,王曉峰.植物生理學實驗指導[M].廣州:華南理工大學出版社,2006.
[22]衛秀英,魯玉貞,單長卷.不同棉花品種的抗低溫性研究[J].安徽農業科學,2006,(12):2786-2787.
[23]Jane L,Bingru H.Changes of lipid composition and saturation level in leaves and roots for heat-stressed and heat-acclimated creeping bentgrass(Agrostis stolomiferɑ)[J].Environ.Exp.Bot,2004,51(1):57-67.
[24]Lagriffoul A,Mocquot B,Mench M,et al.Cadmium toxicity effects on growth,mineral and chlorophyll contents,and activities of stress related enzymes in youngmaize plant(Zeɑmɑys L.)[J].Plant and Soil,1998,200:241-250.
[25]Tsang EW T,Bowler C,HerouartD,etal.Differential regulation of superoxidedismutases in plants exposed to environmental stress[J].Plant Cell,1991,3:783-792.
[26]劉忠國,曹辰興,王濤,等.水楊酸與氯化鈣單一及復配誘導對黃瓜幼苗耐熱性的影響[J].山東農業科學,2010,4(4):27-30.
[27]劉雪琴,李勇慧,仝瑞建,等.硝酸鈣和水楊酸復配處理對鹽脅迫下玉米幼苗生長的影響[J].廣東農業科學,2011,16(1):11-12.
[28]辛慧慧,李志強,李防洲,等.外源調節物質對棉花幼苗耐寒生理特性的效應[J].棉花學報,2015,27(3):254-259.
Effects of seed film coating w ith exogenous regulating substances on cotton germ ination rate and cold tolerance of seedlings
LIFang-zhou,YE Jun,HOU Zhen-an
(The Key Lɑborɑtory of Oɑsis Eco-ɑgriculture of Xinjiɑng Production Construction Group/College of Agriculture,Shihezi University,Shihezi,Xinjiɑng 832003,Chinɑ)
The aim of this experimentwas to study the effects of seed film coating with exogenous regulating substances on the cotton seed germination and physiological response of cold tolerance in cotton seedlings under low temperature stress of 5℃by sand culturemethod.The results showed that the low temperature stress inhibited the cotton seed germination and growth of seedlings.Seeds coated with salicylic acid and exogenous regulator compound had better germination potential,and higher germination rate,germination index and vigor index.Compared with CK,seeds coated with exogenous regulator compound had germination index increased by 41.2%~44.4%,vigor index 30.9%~63.9%.The coating treatments with exogenous regulating substances decreased leaf REC and MDA content.It also increased sharply the soluble protein and soluble sugar contents.The activity of SOD,POD and CATwere significantly increased.The coating treatmentwith a compound preparation of5mmol·L-1salicylic acid,45mmol·L-1calcium chloride and 60 mg·L-1sodium selenitewhich REC and MDA contentwere significantly higher than other coating treatment,and soluble protein contentwere significantly higher than other coating treatment.These results demonstrated that seed film coating with salicylic acid,calcium chloride and sodium selenite has a synergistic effect of improving low temperature resistance of cotton seedling by promoting seed germination and seedling growth.
cotton;germination rate;film coating with exogenous regulating substances;low temperature stress;cold resistance of seedlings
S562
:A
1000-7601(2017)01-0192-06
10.7606/j.issn.1000-7601.2017.01.29
2016-01-10
國家科技支撐計劃“低溫災害防控關鍵技術研究與應用”項目(2012BAD20B03)
李防洲(1987—),男,甘肅天水人,碩士研究生,主要從事植物營養生理生態研究。E-mail:tianshuilifangzhou@163.com。
冶軍(1974—),男,山東牟平人,副教授,主要從事新型肥料與現代施肥技術。E-mail:yejun.shz@163.com。
