鏟形杰氏渦蟲(單腸目,達氏科)新紀錄的形態學描述及其分子系統學分析
2017-02-18 07:05:12張呂蔡慧鮑思全周靖汪安泰
四川動物 2017年1期
張呂, 蔡慧, 鮑思全, 周靖, 汪安泰
(深圳大學生命與海洋科學學院,深圳市海洋生物資源與生態環境重點實驗室,廣東深圳518060)
鏟形杰氏渦蟲(單腸目,達氏科)新紀錄的形態學描述及其分子系統學分析
張呂, 蔡慧, 鮑思全, 周靖, 汪安泰*
(深圳大學生命與海洋科學學院,深圳市海洋生物資源與生態環境重點實驗室,廣東深圳518060)
本文對分布于廣東省深圳市僑城濕地(113°58′E,22°31′N)咸淡水的一種單腸目Rhabdocoel渦蟲進行了形態、發育及18S rDNA與28S rDNA分子系統學的分析與比較。結果表明,此渦蟲為達氏科Dalyelliidae鏟形杰氏渦蟲GieysztoriaknipoviciBeklemischev, 1953,為中國新紀錄種。該物種模式標本分布于歐洲黑海近岸。本研究詳細描述了其外部形態與內部結構特征,首次發現該渦蟲精巢超長,前端達咽部,后端至腹側,呈長條形(該屬渦蟲2個精巢呈球形或橢圓形,位于腸后腹側);除了骨質陰莖之外還具有可再生的骨質陰莖鞘。陰莖鞘位于骨質陰莖外側,半包裹骨質陰莖,呈長條形骨質結構。交配后的個體常在交配囊內發現陰莖鞘1~5個。鑒于該物種特殊的形態學特征,其分類地位還有待進一步研究。
被桿體綱;單腸目;鏟形杰氏渦蟲;形態學;分子系統發生;中國
20世紀中期,渦蟲類(turbellarians)在分類上隸屬于扁形動物門Platyhelminthes渦蟲綱Turbellaria。隨著渦蟲類系統進化研究的深入,原“渦蟲綱”內各類群的分類地位出現了很大變化,2005年后停止使用“渦蟲綱”名詞。一些研究認為無腸類屬于后口動物Deuterostomia,在進化上與棘皮類Echinodermata接近,建議設立“無腸綱”Acoelomorpha(Rhode,1986)。近些年研究發現異渦蟲綱Xenoturbella與“無腸綱”屬于系統進化上的姊妹群,提出設立“異無腸動物門”Xenacoelomorpha(Philippeetal.,2011;Cinar,2014;Perea-Atienzaetal.,2015)。1985年后,扁形動物門內剩下的渦蟲類被重新歸類為2個綱:被桿體綱Rhabditophora和鏈蟲綱Catenulida(Nakano,2015)。被桿體綱在淡水、咸淡水和海水中分布廣泛,下屬6個目,其中單腸目Rhabdocoela物種數量最多,已記錄34科194屬1 000余種。杰氏渦蟲屬Gieysztoria是記錄物種最多的屬,已記錄96種(http://turbellaria.umaine.edu/;容粗徨等,2016),隸屬于達氏科Dalyelliidae(汪安泰,2004)。
中國的杰氏渦蟲屬已記錄11種,分別是深圳杰氏渦蟲G.shenzhensis(汪安泰,吳海龍,2005)、麗杰氏渦蟲G.pulchra(汪安泰,鄧利,2006)、大變杰氏渦蟲九刺亞種G.macrovariata9-spinosa(汪安泰,鄧利,2006)、武夷山杰氏渦蟲G.wuyishanensis(賴曉婷等,2013)、雙斑杰氏渦蟲G.bimaculatas(Luetal.,2013)、廣東杰氏渦蟲G.guangdongensis(Luetal.,2013)、石巖杰氏渦蟲G.shiyanensis(Xiaetal.,2014)、七星巖杰氏渦蟲G.qixingyanensis(容粗徨等,2016)、大變杰氏渦蟲G.macrovariata(Zhangetal.,2014)、汕頭杰氏渦蟲G.shantouensis(Zhangetal.,2014)、惠州杰氏渦蟲G.huizhouensis(Zhangetal.,2014)。以上11種杰氏渦蟲均生活在華南地區淡水環境,迄今尚無咸淡水或海棲物種的報道。
2014年10月,在廣東省深圳市的濱海濕地首次開展咸淡水杰氏渦蟲多樣性研究,獲得20只個體。通過活體、整裝片、連續組織切片、骨質陰莖分離以及個體發育的形態學綜合觀測,結合18S rDNA系統進化分析,鑒定為杰氏渦蟲屬中國新紀錄,即鏟形杰氏渦蟲G.knipoviciBeklemischev, 1953。本研究首次發現獨特于杰氏渦蟲屬的數項形態學特征,并對其做了分析與討論。
1 材料與方法
1.1 采集與培養
用200目自制水網撈取水草,于水桶內擺洗,水網過濾,網內物洗入塑料保鮮盒,室內體視顯微鏡下分離。分離的渦蟲置于直徑6 cm玻璃蒸發皿,近窗。每天喂食1次草履蟲,補充失去水分。培養用水為靜置的原生境水,pH6.8~7.0,室溫培養,空調控溫(25 ℃±1 ℃)。
1.2 個體發育觀測
成熟的渦蟲個體每天產下的卵移至專門的培養皿,注入原生境水,記錄產卵日期與卵數,每日監測卵胚發育與孵化情況,統計每枚卵的孵化周期與孵出的幼蟲數,共觀察593枚卵。取剛孵出幼蟲單獨培養,每天補充草履蟲,觀察幼蟲發育過程。對同個蟲體的子一代、子二代老化個體的骨質陰莖進行分離與觀測,重點計數每個個體骨質陰莖末端小刺的數目。
1.3 模式標本制作
清洗蟲體,置于載玻片,吸出水分,5%乙醇麻醉,用蓋玻片輕壓蟲體,Bouin氏固定液(成分)固定1 h,將蟲體移至稱量瓶,新鮮固定液固定12~24 h,高純水和70%乙醇換洗,直至蟲體無黃色,Mayer氏蘇木精浸染2 h(40 ℃),2%HCl分色20~30 min,0.2%氨水還原10 s,常規梯度乙醇脫水,0.5%伊紅復染5 min,梯度乙醇脫水,二甲苯透明,中性樹膠封片。
1.4 組織切片
蟲體經4%多聚甲醛溶液(4 ℃)處死,Bouin氏固定液(25 ℃)固定過夜,水洗后常規石蠟切片,使用Leica RM2235切片機分別進行定向縱切、橫切及水平切,切片厚8 μm,蘇木精-伊紅染色,逐級乙醇脫水,二甲苯透明后,中性樹膠封片。
1.5 骨質陰莖分離
蟲體清洗干凈,置于載玻片,吸出水分,滴加渦蟲分離劑,用1號昆蟲針在體視顯微鏡下分離骨質陰莖,加蓋玻片,置于Olympus BX51微分干涉顯微鏡下觀察,Olympus DP72數碼相機拍照。
1.6 分子系統學
用于系統發育分析的標本在干凈的原生境水中饑餓處理至少1 d。從饑餓處理的標本中隨機挑選3個經液氮處理后用E.Z.N.A.TMMollusc DNA KIT(Omega,Norcross,GA,USA)提取DNA(標記為Gieysztoriaknipovici1~3)。在目標18S rDNA和28S rDNA基因通過PCR進行擴增的過程中,分別需要用到基于18S rDNA和28S rDNA基因序列設計的1對特異性引物(CY18SF:5’-GCGAATGGCTCATTAAATCAG-3’,CY18SR:5’-CTTGTTACGACTTTTACTTCC-3’;28SF-LSU5:5’-TAGGTCGACCCGCTGAAYTTA-3’,28SR-LSUD6-3:5’-GGAACCCTTCTCCACTTCAGTC-3’)。加入Premix TaqTM(TaKaRa,大連,中國)進行PCR,得到的PCR產物連接、克隆到pMDTM19-T Vector Cloning Kit(TaKaRa,大連,中國),然后轉化到制備成感受態的大腸桿菌EscherichiacoliTop 10中。涂板、挑取單克隆和培養后形成的菌液由北京基因組研究所進行測序(華大基因測序,深圳,中國)。利用NCBI BLAST獲得同源序列(表1)。使用MEGA 6.0進行分子系統的進化分析,所有的序列均被修剪至最短序列。利用最大似然法(ML)和鄰接法(NJ)分別使用Tamura-Nei和Kimura 2-parameter模型重建系統發育樹,以推斷這些物種之間的進化關系。

表1 系統進化分析中選擇的核苷酸序列
1.7 數據收集與處理
整裝片與切片標本在Olympus BX51微分干涉顯微鏡下觀察,Olympus DP72數碼相機拍照,DP 2-BSW測量。應用Photoshop 7.0編輯數碼圖片,參照數碼圖片繪制模式圖,Excel處理數據。
1.8 縮寫詞
ag.黏液腺體adhesive glands, b. 腦brain, bc. 交配囊bursa copulatrix, e. 眼eye, ed. 射精管ejaculatory duct, eg.卵egg, en. 消化道enteron, go. 生殖孔gonopore, m. 口mouth, o. 卵巢ovary, ov. 輸卵管oviduct, p. 咽pharynx, pe. 陰莖penis, pg. 咽腺pharyngeal glands, pvg. 顆粒囊腺prostate vesicle glands, ps. 陰莖鞘penis sheath, pv. 顆粒囊prostate vesicle, rs. 受精囊seminal receptacle, t. 精巢testis, u. 子宮uterus, v. 卵黃腺vitellaria, vd. 輸精管vas deferens, vs. 儲精囊vesicula seminalis。
2 結果
2.1 鏟形杰氏渦蟲GieysztoriaknipoviciBeklemischev, 1953,中國新紀錄(圖1~圖3)
中國標本:編號JX201409-1~JX201409-7,2014年采自廣東省深圳市華僑濕地(113°58′E,22°31′N)的水生植物上。所有模式標本保存于中國科學院動物標本館。
2.2 外部特征
成熟個體體長1 250 μm,體中部寬249 μm。頭部呈半圓形,中部呈圓柱狀,尾部呈圓錐形,具花簇狀突起,具達氏科典型的體型特征。表皮密布纖毛,頭部和尾部均具觸毛。蟲體背部分布有不規則的棕紅色縱向細紋。眼1對,位于體前端,眼點由許多黑色素球形成,呈腎形。眼間距62 μm。腦神經節分布在眼下方。眼后有酒桶狀的咽,230 μm×180 μm,占體長的1/5。咽后連接腸道,無肛門。咽與腸的交接處具有大而明顯的咽腺(圖2:A~C)。
2.3 生殖系統
雌雄同體。雌性生殖器官由卵巢、輸卵管、受精囊、卵黃腺、子宮、生殖腔、生殖孔和交配囊組成。卵巢1個,位于近尾部的體背側,有1根短的輸卵管,連接受精囊和子宮;受精囊是1個充滿精子的橢圓形。卵黃腺2條,呈棒狀,位于身體背部兩側,左右卵黃腺向后延伸,愈合為卵黃總管,通往子宮。子宮2~4個,具1~3個備用子宮和1個成熟子宮,成熟子宮常含1個橢圓形的卵。子宮的另一端與生殖腔相連。生殖孔1個,位于生殖腔的腹側,開口于體外。交配囊1個,由肌肉環肌構成,呈管狀,囊部長33 μm,寬44 μm,柄部長85 μm,寬16 μm,開口于生殖腔。
雄性生殖器官由精巢、輸精管、儲精囊、顆粒囊、射精管、陰莖刺和前列腺組織組成。精巢1對,位于卵黃腺腹側,精巢前端達咽部,后端延伸至消化道后端的腹側,呈長條形,各有1條輸精管通往儲精囊。輸精管呈細管狀。儲精囊1個,呈近球形,內充滿精子。儲精囊后緊接顆粒囊,其外側分布著顆粒囊腺,顆粒囊后連接射精管。射精管后連接骨質化的陰莖刺,被陰莖鞘包裹。陰莖鞘位于陰莖刺的外側,半包裹陰莖刺,為不規則條狀的骨質結構,位于消化道后端的腹側。陰莖刺由環帶和24~32根小刺組成,全長90~100 μm,呈環抱狀。環帶高55 μm,直徑57 μm,由膠原纖維骨質后形成。每根小刺基部有“U”形的肌肉附著層,分兩側和底部2部分,附著層長度有明顯差異;最外側長10 μm,中部長26 μm。兩側的小刺排列呈羽狀,底部的呈尖刀狀。小刺長度有明顯差異,底部的由兩側向中間依次增長,最外側小刺長11 μm,中部小刺長21 μm(圖1,圖2:D~J,圖3)。
A. 整體形態; B. 生殖系統; C. 自然狀態的骨質陰莖; D. 陰莖鞘; E. 壓片展開的骨質陰莖。
A. whole body; B. copulatory apparatus; C. penis stylet, without extension; D. penis stylet sheath; E. penis stylet, with extension.
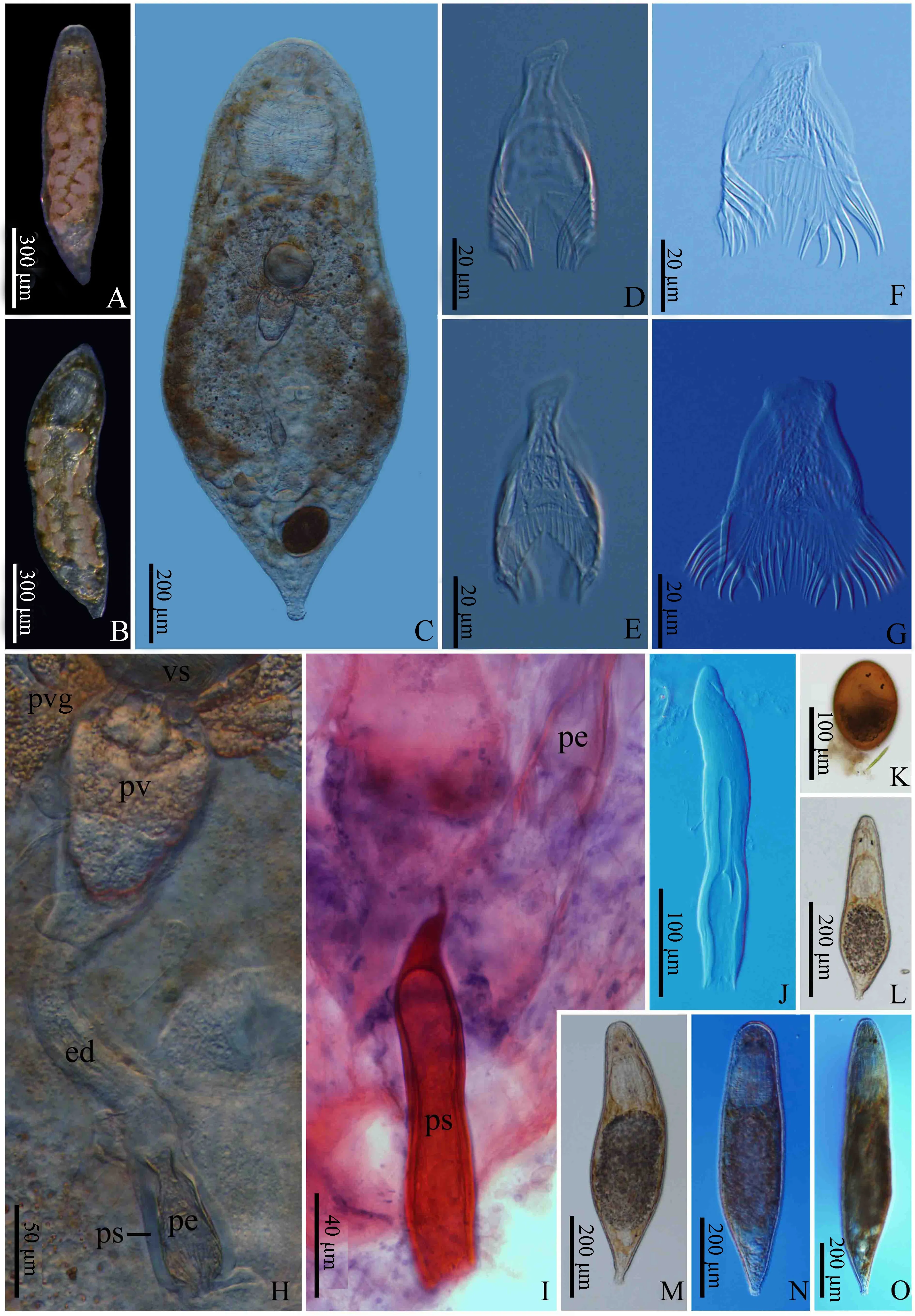
圖2 鏟形杰氏渦蟲
Fig. 2GieysztoriaknipoviciBeklemischev, 1953
A. 活體背面觀; B~C. 活體腹面觀; D~E. 自然形態的骨質陰莖; F. 半展開的骨質陰莖; G. 完全展開的骨質陰莖; H. 壓片狀態下的雄性生殖器官(示儲精囊、顆粒囊、射精管、骨質陰莖、陰莖鞘); I. 整裝片中被伊紅染成紅色的陰莖鞘; J. 單獨剝離分出的陰莖鞘; K~O. 個體發育(依次為產卵后第3天形態、孵化后第1天形態、孵化后第3天形態、孵化后第4天形態、孵化后第7天形態)。
A. free-living individual, dorsal view; B-C. free-living individual, ventral view; D-E. penis stylet, without extension; F. penis stylet, half-extension; G. penis stylet, complete extension; H. male reproductive organ of the squeezed living animal (showing vesicula seminalis, prostate vesicle, ejaculatory duct, penis stylet, penis stylet sheath); I. penis stylet sheath from the whole-mounted specimen, stained by hematoxylin-eosin staining method; J. penis stylet sheath; K-O. individual development (the 3rd day in the spawning period, the 1st day, 3rd day, 4th day, and 7th day in the incubation period, respectively).
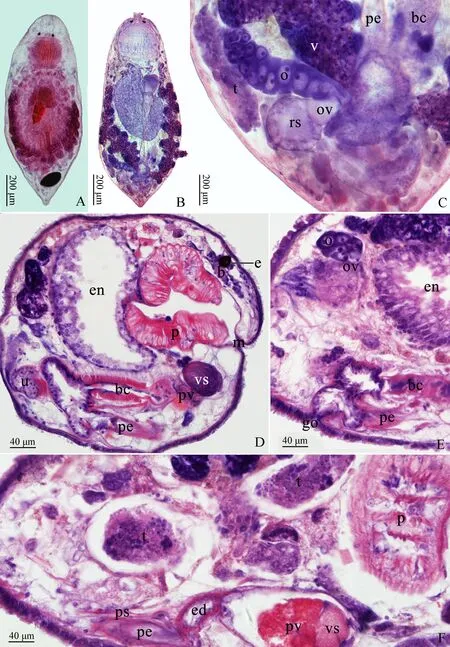
圖3 鏟形杰氏渦蟲整裝片及切片
A~B. 整裝片, 中國標本; C. 整裝片示蟲體尾部的生殖器官; D. 整體縱向切片; E. 局部縱向切片示生殖孔; F. 縱向切片示一側雙精巢結構。
A-B. whole-mounted specimen, Chinese type; C. caudal portion of the whole-mounted specimen (showing copulatory apparatus); D. longitudinal whole body section; E. longitudinal section (showing gonopore); F. longitudinal section (showing testes).
2.4 生殖與發育
渦蟲從產卵到幼體孵出的時間(卵孵化期)平均為2.76 d±0.69 d(1~9 d,n=403),幼體孵出至性成熟并產下第一枚卵平均需要7.14 d±0.90 d(6~8 d,n=7),平均產卵周期18.63 d±9.50 d(10~38 d,n=7),個體平均產卵數84.71枚±28.53枚(74~134枚,n=7)。生活周期(幼體孵出至死亡)平均為29.57 d±15.78 d(16~65 d,n=7)。
渦蟲單系培養觀測發現,繁殖的子一代、子二代的陰莖刺小刺的數目仍有變化(小刺24~32根)。
2.5 生境與習性
本文渦蟲生活于咸淡水環境,屬于深圳灣填海造地預留的城市湖泊濕地,占地面積約6.85×105m2,水溫15~22 ℃,日照11~13 h,海水鹽度0‰~8‰,水質較渾濁,水源來自深圳灣漲潮的海水。渦蟲在自然環境中分布于水生植物根部,在實驗室易于飼養和繁殖(15~25 ℃),以草履蟲為食,用原生境水飼養1年,未見異常。
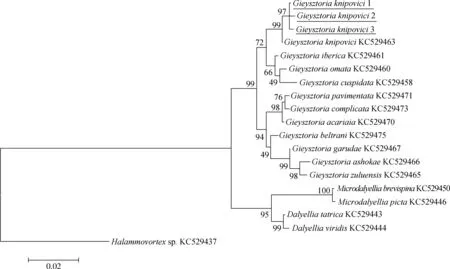
圖4 以最大似然法構建的18S rDNA的進化樹(bootstrap=1 000, Tamura-Nei模型)
圖中劃線部分即為本文研究物種; 下同。
The species studied in this paper is underlined; the same below.
2.6 分子系統學分析
利用MEGA 6.0將測序所得的鏟形杰氏渦蟲的18S rDNA序列與11種杰氏渦蟲屬物種、5種達氏科渦蟲,以及其他類群渦蟲的18S rDNA序列(圖4,圖5),其中渦蟲(Halammovortexsp.,KC529437)為外群,進行基于統計學的最大似然法和基于距離的鄰接法構建系統樹進行系統發育關系分析(圖4,圖5)。
同樣的,將測序所得的鏟形杰氏渦蟲的28S rDNA序列與上述16種渦蟲的28S rDNA序列構建系統樹,進行系統發育關系分析(圖6,圖7)。
ML法構建的進化樹和NJ法構建的進化樹結構一致。本研究的內群分為3支,分別為達氏渦蟲屬Dalyellia支系、小達氏渦蟲屬Microdalyellia支系和杰氏渦蟲屬支系。3只不同個體的鏟形杰氏渦蟲先匯聚成一支,可認為是同種渦蟲。鏟形杰氏渦蟲匯入杰氏渦蟲屬支系中。
3 討論
本研究首次在中國咸淡水水域中發現杰氏渦蟲屬物種,即鏟形杰氏渦蟲。杰氏渦蟲屬物種通常生活于淡水環境,體呈梭形,長1 mm左右,眼點2個,位于咽的前方,咽后有1個橢圓形的腸,口位于腹面。2條卵黃腺分布在體背兩側,后端在腸的后緣會合入輸卵管。腸后緣背部有1個卵巢、消化道后端腹部有2個精巢,儲精囊、顆粒囊和骨質陰莖依次相連。骨質陰莖基部的形狀以及角質刺的數量、形狀、大小是鑒別杰氏渦蟲的關鍵特征(汪安泰,吳海龍,2005;http://turbellaria.umaine.edu/;孫婷等,2016)。小達氏渦蟲屬的渦蟲具有1對精巢,位于卵黃腺前端的腹側(容粗徨等,2016)。鏟形杰氏渦蟲模式標本產地位于里海的伊朗、土庫曼斯坦、阿塞拜疆的海港。該物種早期歸于小達氏渦蟲屬(Beklemischev,1953),后歸于杰氏渦蟲屬(Luther,1955)。模式標本的骨質陰莖由1個柄部與24~32根小刺組成,小刺呈環形分布,其他形態學數據不詳。本研究嚴格單系繁殖的子一代與子二代的骨質陰莖上的小刺數目為24~32根,證明該物種陰莖上的小刺數目有固定的變化范圍。
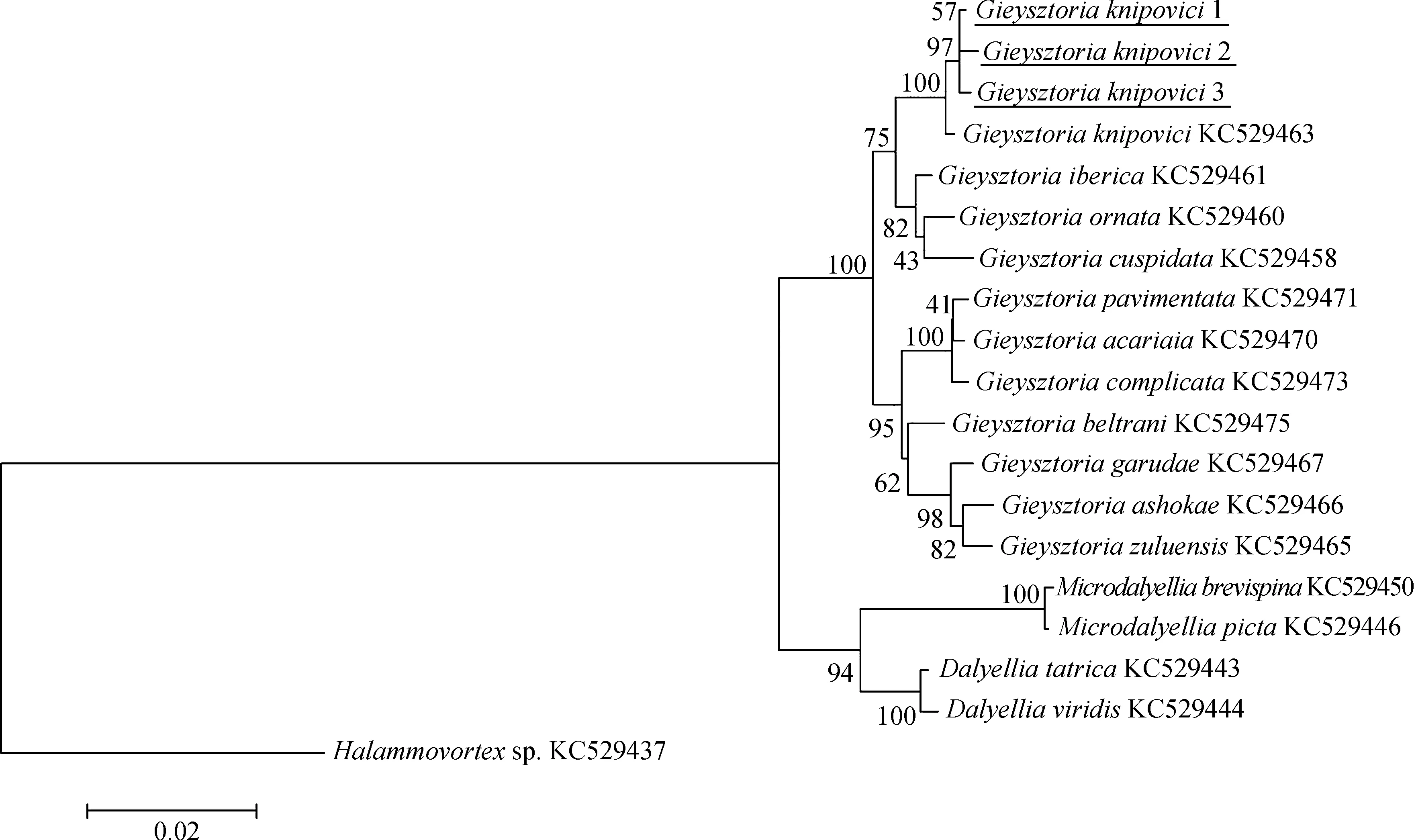
圖5 以鄰接法構建的18S rDNA的進化樹(bootstrap=1 000, Kimura 2-parameter模型)
Fig. 5 Phylogenetic tree of 18S rDNA generated with the neighbor-joining method (bootstrap=1 000, Kimura 2-parameter model)
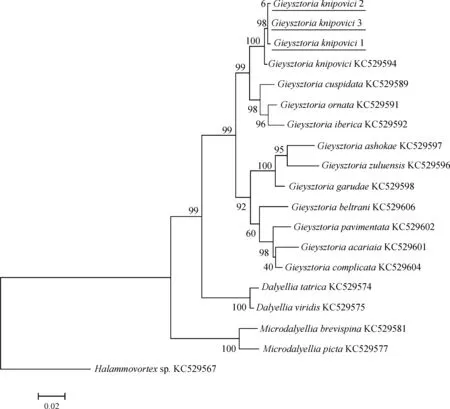
圖6 以最大似然法構建的28S rDNA的進化樹(bootstrap=1 000, Tamura-Nei模型)
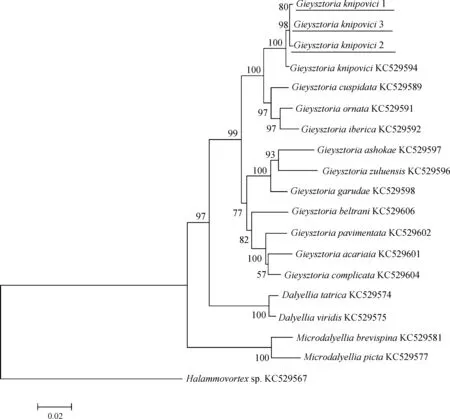
圖7 以鄰接法構建的28S rDNA的進化樹(bootstrap=1 000,Kimura 2-parameter模型)
本研究首次發現,本文標本與杰氏渦蟲屬具有明顯不同的形態學結構:(1)1對精巢向前延伸到卵黃腺前端的腹側,后端延伸到消化道的后端;(2)骨質結構除了骨質陰莖外還存在另一個可再生的骨質陰莖鞘,位于骨質陰莖的外側,呈長條狀,中央內凹,上端呈封閉半橢圓形或鷹爪狀,下端呈鋸齒狀的半圓形,側面呈開放竹筒狀,半包裹骨質陰莖;(3)在成蟲受精囊內常有1~3個交配后遺留的骨質陰莖鞘;(4)蟲體的骨質陰莖微小,射精管長,陰莖鞘有助于受精過程,是雄性生殖器官的重要附件;(5)陰莖鞘在交配丟失后,會重新再生一個;(6)受精囊長度是骨質陰莖長度的2倍以上;(7)骨質陰莖鞘長度是陰莖長度的2~4倍;(8)在輸卵管內與受精囊內均發現有骨質陰莖鞘。
上述中國標本獨特的雄性生殖器官結構與交配特性,與杰氏渦蟲屬的特征差異顯著。另一方面,本研究18S rDNA序列和28S rDNA序列與歐洲標本(Vanetal.,2013)的親緣關系顯示,本文物種屬于鏟形杰氏渦蟲,且屬于杰氏渦蟲屬。即本文分子系統學數據不支持形態學數據,其最后的分類地位有待進一步研究。
致謝:2013級方楚玉與趙佳琪同學,2011級陳鈺輝同學協助修改本文,在此一并致謝!
賴曉婷, 盧彥宏, 汪安泰. 2013. 杰氏渦蟲屬一新種(扁形動物門,單腸目, 達氏科)[J]. 動物分類學報, 38(2): 251-256.
容粗徨, 劉旭坤, 趙佳琪, 等. 2016. 達氏科(扁形動物門, 單腸目)渦蟲二新種[J]. 四川動物, 35(1): 97-103.
孫婷, 何一, 方楚玉, 等. 2016. 華南常見渦蟲類群的的分布與鑒別[J]. 生物學通報, 51(3): 47-51.
汪安泰, 鄧利. 2006. 杰氏渦蟲屬一新種及中國一新紀錄種(扁形動物門, 單腸目, 達氏科)[J]. 動物分類學報, 31(1): 120-124.
汪安泰, 吳海龍. 2005. 達氏科中國一新紀錄屬一新種(扁形動物門, 單腸目, 達氏亞目)[J]. 動物分類學報, 30(3): 516-519.
汪安泰. 2004. 中國渦蟲一新紀錄科達氏渦蟲屬一新種(單腸目,達氏科)[J]. 動物分類學報, 29(4): 697-699.
Beklemischev VN. 1953. Ciliated worms (Turbellaria) of the Caspian Sea. Ⅰ. Rhabdocoela (with some remarks on Rhabdocoea of Aral)[J]. Byull Mosk Ovskobs Isp Prirody, 58: 35-45.
Cinar ME. 2014. Checklist of the phyla Platyhelminthes, Xenacoelomorpha, Nematoda, Acanthocephala, Myxozoa, Tardigrada, Cephalorhyncha, Nemertea, Echiura, Brachiopoda, Phoronida, Chaetognatha, and Chordata (Tunicata, Cephalochordata, and Hemichordata) from the coasts of Turkey[J]. Turkish Journal of Zoology, 38(6): 698-722.
Lu YH, Wu CC, Xia XJ,etal. 2013. Two new species ofGieysztoria(Platyhelminthes, Rhabdocoela, Dalyelliidae) from a freshwater artificial lake in Shenzhen, China[J]. Zootaxa, 3745(5): 569-578.
Luther A. 1955. Die Dalyelliiden (Turbellaria, Neorhabdocoela) Eine Monographie [J]. Acta Zoologica Fennica, 87: 1-337.
Nakano H. 2015. What is Xenoturbella?[J]. Zoological Letters, 1: 22.
Perea-Atienza E, Gavilan B, Chiodin M,etal. 2015. The nervous system of Xenacoelomorpha: a genomic perspective[J]. Journal of Experimental Biology, 218(4): 618-628.
Philippe H, Brinkmann H, Copley RR,etal. 2011. Acoelomorph flatworms are deuterostomes related to Xenoturbella[J]. Nature, 470(7333): 255-258.
Rhode K. 1986. Ultrastructural studies ofAustramphilinaelongata(Cestoda, Amphilinidea)[J]. Zoomorphology, 106(2): 91-102.
Van Steenkiste N, Tessens B, Willems W,etal. 2013. A comprehensive molecular phylogeny of Dalytyphloplanida (Platyhelminthes: Rhabdocoela) reveals multiple escapes from the marine environment and origins of symbiotic relationships[J]. PLoS ONE, 8(3): 1-13.
Xia XJ, Lu YH, Wang AT. 2014. A new species ofAequales, genusGieysztoria(Rhabdocoela: Dalyelliidae)from China[J]. Zoological Systematics, 39(2): 229-235.
Zhang H, Li YK, Wu CC,etal. 2014. Two new species of the genusGieysztoria(Rhabdocoela: Dalyelliidae) from China[J]. Zoological Systematics, 39(4): 485-495.
Gieysztoriaknipovici, A New Record of Turbellarian Species (Rhabdocoela, Dalyelliidae) from China with Morphological Description and Molecular Phylogenetic Analysis
ZHANG Lü, CAI Hui, BAO Siquan, ZHOU Jing, WANG Antai*
(College of Life Sciences and Oceanography, Shenzhen Key Laboratory Bioresources and Ecology, Shenzhen, Guangdong Province 518060, China)
A species of the order Rhabdocoela (Dalyelliidae,Gieysztoria) was collected from a brackish-water wetland (113°58′E, 22°31′N) of the Overseas Chinese Town in Shenzhen city, Guangdong province. The morphology, individual development as well as 18S rDNA and 28S rDNA phylogenesis of the species were analyzed. The results suggested that the present species isGieysztoriaknipoviciBeklemischev, 1953, belonging to the family Dalyelliidae, as a newly recorded species in China. The type specimen was collected from the coast of Black Sea with the only description of its stylet features by Beklemischev (1953). For the first time, the following features were reported in this study: 1) extremely long testes were located in the ventral side of body, between the anterior vitellarium and the posterior intestine; 2) apart from the sclerotic stylet, a reproducible sclerotic structure and elongated penis sheath were found to be located in the outer edge of the stylet, covering half of the stylet; 3) 1 to 5 penis sheaths were found in bursa copulatrix of the individual after mating. Given the special morphological characters ofG.knipovici, its classification warrants further studies.
Rhabditophora; Rhabdocoela;Gieysztoriaknipovici; morphology; molecular phylogeny; China
2016-10-15 接受日期:2016-11-18
廣東省大學生創新項目創新創業訓練計劃項目(201610590041); 深圳市科技計劃項目(JSGG20130411160539208); 深圳市生物產業發展專項基金項目(NYSW20140327010012)
張呂, 男, 本科生, 研究方向:動物學, E-mail:569692501@qq.com
*通信作者Corresponding author, E-mail:wang118@szu.edu.cn
10.11984/j.issn.1000-7083.20160280
Q959.151
A
1000-7083(2017)01-0065-10
