牛磺酸及相關(guān)氨基酸對(duì)大菱鲆幼魚生長性能及TauT mRNA表達(dá)的影響
2017-02-15 08:23:52茜梁萌青鄭珂珂王成強(qiáng)徐后國
水生生物學(xué)報(bào) 2017年1期
關(guān)鍵詞:實(shí)驗(yàn)
柳 茜梁萌青鄭珂珂王成強(qiáng)徐后國
(1. 中國水產(chǎn)科學(xué)研究院黃海水產(chǎn)研究所, 青島 266071; 2. 上海海洋大學(xué)水產(chǎn)與生命學(xué)院, 上海 201306)
牛磺酸及相關(guān)氨基酸對(duì)大菱鲆幼魚生長性能及TauT mRNA表達(dá)的影響
柳 茜1,2梁萌青1鄭珂珂1王成強(qiáng)1,2徐后國1
(1. 中國水產(chǎn)科學(xué)研究院黃海水產(chǎn)研究所, 青島 266071; 2. 上海海洋大學(xué)水產(chǎn)與生命學(xué)院, 上海 201306)
以初始體重為(7.90±0.07) g的大菱鲆為實(shí)驗(yàn)對(duì)象, 魚粉、豆粕、玉米蛋白粉和谷朊粉為主要蛋白質(zhì)來源, 魚油為主要脂肪源, 在此基礎(chǔ)配方中分別添加0、1%、2%牛磺酸, 0.5%蛋氨酸及0.5%半胱氨酸(分別命名為T-0、T-1、T-2、M-0.5和C-0.5), 配制5種等氮等脂的配合飼料, 在室內(nèi)流水養(yǎng)殖系統(tǒng)進(jìn)行為期10周的養(yǎng)殖實(shí)驗(yàn), 目的是研究飼料中含有高比例植物蛋白時(shí)牛磺酸、蛋氨酸和半胱氨酸對(duì)大菱鲆幼魚生長及牛磺酸轉(zhuǎn)運(yùn)載體(TauT) mRNA表達(dá)的影響。結(jié)果表明, 與對(duì)照組相比, T-1、T-2組大菱鲆幼魚的特定生長率(SGR)和飼料效率(FE)提高(P<0.05), 內(nèi)臟指數(shù)(VSI)降低(P<0.05); M-0.5組大菱鲆幼魚SGR和FE較對(duì)照組提高(P>0.05), VSI低于對(duì)照組(P<0.05), 肥滿度(CF)高于對(duì)照組(P<0.05); C-0.5組SGR較對(duì)照組降低(P<0.05), 但FE、VSI和CF與對(duì)照組差異不顯著(P>0.05); T-1、T-2組大菱鲆幼魚肝臟、腦和眼中TauT mRNA相對(duì)表達(dá)量低于對(duì)照組(P<0.05), 且隨著飼料中牛磺酸含量的增加大菱鲆幼魚肝臟、腦和眼中TauT mRNA相對(duì)表達(dá)量降低(P<0.05); M-0.5組大菱鲆幼魚肝臟、腦和眼中TauT mRNA相對(duì)表達(dá)量高于T-0、T-1、T-2組(P<0.05); C-0.5組大菱鲆幼魚肝臟、腦和眼中TauT mRNA相對(duì)表達(dá)量高于T-0、T-1、T-2組(P<0.05), 但與M-0.5組相比差異不顯著(P>0.05)。綜合分析表明, 在實(shí)驗(yàn)條件下, 飼料中牛磺酸含量為0.48%、1.06%時(shí)能夠提高大菱鲆幼魚的生長性能; 大菱鲆幼魚體內(nèi)TauT mRNA表達(dá)可能受飼料中牛磺酸、蛋氨酸和半胱氨酸等含硫氨基酸的影響。
牛磺酸; 蛋氨酸; 半胱氨酸; 大菱鲆; 牛磺酸轉(zhuǎn)運(yùn)載體(TauT)
牛磺酸(Taurine), 化學(xué)名稱為2-氨基乙磺酸, 俗稱牛膽堿, 是一種小分子的β-含硫氨基酸[1]。研究表明, 牛磺酸是機(jī)體的一種條件性必需氨基酸, 餌料中植物蛋白高時(shí)補(bǔ)充牛磺酸可促進(jìn)生長、提高飼料轉(zhuǎn)化率, 并有效改善綠肝綜合癥[2—6]。已有研究表明, 大菱鲆幼體的生長與其體內(nèi)的牛磺酸水平呈正相關(guān)[7]。蛋氨酸(Methionine)是魚類的一種必需氨基酸, 在動(dòng)物體內(nèi)蛋氨酸以S-腺苷甲硫氨酸的形式作為甲基的主要供體[8,9], 用來合成核酸、角質(zhì)素和膽堿等一些重要化合物[10]; 半胱氨酸(Cysteine)是一種重要的含硫氨基酸, 是組成蛋白質(zhì)的20種天然氨基酸中惟一具有還原性基團(tuán)—巰基的氨基酸[11]; 在哺乳動(dòng)物體內(nèi), 牛磺酸可由蛋氨酸(Methionine)和半胱氨酸(Cysteine)合成。作為動(dòng)物組織中含量最豐富的游離氨基酸, 牛磺酸跨膜轉(zhuǎn)運(yùn)是通過細(xì)胞膜上的牛磺酸轉(zhuǎn)運(yùn)載體(TauT)來完成的[12]。TauT屬于Na+、Cl-依賴型轉(zhuǎn)運(yùn)體, 在鱸(Lateolabrax japonicus)的研究中發(fā)現(xiàn), TauT在心、肝、眼、腦等組織中均有表達(dá)[13]。牛磺酸轉(zhuǎn)運(yùn)載體(TauT)通過跨膜轉(zhuǎn)運(yùn)將胞外牛磺酸轉(zhuǎn)運(yùn)至胞內(nèi), 保證其正常功能; TauT表達(dá)可隨細(xì)胞滲透壓的改變而發(fā)生變化, 其活性也可因不同的調(diào)節(jié)劑而改變。目前國內(nèi)外有關(guān)魚類TauT的研究大多集中在其滲透壓調(diào)節(jié)以及外源性牛磺酸對(duì)TauT表達(dá)的影響方面,而飼料中添加牛磺酸及相關(guān)含硫氨基酸對(duì)魚體TauT mRNA表達(dá)的影響未見報(bào)道。本研究以大菱鲆為對(duì)象, 研究飼料中牛磺酸及相關(guān)含硫氨基酸對(duì)大菱鲆幼魚生長及肝臟、腦、眼組織中TauT mRNA相對(duì)表達(dá)量的影響, 為探討牛磺酸在大菱鲆體內(nèi)代謝機(jī)制積累數(shù)據(jù)。
1 材料與方法
1.1 實(shí)驗(yàn)用魚
大菱鲆幼魚購自青島市通用水產(chǎn)養(yǎng)殖有限公司(中國, 青島), 養(yǎng)殖實(shí)驗(yàn)在煙臺(tái)天源水產(chǎn)有限公司(中國, 煙臺(tái))進(jìn)行。實(shí)驗(yàn)開始前, 停食24h, 隨機(jī)挑選大小均勻、體格健壯且體表無病, 平均體重(7.90± 0.07) g的大菱鲆幼魚450尾, 隨機(jī)平均分配到5×3個(gè)養(yǎng)殖桶(40 cm×50 cm×100 cm)中。
1.2 實(shí)驗(yàn)飼料
基礎(chǔ)飼料以魚粉、豆粕、玉米蛋白粉和谷朊粉為主要蛋白質(zhì)來源, 以魚油為主要脂肪來源。對(duì)照組不添加牛磺酸 (T-0), 試驗(yàn)飼料中分別添加1% (T-1)、2% (T-2)牛磺酸、0.5% (M-0.5)蛋氨酸、0.5% (C-0.5)半胱氨酸。5種飼料等氮等脂, 具體配方及營養(yǎng)成分見表 1。飼料原料粉碎后, 過80目篩,按照配方充分混合, 制成粒徑為3 mm的飼料, 然后放入60℃烘箱內(nèi)烘12h, 在-20℃下保存?zhèn)溆谩?shí)驗(yàn)飼料的氨基酸組成見表 2。
1.3 飼養(yǎng)管理
養(yǎng)殖實(shí)驗(yàn)采用流水系統(tǒng), 在200 L的方形塑料養(yǎng)殖桶中進(jìn)行; 水溫(16±1)℃, pH 7.6—8.2, 溶氧含量保持在6 mg/L以上。每天飽食投喂兩次(6:30和16:30), 投喂后0.5h換水以排除殘餌, 實(shí)驗(yàn)從2014.7.19開始至2014.9.26結(jié)束, 養(yǎng)殖周期為10周。
1.4 樣品采集與分析
樣品采集在實(shí)驗(yàn)魚分組前, 隨機(jī)選取10尾大菱鲆作為初始樣品進(jìn)行分析。在實(shí)驗(yàn)結(jié)束后, 饑餓24h, 稱每桶魚總重; 然后每桶取3尾魚分別測(cè)量其體質(zhì)量、體長、內(nèi)臟團(tuán)質(zhì)量以及肝臟質(zhì)量, 用于計(jì)算其形體指標(biāo); 無菌條件下取眼、腦、肝臟樣品于RNAwait保存液(Solarbio)中, -20℃保存, 用于TauT mRNA表達(dá)量分析; 每桶隨機(jī)撈取3尾魚, -20℃保存, 用于常規(guī)分析。
常規(guī)及生化分析飼料和全魚樣品在105℃烘干至恒重, 采用凱式定氮法(VELP, UDK142 automatic distillation unit, 意大利)測(cè)定粗蛋白質(zhì)含量; 粗脂肪含量采用索氏抽提法測(cè)定(FOSS脂肪測(cè)定儀SOXTEC 2050, 瑞典), 樣品在馬福爐中灼燒5h (550℃), 失重法測(cè)定灰分含量。使用日立L-8900型氨基酸分析儀(Titachi L-8900 automatic amino acidanalyzer, Hitachi, 日本)測(cè)定飼料中牛磺酸及氨基酸組成。

表 1 實(shí)驗(yàn)飼料組成及營養(yǎng)成分分析Tab. 1 Formulation and proximate compositions of experimental diets (%)
1.5 牛磺酸轉(zhuǎn)運(yùn)載體(TauT) mRNA表達(dá)量分析
RNA的提取和cDNA的合成 采用RNAiso Plus (TaKaRa)提取組織總RNA, 利用微量核酸測(cè)定儀(Nanodrop ND2000, 美國)檢測(cè)RNA的濃度和純度, 使用1%的瓊脂糖凝膠電泳檢測(cè)RNA的質(zhì)量; 選擇A260/A280在1.8—2.1、凝膠檢測(cè)未降解的RNA, -80℃保存?zhèn)溆谩R蕴崛〉目俁NA為模板, Transcriptor First Strand cDNA Synthesis Kit反轉(zhuǎn)錄試劑盒(Roche, 德國)兩步法, 采用20 μL反應(yīng)體系合成第一鏈cDNA; 得到的cDNA于-80℃保存?zhèn)溆谩?/p>
引物設(shè)計(jì)本實(shí)驗(yàn)采用相對(duì)熒光定量2-??Ct法, 選用在本實(shí)驗(yàn)條件下表達(dá)穩(wěn)定的β-Actin為管家基因(序列號(hào): AY008305), 目的基因?yàn)榇罅怫襎auT (衛(wèi)育良, 未發(fā)表), 引物序列見表 3。
實(shí)時(shí)熒光定量采用SYBR Green Ⅰ染料法, 熒光試劑選用FastStart Essential DNA Green Master (Roche, 德國), 在熒光定量?jī)x(Eppendorf實(shí)時(shí)熒光定量PCR儀, 德國)上進(jìn)行擴(kuò)增和數(shù)據(jù)分析。反應(yīng)總體積20 μL: Master mix 10 μL, 上游引物1 μL,下游引物1 μL, PCR水3 μL, 10倍稀釋的cDNA 5 μL。反應(yīng)條件為: 95.0℃, 10min; 95.0℃, 10s; 60℃, 15s; 72℃, 20s; 進(jìn)行40個(gè)循環(huán); 溶解曲線按儀器默認(rèn)程序進(jìn)行。采用2-??Ct法計(jì)算TauT基因的相對(duì)表達(dá)量,公式中的

表 2 實(shí)驗(yàn)飼料的氨基酸組成(%干物質(zhì))Tab. 2 The amino acid composition of experimental diets (% in dry matter)

表 3 RT-PCR實(shí)驗(yàn)用引物Tab. 3 Real-time PCR primers used in the experiment
1.6 計(jì)算公式及統(tǒng)計(jì)分析
特定生長率(Specific growth rate, SGR, %/d)=100× [ln(終末體重)-ln(初始體重)]/實(shí)驗(yàn)天數(shù)
成活率(Survival rate, SR, %)=100×實(shí)驗(yàn)結(jié)束每桶魚尾數(shù)/實(shí)驗(yàn)開始每桶魚尾數(shù)
攝食率(Feed intake, FI, %/d)=100×每尾魚攝食的飼料干物質(zhì)重/[(實(shí)驗(yàn)結(jié)束時(shí)魚體重+實(shí)驗(yàn)開始時(shí)魚體重)/2×養(yǎng)殖實(shí)驗(yàn)天數(shù)]
飼料效率(Feed efficiency, FE)=(實(shí)驗(yàn)結(jié)束時(shí)魚體重-實(shí)驗(yàn)開始時(shí)魚體重)/每尾魚攝食的飼料干物質(zhì)重
蛋白效率比(Protein efficiency ratio, PER) =(終末體重-初始體重)/蛋白攝入量
蛋白質(zhì)沉積率(Protein productive value, PPV, %)=100×魚體蛋白質(zhì)貯存量/蛋白攝入量
肥滿度(Condition factor, CF, g/cm3)=100×實(shí)驗(yàn)終體重(g)/(魚體長度)3(cm3)
肝指數(shù)(Hepatosomatic index, HSI, %)=100×肝臟重(g)/魚體重(g)
內(nèi)臟指數(shù)(Viscerosomatic index, VSI, %)=100×內(nèi)臟團(tuán)重(g)/魚體重(g)
所有實(shí)驗(yàn)數(shù)據(jù)均用平均值±標(biāo)準(zhǔn)誤表示, 數(shù)據(jù)采用SPSS 17.0進(jìn)行單因素方差分析(One-way ANOVA), 差異顯著(P<0.05)后, 進(jìn)行Tukey多重比較(Tukey's multiple comparison tests)。
2 結(jié)果
2.1 牛磺酸及相關(guān)氨基酸對(duì)大菱鲆幼魚生長性能的影響
牛磺酸及相關(guān)氨基酸對(duì)大菱鲆幼魚生長性能的影響見表 4, 各實(shí)驗(yàn)組成活率無顯著差異(P>0.05);與對(duì)照組相比, 飼料中牛磺酸含量為0.48%和1.06%時(shí)可提高大菱鲆幼魚的終末體重與特定生長率(SGR) (P<0.05), 且隨著飼料中牛磺酸含量的增加, 大菱鲆幼魚的終末體重與SGR提高(P<0.05); 蛋氨酸添加組大菱鲆幼魚終末體重與SGR略高于對(duì)照組, 但差異不顯著(P>0.05); 半胱氨酸添加組大菱鲆幼魚終末體重略低于對(duì)照組, 但差異不顯著(P>0.05), 而SGR則低于對(duì)照組(P<0.05); 大菱鲆幼魚的攝食率(FI)隨著飼料中牛磺酸含量的升高而升高, 且1.06%牛磺酸組FI高于對(duì)照組(P<0.05), 而與對(duì)照組相比,添加蛋氨酸對(duì)FI無顯著性影響(P>0.05), 添加半胱氨酸組FI低于對(duì)照組, 但差異不顯著(P>0.05); 飼料中牛磺酸含量為0.48%和1.06%時(shí)大菱鲆幼魚的飼料效率(FE)高于對(duì)照組(P<0.05), 而蛋氨酸添加組FE略高于對(duì)照組, 但差異不顯著(P>0.05), 半胱氨酸添加組的FE與對(duì)照組無顯著性差異(P>0.05); 飼料中牛磺酸含量為0.48%和1.06%時(shí)蛋白效率比(PER)高于對(duì)照組(P<0.05), 而蛋氨酸添加組與半胱氨酸添加組的PER與對(duì)照組無顯著性差異(P>0.05); 1.06%牛磺酸組的蛋白質(zhì)沉積率(PPV)高于對(duì)照組(P<0.05), 0.48%牛磺酸組及蛋氨酸組的PPV與對(duì)照組相比無顯著性差異(P>0.05), 而半胱氨酸組PPV則低于對(duì)照組(P<0.05)。
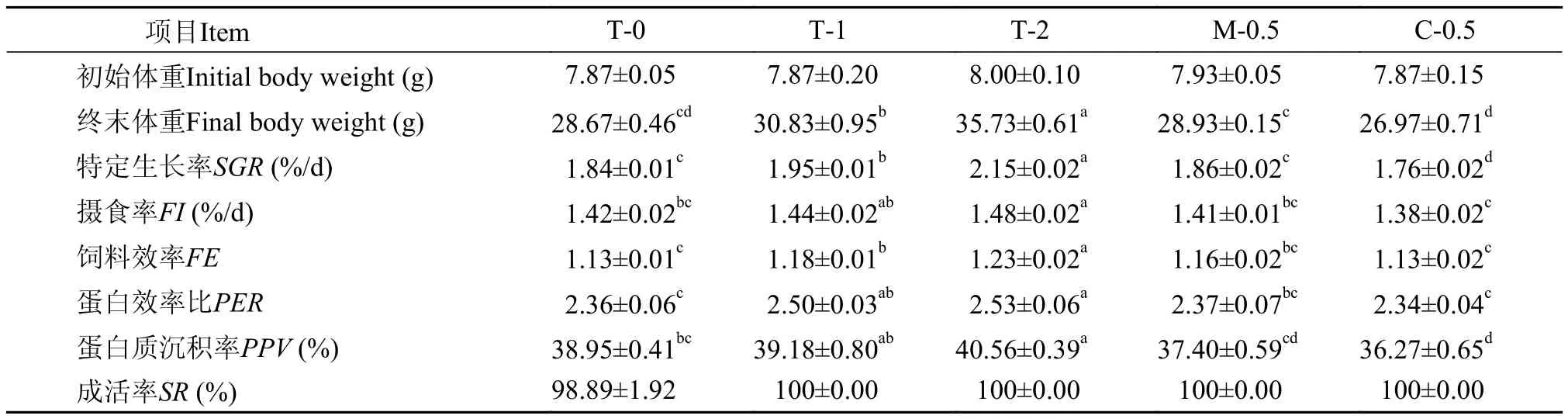
表 4 牛磺酸及相關(guān)氨基酸對(duì)大菱鲆幼魚生長性能的影響Tab. 4 Effects of dietary taurine and its related amino acids on growth performance of juvenile turbot
2.2 牛磺酸及含硫氨基酸對(duì)大菱鲆幼魚魚體化學(xué)組成及形體指標(biāo)的影響
由表 5可知, 與對(duì)照組相比, 0.48%、1.06%牛磺酸組和0.5%蛋氨酸組大菱鲆幼魚魚體水分的變化不顯著(P>0.05), 添加0.5%半胱氨酸可提高魚體水分含量(P<0.05); 隨著飼料中牛磺酸含量的升高,魚體粗蛋白質(zhì)含量提高(P<0.05), 蛋氨酸添加組魚體粗蛋白質(zhì)含量高于對(duì)照組(P<0.05), 而半胱氨酸添加組的魚體粗蛋白質(zhì)含量低于對(duì)照組(P<0.05);魚體粗脂肪含量與粗蛋白質(zhì)含量表現(xiàn)出相同的變化趨勢(shì), 0.48%、1.06%牛磺酸組和0.5%蛋氨酸組魚體的粗脂肪含量升高(P<0.05), 而半胱氨酸添加組魚體的粗脂肪含量與對(duì)照組顯著不差異(P>0.05); 與對(duì)照組相比, 0.48%、1.06%牛磺酸組和0.5%蛋氨酸組魚體的粗灰分含量降低(P<0.05), 而添加0.5%半胱氨酸對(duì)魚體粗灰分含量無顯著性影響(P>0.05)。
T-1、T-2和M-0.5的內(nèi)臟指數(shù)均低于對(duì)照組(P<0.05), 而C-0.5組的內(nèi)臟指數(shù)與對(duì)照組無顯著差異(P>0.05); T-1、T-2和M-0.5大菱鲆幼魚肝指數(shù)與對(duì)照組無顯著差異(P>0.05), 而添加0.5%半胱氨酸則會(huì)降低肝指數(shù)(P<0.05); T-1、T-2組的肥滿度略高于對(duì)照組, C-0.5組肥滿度略低于對(duì)照組, 但無顯著差異(P>0.05), M-0.5組肥滿度高于對(duì)照組(P<0.05)。
2.3 飼料中牛磺酸及相關(guān)含硫氨基酸對(duì)TauT mRNA表達(dá)的影響
由表 6不同組織中TauT mRNA相對(duì)表達(dá)量可知, 與對(duì)照組相比, 飼料中牛磺酸含量為0.48%、1.06%時(shí)均可降低大菱鲆幼魚肝臟、腦和眼組織中的TauT mRNA相對(duì)表達(dá)量(P<0.05), 且隨著飼料中牛磺酸含量增加, TauT mRNA相對(duì)表達(dá)量降低(P<0.05); 飼料中添加0.5%蛋氨酸大菱鲆幼魚肝臟、腦和眼組織中的TauT mRNA相對(duì)表達(dá)量高于T-0、T-1及T-2組(P<0.05); 0.5%半胱氨酸添加組大菱鲆幼魚肝臟、腦和眼組織中的TauT mRNA相對(duì)表達(dá)量高于T-0、T-1及T-2組(P<0.05), 但與M-0.5組相比差異不顯著(P>0.05)。
3 討論
3.1 牛磺酸及相關(guān)氨基酸對(duì)大菱鲆幼魚生長性能的影響
飼料中添加適量牛磺酸能夠提高軍曹魚(Rachycentron canadum)、虹鱒(Oncorhynchus mykiss)、真鯛(Pagrus major)、羅非魚(Oreochromis niloticus)、草魚(Ctenopharyngodon idellus)、牙鲆(Paralichthys olivaceus)和大菱鲆等魚類的生長性能, 因此牛磺酸是一種條件性必需氨基酸[2,5,6,14—22], 這些研究與本實(shí)驗(yàn)的結(jié)果相似, 在本實(shí)驗(yàn)中, 大菱鲆幼魚的終末體重、特定生長率、攝食率、飼料效率和蛋白效率比隨著飼料中牛磺酸含量的增加而升高, 說明飼料中添加適量牛磺酸可改善大菱鲆幼魚的生長性能。齊國山研究表明6.3 g的大菱鲆幼魚飼料中牛磺酸的最適需求量為1.0%[10]。高植物蛋白飼料中因缺乏牛磺酸, 會(huì)導(dǎo)致魚類攝食后生長下降,而在本研究中T-1、T-2組飼料中補(bǔ)充牛磺酸后, 飼料中牛磺酸含量分別為0.48%、1.06% (表 1), 均高于對(duì)照組的0.08%, 從而促進(jìn)生長; 其中T-2組牛磺酸含量接近其需求量, 滿足其最大生長需求, 因此獲得最高特定生長率。在本研究中, 攝食率和飼料效率與特定生長率變化趨勢(shì)一致, 表明生長的改善可能是攝食率和飼料效率的提高引起的, 這在牙鲆[17]、大菱鲆[10]上得到類似結(jié)果。牛磺酸作為魚類的優(yōu)良誘食劑, 在草魚、牙鲆等種類中已有報(bào)道[16,17], 它可以通過刺激魚類的嗅覺和味覺而促進(jìn)魚類攝食[23], 提高攝食率。牛磺酸可以調(diào)節(jié)消化酶的活性, 增強(qiáng)魚體對(duì)飼料中營養(yǎng)物質(zhì)的消化吸收[24],從而提高飼料效率。在本實(shí)驗(yàn)中牛磺酸的添加量與實(shí)際檢測(cè)值不符, 可能是當(dāng)時(shí)所用的牛磺酸實(shí)際含量并未達(dá)到所標(biāo)注的濃度, 或者與飼料制作過程有關(guān)。因?yàn)樵陲暳现谱鲿r(shí), 選用的國藥試劑牛磺酸,其中一瓶是已經(jīng)開口用過的, 并在-20℃保存一年,當(dāng)時(shí)并未測(cè)定這一瓶牛磺酸純度是否依然符合標(biāo)準(zhǔn), 而且飼料制作成功后未能及時(shí)檢測(cè)氨基酸含量,而是在養(yǎng)殖實(shí)驗(yàn)結(jié)束后進(jìn)行的氨基酸成分分析, 從而導(dǎo)致飼料中牛磺酸實(shí)際含量與添加量不符, 但具體原因尚待進(jìn)一步研究。
本實(shí)驗(yàn)在飼料中添加0.5%蛋氨酸未能顯著提高大菱鲆幼魚的生長性能, 與此相似, 飼料中添加蛋氨酸并未提高虹鱒魚的生長、攝食和飼料轉(zhuǎn)化率[18]。魚體內(nèi)蛋氨酸主要用來合成蛋白質(zhì), 作為甲基供體及解毒作用[25,26], 并非通過誘食作用、改善飼料利用率提高生長。對(duì)大黃魚(Pseudosciaena crocea R.)[27]、羅非魚[28]的研究表明, 飼料中添加蛋氨酸表現(xiàn)出顯著的促生長作用, 這可能與魚的種類和飼料組成的差異有關(guān)。Ma等[29]研究表明大菱鲆幼魚對(duì)飼料中蛋氨酸需求量以1.59%適宜, 本實(shí)驗(yàn)M-0.5組飼料的蛋氨酸含量為1.26%, 未達(dá)到其最適需求量, 這也可能是造成其生長未能顯著提高的原因; 齊國山[10]研究表明, 在高植物蛋白飼料中添加1%的蛋氨酸顯著促進(jìn)大菱鲆生長, 但攝食率及飼料效率無顯著提高, 這可能與實(shí)驗(yàn)飼料組成差別有關(guān); Wang等[30]研究表明, 飼料中添加0.5%及1.5%的蛋氨酸均顯著提高大菱鲆的特定生長率, 出現(xiàn)該結(jié)果可能是其實(shí)驗(yàn)飼料中蛋氨酸含量分別為1.53%、2.53%, 均高于本實(shí)驗(yàn)蛋氨酸組飼料中1.26%的蛋氨酸含量, 接近或達(dá)到大菱鲆的最適需求量1.59%所致。在本實(shí)驗(yàn)中C-0.5組大菱鲆幼魚終末體重、攝食率、飼料效率與對(duì)照組無顯著差異, 而特定生長率顯著低于對(duì)照組, 這與Matsunari等[31]、齊國山[10]的研究結(jié)果相似, 說明添加半胱氨酸不能提高大菱鲆的生長性能。
3.2 牛磺酸及相關(guān)氨基酸對(duì)大菱鲆幼魚魚體化學(xué)組成及形體指標(biāo)的影響
在本實(shí)驗(yàn)中, 添加牛磺酸可顯著提高魚體的蛋白質(zhì)含量, 與在吉富羅非魚(Oreochromis nilotictus)和斜帶石斑魚(Epinephelus coioides)[15]、虹鱒[32]、鯽[33]和大菱鲆[19]中的研究一致。一方面牛磺酸可以刺激蛋白質(zhì)消化相關(guān)的激素和酶的分泌, 提高蛋白質(zhì)利用率; 另一方面飼料中補(bǔ)充牛磺酸后, 可使更多的含硫氨基酸參與到蛋白質(zhì)合成過程中, 以此提高魚體蛋白質(zhì)含量[32,33]。研究顯示, 飼料中添加牛磺酸顯著提高虹鱒[5]、羅非魚[15]、鯽[34]體內(nèi)的脂肪含量, 這些結(jié)果與本實(shí)驗(yàn)結(jié)果一致; 牛磺酸參與脂類代謝, 主要通過與膽酸結(jié)合, 生成膽汁酸, 進(jìn)而促進(jìn)脂肪乳化、增加脂肪酶活性, 促進(jìn)了脂肪和脂溶性物質(zhì)的消化吸收率, 從而增加魚體內(nèi)脂肪含量[34,35]。與本實(shí)驗(yàn)結(jié)果相反, 補(bǔ)充牛磺酸石斑魚體內(nèi)脂肪含量顯著降低[15]; 齊國山添加牛磺酸6.3 g和48 g的大菱鲆全魚脂肪含量降低, 認(rèn)為是牛磺酸參與了膽汁酸以外的其他代謝途徑[10]; 對(duì)大西洋鮭的研究表明, 添加牛磺酸降低了魚體的脂肪沉積, 其認(rèn)為該結(jié)果可能與牛磺可提高多胺濃度有關(guān), 而多胺代謝與脂肪沉積有關(guān)[36]。對(duì)于牛磺酸與魚類脂肪沉積的關(guān)系, 不同的研究得出的結(jié)果不同, 其根本原因有待深入研究。在本實(shí)驗(yàn)飼料中添加牛磺酸,大菱鲆魚體肥滿度有升高趨勢(shì), 而內(nèi)臟指數(shù)則顯著降低, 這可能與蛋白質(zhì)、脂肪這些常規(guī)營養(yǎng)成分在體內(nèi)的含量增加, 從而提高魚體的含肉率有關(guān)。

表 5 大菱鲆幼魚魚體化學(xué)組成及形體指標(biāo)(濕重)Tab. 5 Body chemical composition and physical indicators of juvenile turbot (wet weight)

表 6 不同組織中TauT mRNA相對(duì)表達(dá)量Tab. 6 Relative expression levels of TauT mRNA in different tissues
在飼料中添加蛋氨酸可顯著提高魚體的蛋白質(zhì)含量, 在對(duì)奧尼羅非魚(Oreochromis niloticus O.)、印度鯉(Cirrhinus mrigala)的實(shí)驗(yàn)中得到同樣結(jié)果[37,38]。蛋氨酸是魚類的必需氨基酸, 飼料中添加蛋氨酸可提高飼料的營養(yǎng)水平, 提高飼料效率及體蛋白質(zhì)沉積[8]。在本研究中, 蛋氨酸添加組的CF顯著升高, VSI顯著降低, HSI降低(P>0.05), 這與侯華鵬等[39]結(jié)果一致。
3.3 飼料中牛磺酸及相關(guān)氨基酸對(duì)TauT mRNA表達(dá)的影響
牛磺酸是可興奮組織細(xì)胞中最豐富的游離氨基酸, 具有廣泛的生物學(xué)效應(yīng), 主要通過細(xì)胞膜上的牛磺酸轉(zhuǎn)運(yùn)載體轉(zhuǎn)運(yùn)入膜[12]。牛磺酸轉(zhuǎn)運(yùn)載體基因在魚體不同組織中廣泛表達(dá), 研究顯示, 鯉(Cyprinus carpio)的所有組織、鱸檢測(cè)的9個(gè)組織和塞內(nèi)加爾鰨(Solea senegalensis)的12個(gè)組織中均有TauT表達(dá)[13,40,41]。另外, 本實(shí)驗(yàn)室研究表明, TauT在大菱鲆眼、腦、肝、脾、腸、心等組織中均有表達(dá)(衛(wèi)育良, 未發(fā)表)。
影響TauT mRNA表達(dá)的因素有很多, 如食物、環(huán)境滲透壓、激素、蛋白激酶C等[13]。原位雜交顯示攝食低牛磺酸水平飼料的動(dòng)物腎近端小管S3段的TauT表達(dá)上調(diào), 而攝食高牛磺酸水平飼料的動(dòng)物TauT表達(dá)顯著下調(diào)(P<0.05)[42]。大西洋鮭(Salmo salar)幼鮭攝食補(bǔ)充牛磺酸的飼料, TauT mRNA水平顯著低于對(duì)照組[43]。與上述研究結(jié)果相似, 在本研究中隨著飼料中牛磺酸含量增加, 大菱鲆幼魚肝臟、腦和眼3個(gè)組織中的TauT mRNA相對(duì)表達(dá)量顯著降低。有研究表明對(duì)飲食的長期適應(yīng)性調(diào)節(jié)是在mRNA水平進(jìn)行的[41], 飼料中較高的可利用性牛磺酸含量降低了所需的TauT數(shù)量[43]。而在體外培養(yǎng)人HepG2細(xì)胞時(shí), 發(fā)現(xiàn)在牛磺酸富裕的培養(yǎng)基中, TauT的活性及其mRNA表達(dá)水平被下調(diào)[43]。TauT是調(diào)節(jié)細(xì)胞牛磺酸水平的關(guān)鍵因子[45], 牛磺酸主要通過細(xì)胞內(nèi)外轉(zhuǎn)運(yùn)調(diào)整機(jī)體牛磺酸生物“穩(wěn)態(tài)”[46], 因此, TauT mRNA表達(dá)量的降低可能是應(yīng)對(duì)飼料中較高含量的牛磺酸, 維持細(xì)胞內(nèi)牛磺酸含量穩(wěn)定的一種機(jī)制。
本研究顯示在飼料中添加蛋氨酸和半胱氨酸可顯著提高大菱鲆幼魚肝臟、腦和眼3個(gè)組織中的TauT mRNA相對(duì)表達(dá)量(P<0.05)。塞內(nèi)加爾鰨幼魚的腦、心、眼組織中TauT表達(dá)最高, 通常這些部位的牛磺酸濃度很高, 因?yàn)榕;撬嵩谄渲邪l(fā)揮重要的生物學(xué)作用[41]。Watson等[47]研究認(rèn)為, 因?yàn)楦闻K是牛磺酸主要合成部位, 飼料中缺乏牛磺酸應(yīng)首先在肝臟中被識(shí)別, 而缺乏牛磺酸的飼料可能會(huì)導(dǎo)致肝臟中TauT轉(zhuǎn)錄的升高; 牛磺酸在肝臟中合成后,經(jīng)由血液循環(huán)及TauT被其他組織攝取, 而其他組織的TauT表達(dá)應(yīng)上調(diào), 以促進(jìn)在飼料中牛磺酸水平較低情況下的牛磺酸回收。有研究顯示牛磺酸吸收的增加是TauT表達(dá)量上調(diào)的結(jié)果[44]。齊國山[10]研究表明, 大菱鲆體內(nèi)可能存在將蛋氨酸和胱氨酸轉(zhuǎn)化為牛磺酸的途經(jīng)。因此, 筆者推測(cè)在本研究中,大菱鲆幼魚可能能夠利用飼料中的蛋氨酸和半胱氨酸合成牛磺酸, 而TauT mRNA表達(dá)量上調(diào)是將肝臟中合成的牛磺酸轉(zhuǎn)運(yùn)至眼、腦這些部位, 以利于其生理功能的發(fā)揮。但是, 由于大菱鲆幼魚的牛磺酸合成水平可能較低[10], 所合成的牛磺酸量不能滿足攝食高植物蛋白飼料的大菱鲆幼魚的最大生長需求, 因此, 并未表現(xiàn)出顯著的促生長作用。在對(duì)人的研究中發(fā)現(xiàn), TauT與胎兒生長、胎兒發(fā)育遲緩的發(fā)病機(jī)制及出生體重有關(guān)[48], 說明TauT可能通過某種途徑對(duì)機(jī)體生長產(chǎn)生作用。Pinto等[41]認(rèn)為, 在一個(gè)生長潛力非常高的階段, 高的牛磺酸轉(zhuǎn)運(yùn)能力可能對(duì)幼體生長非常重要, 已有研究指出TauT介導(dǎo)的肌漿網(wǎng)鈣攝取對(duì)肌纖維的興奮—收縮機(jī)制有一定作用, 并誘導(dǎo)骨骼肌細(xì)胞攝取氨基酸, 由于氨基酸是成長的基石, 肌肉中高的牛磺酸轉(zhuǎn)運(yùn)能力可能與氨基酸沉積有關(guān), 并造成較高的生長潛力。而本實(shí)驗(yàn)飼料牛磺酸含量與其在眼、腦及肝的TauT mRNA表達(dá)均為反向調(diào)節(jié), 由于本研究未測(cè)定肌肉中TauT mRNA表達(dá), 因此造成反向調(diào)節(jié)的具體原因有待更深入研究。目前, 對(duì)TauT的研究多集中在組織表達(dá)、外源牛磺酸或高滲刺激等對(duì)TauT mRNA表達(dá)量的影響, 而有關(guān)飼料中牛磺酸及相關(guān)含硫氨基酸對(duì)TauT mRNA表達(dá)量及TauT mRNA表達(dá)與魚體生長的關(guān)系還有待進(jìn)一步研究。
綜合分析表明, 在本實(shí)驗(yàn)條件下, 飼料中牛磺酸含量為0.48%、1.06%時(shí)均能夠提高大菱鲆的生長性能; 大菱鲆幼魚體內(nèi)TauT mRNA表達(dá)可能受飼料中牛磺酸、蛋氨酸和半胱氨酸等含硫氨基酸的影響。
[1]Dong X Q, Zhang D M, Ge C X. Research progress of taurine in fish nutrition [J]. China Animal Husbandry & Veterinary Medicine, 2012, 39(6): 125—127 [董曉慶, 張東鳴, 葛晨霞. 牛磺酸在魚類營養(yǎng)上的研究進(jìn)展. 中國畜牧獸醫(yī), 2012, 39(6): 125—127]
[2]Lunger A N, McLean E, Gaylord T G, et al. Taurine supplementation to alternative dietary proteins used in fish meal replacement enhances growth of juvenile cobia (Rachycentron canadum) [J]. Aquaculture, 2007, 271(1): 401—410
[3]Park G S, Takeuchi T, Yokoyama M, et al. Optimal dietary taurine level for growth of juvenile Japanese flounder Paralichthys olivaceus [J]. Fisheries Science, 2002, 68(4): 824—829
[4]Kim S K, Takeuchi T, Yokoyama M, et al. Effect of dietary taurine levels on growth and feeding behavior of juvenile Japanese flounder Paralichthys olivaceus [J]. Aquaculture, 2005, 250(3): 765—774
[5]Gaylord T G, Teague A M, Barrows F T. Taurine supplementation of all-plant protein diets for rainbow trout (Oncorhynchus mykiss) [J]. Journal of the World Aquaculture Society, 2006, 37(4): 509—517
[6]Takagi S, Murata H, Goto T, et al. Necessity of dietary taurine supplementation for preventing green liver symptom and improving growth performance in yearling red sea bream Pagrus major fed nonfishmeal diets based on soy protein concentrate [J]. Fisheries Science, 2010, 76 (1): 119—130
[7]Concei??o L E C, Vander Meeren T, Verreth J A J, et al. Amino acid metabolism and protein turnover in larval turbot (Scophthalmus maximus) fed natural zooplankton or Artemia [J]. Marine Biology, 1997, 129(2): 255—265
[8]Wang L L, Shao Q J. Effects of dietary methionine supplementation on fish growth [J]. Reservoir Fisheries, 2007, 27(1): 108—110 [王蕾蕾, 邵慶均. 飼料中添加蛋氨酸對(duì)魚類生長的影響. 水利漁業(yè), 2007, 27(1): 108—110]
[9]Han C X, Miao C. The function and the metabolism and absorption process of methionine [J]. Modern Journal of Animal Husbandry and Veterinary Medicine, 2013, (9): 75—80 [韓春曉, 苗翠. 蛋氨酸的功能及代謝吸收過程.現(xiàn)代畜牧獸醫(yī), 2013, (9): 75—80]
[10]Qi G S. Effects of dietary taurine, methionine, cystine, serine and cysteamine on growth performance and metabolism of taurine synthesis in turbot [D]. Thesis for Doctor's Degree. Ocean University of China. 2012 [齊國山.飼料中牛磺酸, 蛋氨酸, 胱氨酸, 絲氨酸和半胱胺對(duì)大菱鲆生長性能及牛磺酸合成代謝的影響. 博士學(xué)位論文, 中國海洋大學(xué). 2012]
[11]Cao B H, Chang X Y, Zhang E P, et al. Effect of dietary cysteine supplement levels on growth performance, nitrogen metabolism and serum biochemical indicators of 2-4 month-old rex rabbits [J]. Acta Ecologiae Animalis Domastici, 2012, 33(4): 33—37 [曹寶紅, 常馨月, 張恩平,等. 日糧中添加半胱氨酸對(duì)2—4月齡獺兔生長性能, 氮代謝和血清生化指標(biāo)的影響. 家畜生態(tài)學(xué)報(bào), 2012, 33(4): 33—37]
[12]Zhang X L, Deng H, Chen C, et al. Taurine transporter: Regulation mechanism and significance in animal nutrition studies [J]. Chinese Journal of Animal Nutrition, 2013, 25(10): 2222—2230 [張小龍, 鄧歡, 陳澄, 等. 牛磺酸轉(zhuǎn)運(yùn)體的調(diào)節(jié)機(jī)制及其在動(dòng)物營養(yǎng)研究中的意義.動(dòng)物營養(yǎng)學(xué)報(bào), 2013, 25(10): 2222—2230]
[13]Zheng W X. Cloning of cDNA and expression analysis of TauT in Lateolabrax japonicas [D]. Thesis for Master's Degree. Ningbo University. 2012 [鄭偉賢. 鱸魚TauT基因的cDNA克隆及組織表達(dá). 碩士學(xué)位論文, 寧波大學(xué). 2012]
[14]Lu Y B, You C H, Wang S Q, et al. Physiological changes in Siganus canaliculatus after shallow water stress and the anti-stress effects of taurine [J]. Acta Hydrobiologica Sinica, 2014, 38(1): 68—74 [盧玉標(biāo), 游翠紅, 王樹啟, 等. 淺水應(yīng)激后黃斑藍(lán)子魚生理指標(biāo)變化及牛磺酸的抗應(yīng)激作用. 水生生物學(xué)報(bào), 2014, 38(1): 68—74]
[15]Wang H W. Effect of dietary taurine level on the growth performance of GIFT tilapia Oreochromis nilotictus and grouper Epinephelus coioides [D]. Thesis for Master's Degree. Jimei University. 2013 [王和偉. 飼料牛磺酸水平對(duì)吉富羅非魚和斜帶石斑魚生長的影響. 碩士學(xué)位論文, 集美大學(xué). 2013]
[16]Luo L, Wang L, Long Y, et al. Study on effects of taurine on growth performance of grass carp [J]. Feed Industry, 2005, 26(12): 22—24 [羅莉, 王琳, 龍勇, 等. 牛磺酸對(duì)草魚生長效應(yīng)研究. 飼料工業(yè), 2005, 26(12): 22—24]
[17]Kim S K, Takeuchi T, Akimoto A, et al. Effect of taurine supplemented practical diet on growth performance and taurine contents in whole body and tissues of juvenile Japanese flounder Paralichthys olivaceus [J]. Fisheries Science, 2005, 71(3): 627—632
[18]Gaylord T G, Barrows F T, Teague A M, et al. Supplementation of taurine and methionine to all-plant protein diets for rainbow trout (Oncorhynchus mykiss) [J]. Aquaculture, 2007, 269(1): 514—524
[19]Qi G S, Ai Q H, Mai K S, et al. Effects of dietary taurine supplementation to a casein-based diet on growth performance and taurine distribution in two sizes of juvenile turbot (Scophthalmus maximus L.) [J]. Aquaculture, 2012, (358—359): 122—128
[20]Yokoyama M, Takeuchi T, Park G S, et al. Hepatic cysteinesulphinate decarboxylase activity in fish [J]. Aquaculture Research, 2001, 32(Suppl. 1): 216—220
[21]Wang Q R, Takeuchi T, Chen Z X. Feeding results on red drum Sciaenop socellatus larvae rearing with taurine-enriched rotifers [J]. Journal of Jimei University (Natural Science), 2006, 11(4): 301—304 [王秋榮, 竹內(nèi)俊郎, 陳振湘. 牛磺酸強(qiáng)化輪蟲對(duì)眼斑擬石首魚仔魚的投喂效果. 集美大學(xué)學(xué)報(bào)(自然科學(xué)版), 2006, 11(4): 301—304]
[22]Takeuchi T, Park G S, Seikai T, et al. Taurine content in Japanese flounder Paralichthys olivaceus T. & S. and red sea bream Pagrus major T. & S. during the period of seed production [J]. Aquaculture Research, 2001, 32(Suppl. 1): 244—248
[23]Qiu X C, Zhao H X, Wei Z Q. The feeding attractant activity of taurine to common carp [J]. Hebei Fisheries, 2006, (8): 10—11 [邱小琮, 趙紅雪, 魏智清. 牛磺酸對(duì)鯉魚誘食活性的初步研究. 河北漁業(yè), 2006, (8): 10—11]
[24]Gao C S, Fan G L, Wang Y L. Effect of taurine on growth performance and digestive enzyme activity of carp [J]. Chinese Agricultural Science Bulletin, 2007, 23(6): 645—647 [高春生, 范光麗, 王艷玲. 牛磺酸對(duì)黃河鯉魚生長性能和消化酶活性的影響. 中國農(nóng)學(xué)通報(bào), 2007, 23(6): 645—647]
[25]Li P, Mai K S, Trushenski J, et al. New developments in fish amino acid nutrition: towards functional and environmentally oriented aquafeeds [J]. Amino Acids, 2009, 37(1): 43—53
[26]Tan X Z, Rong F M. A brief analysis of the influence of methionine on the animal production performance [J]. Shandong Journal of Animal Science and Veterinary Medicine, 2010, (7): 90—92 [譚興智, 榮鳳梅. 淺析蛋氨酸對(duì)動(dòng)物生產(chǎn)性能的影響. 山東畜牧獸醫(yī), 2010, (7): 90—92]
[27]Mai K S, Wan J L, Ai Q H, et al. Dietary methoinine requirement of large yellow croaker, Pseudosciaena crocea R. [J]. Aquaculture, 2006, 253(1—4): 564—572
[28]Leng X J, Tian J, Chen B A, et al. Comparison study on utilizing crystalline or coated methionine by tilapia (Oreochromis niloticus×O. aureus) [J]. Acta Hydrobiologica Sinica, 2013, 37(2): 235—242 [冷向軍, 田娟, 陳丙愛,等. 羅非魚對(duì)晶體蛋氨酸, 包膜蛋氨酸利用的比較研究.水生生物學(xué)報(bào), 2013, 37(2): 235—242]
[29]Ma R, Hou H P, Mai K S, et al. Comparative study on the effects of L-methionine or 2-hydroxy-4-(methylthio) butanoic acid as dietary methionine source on growth performance and anti-oxidative responses of turbot (Psetta maxima) [J]. Aquaculture, 2013, (412—413): 136—143
[30]Wang Q C, He G, Wang X, et al. Dietary sulfur amino acidmodulations of taurine biosynthesis in juvenile turbot (Psetta maxima) [J]. Aquaculture, 2014, (422—423): 141—145
[31]Matsunari H, Furuita H, Yamamoto T, et al. Effect of dietary taurine and cystine on growth performance of juvenile red sea bream Pagrus major [J]. Aquaculture, 2008, 274(1): 142—147
[32]Xu Q Y, Xu H, Zheng Q S, et al. Effects of taurine on growth, body composition and immunity of rainbow trout juvenile [J]. Chinese Journal of Animal Nutrition, 2007, 19(5): 544—548 [徐奇友, 許紅, 鄭秋珊, 等. 牛磺酸對(duì)虹鱒仔魚生長、體成分和免疫指標(biāo)的影響. 動(dòng)物營養(yǎng)學(xué)報(bào), 2007, 19(5): 544—548]
[33]Zhao X F, Jia L, Zhou J P. Effect of taurine on protein digestibility of crucian carp [J]. Journal of Aquaculture, 2006, 27(5): 5—6 [趙小峰, 賈麗, 周劍平. 牛磺酸對(duì)鯽魚蛋白質(zhì)消化吸收的影響. 水產(chǎn)養(yǎng)殖, 2006, 27(5): 5—6]
[34]Zhao X F, Wang Z Y, Zhou J P, et al. Effect of taurine on fat digestibility of crucian carp [J]. Fisheries Science, 2007, 26(8): 453—454 [趙小鋒, 王治業(yè), 周劍平, 等. 牛磺酸對(duì)鯽魚脂肪消化吸收的影響. 水產(chǎn)科學(xué), 2007, 26(8): 453—454]
[35]Mizushima S, Nara Y, Sawamura M. Effects of oral taurine supplementation on lipids and symapathetic nerve tone [J]. Advances in Experimental Medicine and Biology, 1996, 403: 615—622
[36]Espe M, Ruohomen K, EI-Mowafi A. Effect of taurine supplementation on the metabolism and body lipid-toprotein ratio in juvenile atlantic salmon (Salmo salar) [J]. Aquaculture Research, 2012, 43(3): 349—360
[37]Lin S M, Mai K S, Tan B P. Influence of practical diet supplementation with free D-methionine on the growth and body composition in tilapia Oreochromis niloticus × O. aureus [J]. Acta Hydrobiologica Sinica, 2008, 32(5): 741—749 [林仕梅, 麥康森, 譚北平. 實(shí)用飼料中添加結(jié)晶蛋氨酸對(duì)羅非魚生長、體組成的影響. 水生生物學(xué)報(bào), 2008, 32(5): 741—749]
[38]Ahmed I, Khan M A, Jafri A K. Dietary methionine requirement of fingerling Indian major carp, Cirrhinus mrigala (Hamilton) [J]. Aquaculture International, 2003, 11(5): 449—462
[39]Hou H P, Ma R, Zhang W B, et al. The influence of overdose dietary DL-methionine or HMTBA on growth, feed utilization and anti-oxidant responses of turbot (Scophthalmus maximus L.) [J]. Periodical of Ocean University of China, 2012, 42(3): 36—43 [侯華鵬, 馬睿, 張文兵,等. 飼料中過量添加晶體蛋氨酸和蛋氨酸羥基類似物對(duì)大菱鲆生長、飼料利用和抗氧化反應(yīng)的影響. 中國海洋大學(xué)學(xué)報(bào), 2012, 42(3): 36—43]
[40]Takeuchi K, Toyohara H, Sakaguchi M. A hyperosmotic stress-induced mRNA of carp cell encodes Na+-and Cl--dependent high affinity taurine transporter [J]. Biochi-mica et Biophysica Acta (BBA)-Biomembranes, 2000, 1464(2): 219—230
[41]Pinto W, R?nnestad I, Jordal A E O, et al. Cloning, tissue and ontogenetic expression of the taurine transporter in the flatfish Senegalese sole (Solea senegalensis) [J]. Amino Acids, 2012, 42(4): 1317—1327
[42]Matsell D G, Bennett T, Han X, et al. Regulation of the taurine transporter gene in the S3 segment of the proximal tubule [J]. Kidney International, 1997, 52(3): 748—754
[43]Zarate J M, Bradley T M. Molecular cloning and characterization of the taurine transporter of Atlantic salmon [J]. Aquaculture, 2007, 273(2): 209—217
[44]Satsu H, Terasawa E, Hosokawa Y, et al. Functional characterization and regulation of the taurine transporter and cysteine dioxygenase in human hepatoblastoma HepG2 cells [J]. Biochemical Society, 2003, 375(2): 441—447
[45]Ito T, Fujio Y, Schaffer S W, et al. Involvement of Transcriptional Factor TonEBP in the Regulation of the Taurine Transporter in the Cardiomyocyte [M]. Taurine 7. Springer New York. 2009, 523—532
[46]Yun B. Effects of dietary cholesterol, taurine and soya saponins on growth performance and cholesterol metabolism in juvenile turbot (Scophthalmus maximus L.) fed high plant protein [D]. Thesis for Doctor's Degree. Ocean University of China. 2012 [贠彪. 在高植物蛋白飼料中添加膽固醇、牛磺酸和大豆皂甙對(duì)大菱鲆(Scophthalmus maximus L.)生長性能和膽固醇代謝的影響. 博士學(xué)位論文, 中國海洋大學(xué). 2012]
[47]Watson A M, Barrows F T, Place A R. Effects of graded taurine levels on juvenile cobia [J]. North American Journal of Aquaculture, 2014, 76(3): 190—200
[48]Chen Z G, He P, Ding X Y, et al. PPARγ stimulates expression of L-type amino acid and taurine transporters in human placentas: the evidence of PPARγ regulating fetal growth [J]. Scientific Reports, 2015, 5(12650): 1—13
EFFECTS OF DIETARY TAURINE AND RELATED AMINO ACIDS ON GROWTH PERFORMANCE AND TAURINE TRANSPORTER (TAUT) MRNA EXPRESSION OF JUVENILE TURBOT (SCOPHTHALMUS MAXIMUS L.)
LIU Xi1,2, LIANG Meng-Qing1, ZHENG Ke-Ke1, WANG Cheng-Qiang1,2and XU Hou-Guo1
(1. Yellow Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Qingdao 266071, China; 2. College of Fisheries and Life Sciences, Shanghai Ocean University, Shanghai 201306, China)
A 10-week feeding experiment in indoor flowing-through water system was conducted to evaluate the effects of taurine, methionine and cysteine in diets on growth performance and taurine transporter (TauT) mRNA expression of juvenile turbot (Scophthalmus maximus L.) [initial weight: (7.90±0.07) g]. The basal diet was supplemented with 0, 1%, 2% taurine, 0.5% methionine and 0.5% cysteine (Control T-0, T-1, T-2, M-0.5 and C-0.5, respectively) to formulate five isonitrogenous and isolipidic diets. The results showed that the specific growth rate (SGR) and feed efficiency (FE) of T-1 and T-2 group were significantly higher than that of the control group (P<0.05), and the visceral indexes (VSI) of T-1 and T-2 group was significantly lower than that of the control group (P<0.05). Compared with T-0 group, the SGR and FE of M-0.5 group improved (P>0.05), and VSI decreased significantly (P<0.05), while condition factors (CF) increased significantly (P<0.05). Compared with T-0 group, the SGR of C-0.5 group decreased significantly (P<0.05), and no significant differences in FE, VSI and CF were observed (P>0.05) between the two groups. T-1 and T-2 group had significantly lower TauT mRNA relative expression (P<0.05) in liver, brain and eye tissues than those in T-0 group, and the TauT mRNA relative expression in liver, brain and eye tissues decreased significantly (P<0.05) with increasing dietary taurine level. On the contrary, the TauT mRNA relative expression in liver, brain and eye tissues of M-0.5 group was significantly higher (P<0.05) than those in T-0, T-1 and T-2 groups. C-0.5 group had a significantly higher TauT mRNA relative expression (P<0.05) in liver, brain and eye than those in T-0, T-1 and T-2 groups, but no significant differences (P>0.05) were found when compared with M-0.5 group. In conclusion, 0.48%, 1.06% taurine in diets could promote growth performance of juvenile turbot under these experimental conditions. The TauT mRNA expression in juvenile turbot may be affected by dietary sulfur-containing amino acids such as taurine, methionine and cysteine.
Taurine; Methionine; Cysteine; Turbot; Taurine transporter (TauT)
S963
A
1000-3207(2017)01-0165-09
10.7541/2017.21
2016-01-04;
2016-04-11
國家自然科學(xué)基金(31340076); 農(nóng)業(yè)公益行業(yè)專項(xiàng)(201303053)資助 [Supported by the National Natural Science Foundation of China (31340076); the Special Fund for Agro-scientific Research in the Public Interest (201303053)]
柳茜(1989—), 女, 河北石家莊人; 碩士; 主要從事水產(chǎn)動(dòng)物營養(yǎng)與飼料研究。E-mail: 371707080@qq.com
梁萌青, 女, 研究員; E-mail: liangmq@ysfri.ac.cn
猜你喜歡
作文·小學(xué)低年級(jí)(2025年2期)2025-02-13 00:00:00
小雪花·小學(xué)生快樂作文(2024年11期)2024-12-31 00:00:00
作文·小學(xué)低年級(jí)(2024年2期)2024-04-29 00:00:00
作文·小學(xué)低年級(jí)(2023年3期)2023-04-29 00:00:00
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
小主人報(bào)(2022年4期)2022-08-09 08:52:06
中學(xué)生數(shù)理化·中考版(2022年11期)2022-02-16 07:01:20
小哥白尼(趣味科學(xué))(2019年6期)2019-10-10 01:01:50
發(fā)明與創(chuàng)新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
