全基因組預測希金斯炭疽菌中碳水化合物酶類蛋白
2017-03-01 05:53:53韓長志
江蘇農業科學 2017年2期
關鍵詞:分析
韓長志
(西南林業大學林學院/云南省森林災害預警與控制重點實驗室,云南昆明 650224)
全基因組預測希金斯炭疽菌中碳水化合物酶類蛋白
韓長志
(西南林業大學林學院/云南省森林災害預警與控制重點實驗室,云南昆明 650224)
希金斯炭疽菌侵染菜心等十字花科植物引起的炭疽病,給各國農業生產造成了巨大的經濟損失。基于前期研究結果,以658個分泌蛋白為基礎序列,利用CAZymes Analysis Toolkit預測程序,分析上述蛋白中的碳水化合物酶類(CAZymes)蛋白,明確該菌中含有238個CAZymes,分為主要類別和復合類別2類,前者包括75個糖苷水解酶(GHs)、48個碳水化合物綁定結構(CBMs)、33個輔助酶類家族(AAs)、30個碳水化合物酯酶(CEs)、23個多糖裂解酶(PLs)、4個糖基轉移酶(GTs),后者則包括17個GHs/CBMs、4個AAs/CBMs、4個CEs/CBMs等。研究結果可為深入開展該病菌侵染植物作用機制的研究提供一定的理論基礎。
希金斯炭疽菌;碳水化合物酶類蛋白;預測程序;全基因組預測
碳水化合物酶類(carbohydrate-active enzymes,簡稱CAZymes)是植物病原菌侵染過程中突破寄主細胞第一道屏障——細胞壁的關鍵因素[1]。同時,對不同營養類型的植物病原真菌進行分析,可以明確非活體營養型病原菌基因組較活體營養型編碼更多的CAZymes[2]。隨著大量植物病原菌基因組測序的完成,近些年關于CAZymes的研究進展較快[3]。根據最新研究成果,以涵蓋木質纖維素降解所需要的相關酶類進行分類,主要涉及以下6類:糖苷水解酶(glycoside hydrolases,簡稱GHs)[4]、糖基轉移酶(glycosyl transferases,簡稱GTs)[5]、多糖裂解酶(polysaccharide lyases,簡稱PLs)[6]、碳水化合物酯酶(carbohydrate esterases,簡稱CEs)[6]、輔助酶類家族(auxiliary activities,簡稱AAs)[7]以及碳水化合物綁定結構(carbohydrate-binding modules,簡稱CBMs)[8]。
希金斯炭疽菌(ColletotrichumhigginsanumSacc.)作為半活體營養型植物病原菌,可以侵染菜心、白菜、羽衣甘藍、大白菜、蘿卜、結球甘藍等十字花科蔬菜引起炭疽病[9-10],是農業生產上較為嚴重的病害之一[11]。國內外學者對其生物學特性、生防菌篩選以及遺傳轉化[12]、致病基因[13-14]、轉化子的突變體庫建立[15-16]、防治[17]、絲裂原活化蛋白激酶(MAPK)級聯信號通路繪制[18],以及G蛋白信號調控因子(RGS)[19]、磷脂酰肌醇轉移蛋白(PITP)[20]、G蛋白偶聯受體(GPCR)[21]、腺苷酸環化酶及其相關蛋白[22-23]、14-3-3[24]、胞裂蛋白(septin)[25]、磷酸二酯酶(PDE)[26]等在內的諸多蛋白進行了生物信息學分析。
本研究基于前期所獲得的希金斯炭疽菌中658個分泌蛋白,利用CAZymes Analysis Toolkit(CAT)預測工具明確該菌中CAZymes不同亞家族的分布情況,以期為深入開展其功能研究打下堅實的基礎。
1 材料與方法
1.1 希金斯炭疽菌分泌蛋白序列來源
希金斯炭疽菌分泌蛋白,來自筆者在前期研究中獲得的658個分泌蛋白[27]。
1.2 CAZymes預測
利用在線工具CAZymes Analysis Toolkit[28]進行預測。
2 結果與分析
2.1 希金斯炭疽菌中含有的CAZymes
通過CAT分析,明確希金斯炭疽菌中共含有238個CAZymes,其中GHs含量較高,所占比例為31.51%;其后依次為CBMs、AAs、CEs、PLs、GTs,所占比例分別為20.17%、13.87%、12.61%、9.66%、1.68%(圖1)。此外,在上述CAZymes的預測中,不僅具有單一的酶類型,而且有部分蛋白同屬于2種酶的類型,涉及GHs/CBMs、AAs/CBMs、CEs/CBMs 等(圖1)。由于CAZymes分類系統的理論基礎是建立在氨基酸序列的相似性上, 不同蛋白具有不同的結構折疊類型,因此,對于同一蛋白具有不同酶類型的情況,推測病原菌在侵染植物過程中上述酶所發揮的功能具有較多特性。

2.2 希金斯炭疽菌中含有的GHs
通過對上述糖基水解酶進行分析,發現GH28中蛋白數量較多,為9個,其次為GH16,數量為7個(表1)。值得關注的是,GH105、GH128、GH132、GH20、GH30等類別的酶均只含有1個蛋白,推測希金斯炭疽菌侵染不同植物所利用的酶并不相同,也從分泌酶角度可以解釋其寄主廣泛的原因。然而,對于為什么有些酶類所含數量較多,而大多數所含數量較少,有待進一步研究。
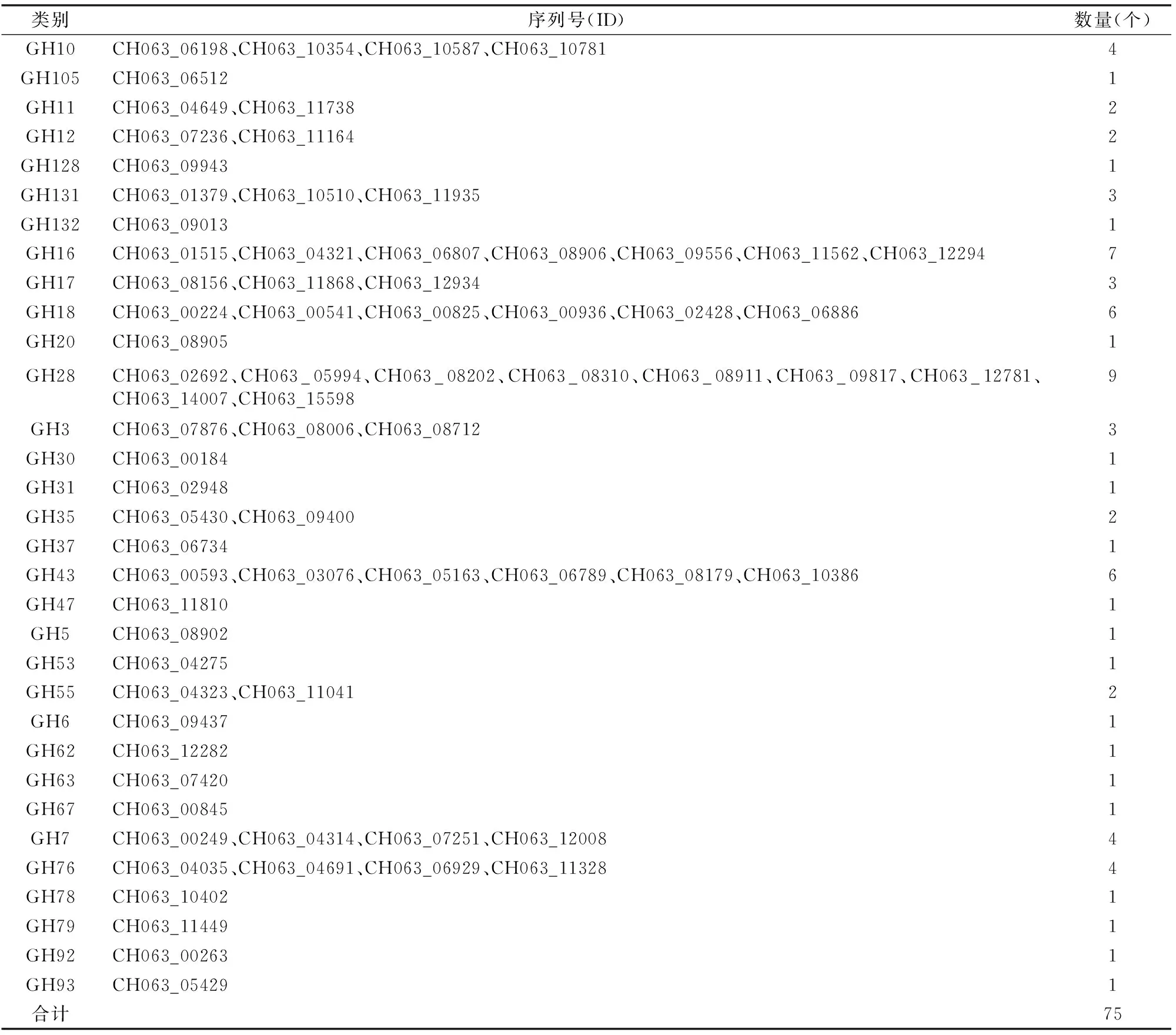
表1 希金斯炭疽菌中糖基水解酶分布情況
注:ID來自Broad數據庫。下表同。
2.3 希金斯炭疽菌中含有的CBMs
通過對碳水化合物綁定結構蛋白進行分析,發現CBM50數量最多,為18個,其次為CBM13,數量為10個,上述蛋白所占比例為58.33%;此外,還涉及CBM1、CBM2、CBM3、CBM5、CBM12、CBM32、CBM63、CBM67等類別蛋白(表2)。值得關注的是,CH063_04845屬于CBM10/CBM2類別,是CZAymes酶類主要類別中存在的唯一1個CBM蛋白,推測它在發揮CBM功能方面具有特殊性。因此,對于該蛋白有待于開展更為細致的工作,以期解釋它們與其他蛋白之間的差別。
2.4 希金斯炭疽菌中含有的AAs
通過對輔助酶家族進行分析,發現AA9數量較多,為12個,其次為AA7,為6個,上述蛋白所占比例為54.55%;此外,還涉及AA1、AA2、AA3、AA5等類別蛋白(表3)。該類蛋白作為近些年新分類蛋白,具有重要的生物學功能,因此,有待于開展后續生物學研究,從而更好地解析該類蛋白的功能。
2.5 希金斯炭疽菌中含有的CEs
通過對碳水化合物酯酶蛋白進行分析,發現CE5、CE10數量較多,均為9個,其次為CE1,數量為3個,上述蛋白所占比例為70.00%;此外,還涉及CE3、CE4、CE6、CE8、CE10等類別蛋白(表4)。
2.6 希金斯炭疽菌中含有的PLs
通過對多糖裂解酶蛋白進行分析,發現PL1數量較多,為10個,其次為PL3,數量為8個,上述蛋白所占比例為 78.26%;此外,還涉及PL4、PL9等類別蛋白,其數量較少(表5)。

表2 希金斯炭疽菌中碳水化合物綁定結構分布情況

表3 希金斯炭疽菌中輔助酶類家族分布情況

表4 希金斯炭疽菌中碳水化合物酯酶的分布情況
2.7 希金斯炭疽菌中含有的GTs
通過對糖基轉移酶蛋白進行分析,發現僅包含GT2、GT8、GT22、GT34等4個蛋白,其ID分別為CH063_00779、CH063_11721、CH063_11413、CH063_15057(表6)。與其他酶類進行比較,該酶類較少,推測它們在希金斯炭疽菌危害植物過程中發揮著特有功能。
2.8 希金斯炭疽菌中含有的復合類別蛋白
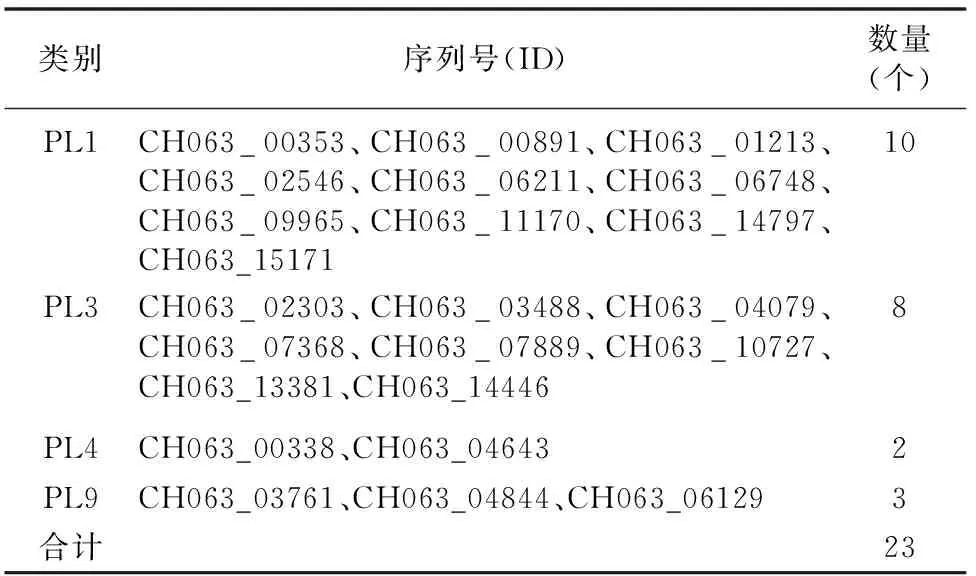
表5 希金斯炭疽菌中多糖裂解酶的分布情況

表6 希金斯炭疽菌中糖基轉移酶的分布情況
通過對希金斯炭疽菌中CAZymes進行分析,發現除含有典型的GHs、CBMs、AAs、CEs、GTs以外,還含有一些復合類別的蛋白,涉及GHs/CBMs、AAs/CBMs、CEs/CBMs等3類,尤以GHs/CBMs類別中的蛋白數量居多,為17個,所占比例高達68.00%(表7)。上述復合類別的蛋白所具有的具體類別呈現出多樣性特點,推測它們在希金斯炭疽菌危害植物過程中發揮著多種功能。

表7 希金斯炭疽菌中復合類別蛋白的分布情況
3 討論與結論
隨著希金斯炭疽菌全基因組序列的釋放[29],學術界對其致病因子開展了大量研究。本研究結果與前人報道[29]不僅在CAZymes總數量方面有所差別,而且在具體的酶類別數量方面也有一定的差別。究其原因,就總數量而言,由于本研究所依賴的蛋白屬于前期通過生物信息學分析所獲得的分泌蛋白;在數量統計方面,以CAZymes主要類別和復合類別分別進行的統計,使得歸屬類別方面存在一定的差異,若將復合類別中的CAZymes分別歸屬到主要類別中,那么總的數量為263個,當然該統計存在25個重復蛋白;此外,就不同類別中CAZymes數量而言,按照上述統計方法,本研究所獲得GHs、GTs、CBMs、CEs、PLs以及AAs則分別含有75、4、48、30、23、33個。可以發現,本研究所獲得GHs數量明顯少于前人所報道的361個[29],原因尚不明確,有待于進一步研究。前人所開展的研究中CAZymes的具體類別有5個[29],而最新的分類類別有6個,造成具體不同類別分項數量方面的差異。
本研究基于前期通過全基因組序列預測所獲得的分泌蛋白,利用現有在線工具所獲得的CAZymes是否為希金斯炭疽菌中的全部,均有待于進一步開展試驗進行驗證。在數量方面,O′Connell等報道為689個[29],該報道的數量遠遠高于本研究結果,就每一類中的數量報道也有不同,原因也有待于進一步明確。
前期明確希金斯炭疽菌中含有658個分泌蛋白,本研究利用CAT在線預測程序,明確該菌中含有238個CAZymes,分為主要類別和復合類別2類,前者包括75個GHs、48個CBMs、33個AAs、30個CEs、23個PLs、4個GTs;后者則包括17個GHs/CBMs、4個AAs/CBMs、4個CEs/CBMs等。本研究為深入開展希金斯炭疽菌中CAZymes的功能研究以及進一步找尋新的藥劑作用靶標打下了一定的理論基礎。
[1]van den Brink J,de Vries R P.Fungal enzyme sets for plant polysaccharide degradation[J]. Applied Microbiology and Biotechnology,2011,91(6):1477-1492.
[2]陳相永,陳捷胤,肖紅利,等. 植物病原真菌寄生性與分泌蛋白組CAZymes的比較分析[J]. 植物病理學報,2014,44(2):163-172.
[3]Lombard V,Ramulu H G,Drula E,et al.The carbohydrate-active enzymes database(CAZy) in 2013[J]. Nucleic Acids Research,2014,42:D490-D495.
[4]Bourne Y,Henrissat B.Glycoside hydrolases and glycosyltransferases: families and functional modules[J]. Current Opinion in Structural Biology,2001,11(5):593-600.
[5]Campbell J A,Davies G J,Bulone V,et al.A classification of nucleotide-diphospho-sugar glycosyltransferases based on amino acid sequence similarities[J]. Biochemical Journal,1998,329(3):939-941.
[6]Lombard V,Bernard T,Rancurel C,et al.A hierarchical classification of polysaccharide lyases for glycogenomics[J]. Biochemical Journal,2010,432(3):437-444.
[7]Levasseur A,Drula E,Lombard V,et al.Expansion of the enzymatic repertoire of the CAZy database to integrate auxiliary redox enzymes[J]. Biotechnology for Biofuels,2013,6(1):1-14.
[8]Boraston A B,Bolam D N,Gilbert H J,et al.Carbohydrate-binding modules: fine-tuning polysaccharide recognition[J]. Biochemical Journal,2004,382(3):769-81.
[9]沈瑞清,張 萍,郭成瑾,等. 寧夏炭疽菌屬真菌資源研究[J]. 河南農業科學,2012,41(5):100-102.
[10]Hyde K D,Cai L,Cannon P F,et al.Colletotrichum-names in current use[J]. Fungal Diversity,2009,39(1):147-182.
[11]盧博彬,楊 暹. 菜心炭疽病研究進展[J]. 長江蔬菜,2009(24):1-5.
[12]況福元,吳小麗,呂風青,等. 菜心炭疽病菌拮抗細菌的篩選及鑒定[J]. 微生物學通報,2009,32(9):1350-1355.
[13]Huser A,Takahara H,Schmalenbach W,et al.Discovery of pathogenicity genes in the crucifer anthracnose fungusColletotrichumhigginsianum,using random insertional mutagenesis[J]. Molecular Plant-Microbe Interactions,2009,22(2):143-156.
[14]Ushimaru T,Terada H,Tsuboi K,et al.Development of an efficient gene targeting system inColletotrichumhigginsianumusing a non-homologous end-joining mutant andAgrobacteriumtumefaciens-mediated gene transfer[J]. Molecular Genetics and Genomics,2010,284(5):357-371.
[15]周 鵬. 希金斯刺盤孢T-DNA插入體庫的構建、篩選及相關突變體基因的克隆[D]. 武漢:華中農業大學,2011.
[16]趙 典. 希金斯刺盤孢T-DNA插入體庫的篩選及相關突變體基因的克隆[D]. 武漢:華中農業大學,2012.
[17]郭巨先,劉玉濤,楊 暹. 鉀營養對菜薹(菜心)炭疽病發生和植株防御酶活性的影響[J]. 中國蔬菜,2012(14):86-89.
[18]林春花,蔡志英,黃貴修. 全基因組法繪制禾谷炭疽菌和希金斯炭疽菌中MAPK級聯信號途徑簡圖[J]. 熱帶作物學報,2012,33(4):674-680.
[19]韓長志. 希金斯炭疽菌RGS蛋白生物信息學分析[J]. 生物技術,2014,49(1):36-41.
[20]韓長志. 希金斯炭疽菌PITP生物信息學分析[J]. 湖北農業科學,2015,54(3):713-716.
[21]韓長志. 希金斯炭疽菌GPCR蛋白生物信息學分析[J]. 華中師范大學學報(自然科學版),2015(2):246-251.
[22]韓長志. 希金斯炭疽菌腺苷酸環化酶生物信息學分析[J]. 湖北農業科學,2014,53(20):4992-4996.
[23]韓長志. 希金斯炭疽菌腺苷酸環化酶相關蛋白生物信息學分析[J]. 生物技術,2014,53(6):4992-4996.
[24]韓長志. 希金斯炭疽菌14-3-3蛋白質生物信息學分析[J]. 湖北農業科學,2014,53(15):3669-3672.
[25]韓長志. 希金斯炭疽菌中5個典型Septin的生物信息學分析[J]. 河南農業科學,2014,43(8):91-96.
[26]韓長志. 希金斯炭疽菌磷酸二酯酶生物信息學分析[J]. 科學技術與工程,2014,14(27):163-167.
[27]韓長志. 全基因組預測希金斯炭疽菌的候選效應分子[J]. 生物技術,2015,25(6):546-551.
[28]Park B H,Karpinets T V,Syed M H,et al.CAZymes Analysis Toolkit (CAT): web service for searching and analyzing carbohydrate-active enzymes in a newly sequenced organism using CAZy database[J]. Glycobiology,2010,20(12):1574-1584.
[29]O’Connell R J,Thon M R,Hacquard S,et al.Lifestyle transitions in plant pathogenicColletotrichumfungi deciphered by genome and transcriptome analyses[J]. Nature Genetics,2012,44(9):1060-1065.
10.15889/j.issn.1002-1302.2017.02.006
2015-12-16
國家自然科學基金(編號:31560211);云南省森林災害預警與控制重點實驗室開放基金(編號:ZK150004);云南省優勢特色重點學科生物學一級學科建設項目(編號:50097505);云南省高校林下生物資源保護及利用科技創新團隊(編號:2014015)。
韓長志(1981—),男,河北石家莊人,博士,副教授,研究方向為經濟林木病害生物防治與真菌分子生物學。E-mail:hanchangzhi2010@163.com。
S432.4+4
A
1002-1302(2017)02-0024-04
韓長志. 全基因組預測希金斯炭疽菌中碳水化合物酶類蛋白[J]. 江蘇農業科學,2017,45(2):24-28.
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
