空心菜浮床對魚塘水質和微生物多樣性的影響
2017-01-19 08:44:14李建柱張鵬飛柳堯全夏閏紅馬徐發華中農業大學湖北武漢430070
中國環境科學 2016年10期
關鍵詞:環境
李建柱,侯 杰,張鵬飛,柳堯全,夏閏紅,馬徐發(華中農業大學,湖北 武漢 430070)
空心菜浮床對魚塘水質和微生物多樣性的影響
李建柱,侯 杰,張鵬飛,柳堯全,夏閏紅,馬徐發*(華中農業大學,湖北 武漢 430070)
為從環境微生物的角度探討生物浮床的水質調節機理,采用宏基因組學測序技術和生物信息學手段,分析了環境微生物和根系微生物群落的多樣性.研究結果表明,魚菜共生模式下優勢菌門是變形菌門(Proteobacteria)、藍細菌門(Cyanobacteria)、擬桿菌門(Bacteroidetes)、放線菌門(Actinobacteria)、綠彎菌門(Chloroflexi)和厚壁菌門(Firmicutes)等,主要菌屬是聚球藻屬(Synechococcus)、hgcI_clade、CL500-29_marine_group、芽孢桿菌屬(Bacillus)和分支桿菌屬(Mycobacterium)等;空心菜根系微生物群落中最豐富的優勢菌門是變形菌門和藍細菌門,主要菌屬是紅細菌屬(Rhodobacter)、噬氫菌屬(Hydrogenophaga)和聚球藻屬等.研究結果還表明低覆蓋率(3.5%)的空心菜生物浮床能夠增加水體的溶解氧含量,降低水體中含氮化合物的濃度和改善養殖環境中的微生態平衡,提高有益菌和氮循環細菌的含量.
空心菜浮床;環境微生物多樣性;水質;16S rDNA
池塘養殖是中國主要的淡水水產養殖方式,由養殖導致的環境污染日益引起了人們的擔憂.廢氮積累是水產養殖的一個主要缺點[1],這些廢物通常是養殖生物以氨氮的形式排泄[2-3],氨氮對養殖生物具有長期重毒作用[4-5].另外,亞硝酸氮對養殖生物也具有毒性作用[6-7].因此,尋求一個高效科學健康的治理養殖水污染的措施或更加可持續發展的養殖模式是亟待解決的問題.
魚菜共生模式是一種結合生物浮床技術的新型健康生態養殖模式.研究表明生物浮床技術革命性地推進了水質凈化技術的發展,提高了空間利用效率,在凈化水質的同時,也可以產生一定的生態效益和經濟效益[8-10],具有非常廣泛的應用前景.目前,對魚菜共生模式的研究多側重于研究選用不同浮床植物對污染水體的凈化效果[11-14],而對浮床植物根系微生物的群落結構和多樣性及其變化研究較少,且沒有從整個環境中全部微生物的角度對生物浮床水質調節機理做出一個科學的解釋.因此,研究魚菜共生模式中環境微生物和浮床植物根系微生物群落結構的多樣性,對深入探討生物浮床的水體調節機理具有非常重要的意義.本研究旨在通過宏基因組學等分子生物學方法,揭示生物浮床對魚菜共生模式中微生物多樣性的影響,并從微生物的角度對生物浮床水質調節機理做出一個科學的解釋,為養殖水體生物調控技術的推廣與應用提供一定的理論基礎.
1 實驗材料與方法
1.1 實驗地點
實驗點位于湖北省紅安縣中海農業科技有限公司高橋養殖基地,選取6個基本條件(養殖年限、面積、水深、水源與魚類放養模式等)一致的草魚精養池塘,其中3個水面上架設覆蓋率為3.5% 的空心菜生物浮床構建魚菜共生模式作為實驗組,3個池塘為空白對照組.為防止草魚攝食植物根系,本實驗制作的生物浮床植物載體采用了雙層PE網,把空心菜移植在上層網片上,成功防止植物根系被魚類攝食.本試驗周期為2015-06-10~2015-08-23.池塘面積為0.80hm2,平均水深1.80m.每天用商業魚飼料(草魚707系列,海大集團)投喂2次(08:30和15:30).
1.2 水質監測方法
水質理化參數溶解氧(DO)、溫度(T)和pH用HACH HQ 40d便攜式多參數水質分析儀實時測得.
定期采取離水面50cm深處的水層水樣,在每個養殖池塘選取5個取樣點取同等體積的水樣,混合均勻后即為全層多點取樣.測定方法參照《水質—中華人民共和國國家標準》測定,其中總氮(TN)用堿性過硫酸鉀消解,紫外分光光度法測定;硝態氮測定采用酚二磺酸分光光度法;總氨氮(TAN)采用納氏試劑比色法;亞硝態氮采用磺胺試劑分光光度計比色法[15-18].
實驗組和對照組的各個水質參數平均15d監測1次,共5次,編號分別為T和C.
1.3 樣品收集與DNA提取
本實驗所有菌體樣品均于2015-8-03上午采集.
水體菌體收集:采用5 點取樣法,用高溫滅菌的有機玻璃取水器采取每個養殖池塘水深50cm的池水,每個樣取樣量大體一致,均勻混合后裝入滅菌的三角燒瓶,隨后將50mL水樣用0.22pm的濾膜過濾后,將濾膜置于無菌冷凍管中.泥樣采集:采用3點取樣法,3個采集點均位于投食區,用采泥器采集每個池塘采樣點的底泥,混勻并置于無菌冷凍管中.根系菌體收集:隨機剪取空心菜根系,懸浮于PBS緩沖溶液中,所得的懸浮液即為根面菌液,搖勻,然后用0.22pm的濾膜過濾100mL根面菌液,將濾膜置于無菌冷凍管中.上述所有樣品均置于-20℃保存備用,直至提取DNA.
實驗組水體和沉積物的樣品編號分別為ST(ST1~ST3)和NT(NT1~NT3),對照組水體和沉積物的樣品編號SC(SC1~SC3)和NC(NC1~NC3),空心菜根系樣品編號為R(R1~R3).使用Soil DNA Kit試劑盒(OMEGA,美國)提取上述所有樣品的DNA.所有樣品的 DNA 提取步驟均參照Soil DNA Kit試劑盒說明書,并對提取到的樣品基因組 DNA用1% 瓊脂糖凝膠電泳檢測質量.
1.4 高通量測序分析
針對細菌16S rRNA基因V3+V4區設計含barcode的特異引物338F/806R,引物序列為:338F(5'-ACTCCTACGGGAGGCAGCA-3'),806R(5'-GGACTACHVGGGTWTCTAAT-3').PCR擴增采用rTaq DNA Polymerase-20μL反應體系:10×Buffer 2μL,2.5mmol/L dNTPs 2μL,引物338F/806R各0.8μL,Taq酶0.2μL, BSA 0.2μL,模板DNA10ng,補ddH2O至20μL. PCR條件:95℃變性 3min;95℃ 30s,55℃ 30s,72℃ 45s,27個循環;最后72℃ 10min延伸(PCR儀:ABⅠGeneAmp? 9700型).
PCR產物經過2%瓊脂糖凝膠電泳檢測,條帶清晰,片段大小合適(430bp左右),使用AxyPrepDNA凝膠回收試劑盒(Axygen,美國)切膠回收PCR產物,以Tris-HCl 洗脫后連接“Y”接頭,使用磁珠篩選去除接頭自連片段,按照每個樣品測一萬條序列加入1ng PCR產物的標準富集PCR產物,然后用0.1mol/L的NaOH 溶液變性,獲得單鏈DNA片段,構建測序文庫后在16S-338F-806R Miseq平臺進行高通量測序(美吉生物,上海).
1.5 數據分析
對獲得的測序數據,用 MOTHUR 軟件結合EXCEL和SPSS 軟件進行數據統計和分析.數據的統計分析以每個樣品中可操作的分類單元(OUT,分類單元是分類工作中的客觀操作單位,有特定的名稱和分類特征,是指具體的分類群,可能是菌株、種或屬等,將每個分類單位統稱為一個操作分類單位)作為分類和計算的依據,并設置差異顯著性水平P = 0.05,當P<0.05時表示顯著性差異.
2 結果
2.1 水質指標
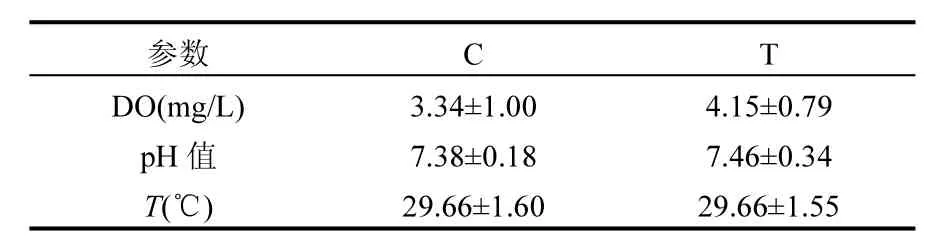
表1 水質的物理參數Table 1 Physical parameter of water in Control and Test
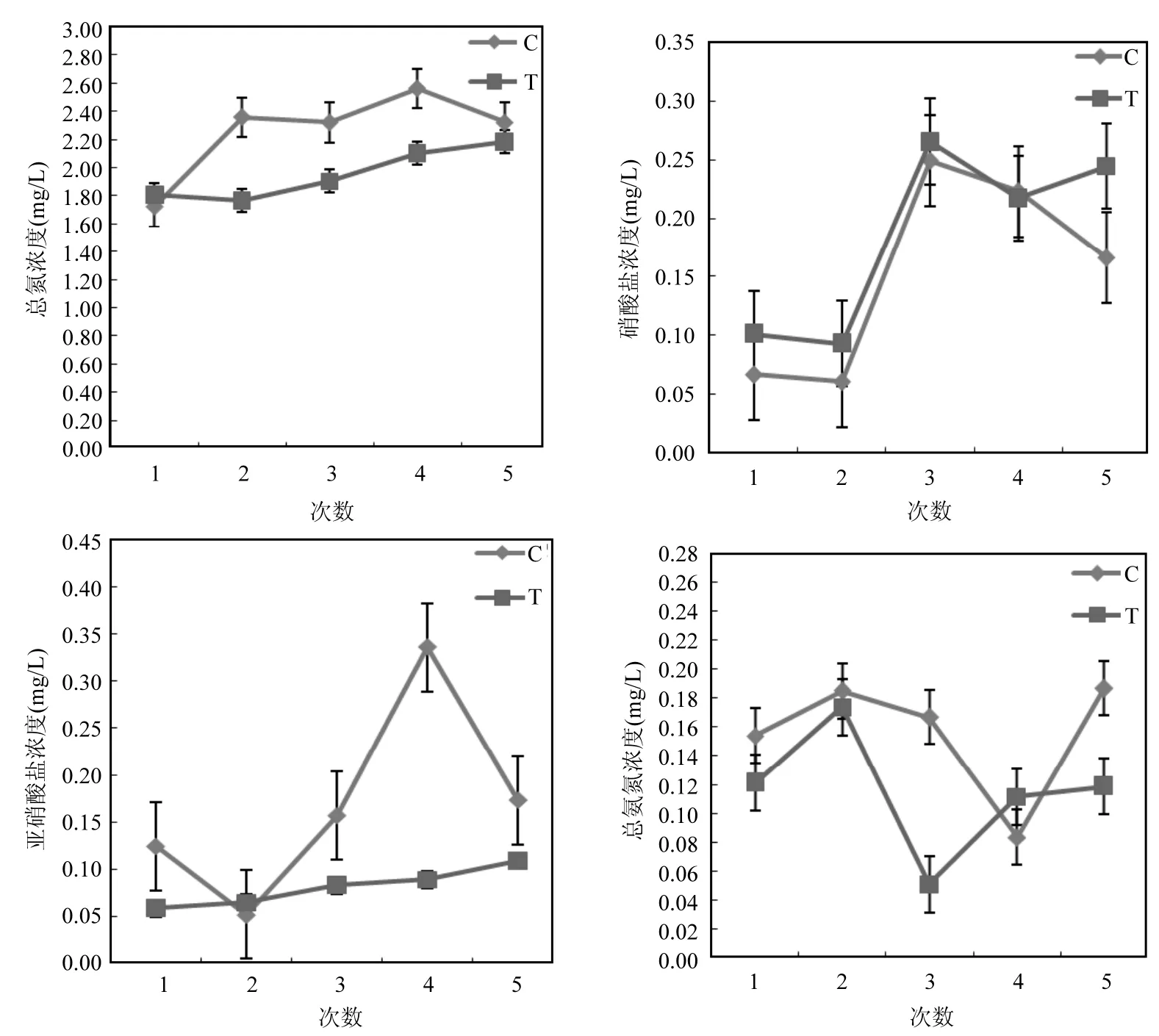
圖1 試驗期間總氮、總氨氮、硝酸鹽和亞硝酸鹽濃度的變化Fig.1 Variration of TN, TAN, nitrate, and nitrite concentrations during the study period
實驗組和對照組的DO濃度、pH值和溫度的監測結果如表1所示,且各參數之間并沒有顯著性差異(P>0.05).由研究結果可知魚菜共生系統中的pH值和溫度與非魚菜共生系統的基本相同;但魚菜共生模式下水體的DO濃度明顯較高,分別為(4.15±0.79)mg/L和(3.34±1.00)mg/L.說明魚菜共生系統中水質的理化性質比非魚菜共生系統中更優良,更有利于魚類養殖.
實驗過程中,實驗組和對照組的總氮、硝酸鹽、亞硝酸鹽和總氨氮的濃度變化如圖1所示,且各參數之間并沒有顯著性差異(P>0.05).由研究結果可知實驗組TN濃度的波動范圍為1.76~2.18mg/L,對照組TN濃度的波動范圍為1.72~2.32mg/L,且2個組別的總氮濃度都呈現出增加的趨勢,實驗組TN的濃度一直低于對照組TN的濃度;實驗組亞硝酸鹽濃度的波動范圍為0.06~0.11mg/L,對照組亞硝酸鹽濃度的波動范圍為0.05~0.34mg/L,實驗組亞硝酸鹽的濃度明顯低于對照組亞硝酸鹽的濃度;實驗組TAN濃度的波動范圍為0.05~0.17mg/L,對照組TAN濃度的波動范圍為0.08~0.19mg/L;實驗組硝酸鹽濃度的波動范圍為0.09~0.27mg/L,對照組硝酸鹽濃度的波動范圍為0.06~0.25mg/L,實驗組硝酸鹽的濃度高于對照組硝酸鹽的濃度(圖1).這些結果說明盡管生物浮床能夠降低水環境中的總氮、亞硝酸鹽和總氨氮濃度,有利于保持養殖水,境的良好,但養殖過程中含氮化合物會不斷累積,需要適時調節水質,保證健康養殖.
2.2 樣品細菌群落多樣性
對各個樣品的有效序列進行系譜分類,以97%相似性水平為標準劃分操作分類單元(OUT),SC、ST、NC、NT和R樣品數據分別可劃分為833、962、1989、2164和986個OUT(表2).通過計算在同一樣本中檢測到的隨機選擇擴增子序列的覆蓋率進行評估抽樣的完整性.OTUs的覆蓋率的范圍是99.17%~99.63%(表2),表明本研究樣品中的大多數微生物種類都被檢測到了.由樣品的菌群豐度指數(Ace和Chao)可知實驗組水體和沉積物的微生物豐度高于對照組;由樣品的多樣性指數(Shannon和Simpson)可知實驗組水體的微生物多樣性低于對照組,實驗組沉積物的微生物多樣性高于對照組(表2).這些研究結果表明與非魚菜共生模式相比,魚菜共生模式中養殖水環境中微生物具有高豐度和低多樣性,沉積物中微生物具有高豐度和高多樣性,進而表明空心菜生物浮床有效改變了養殖環境中的微生態平衡.
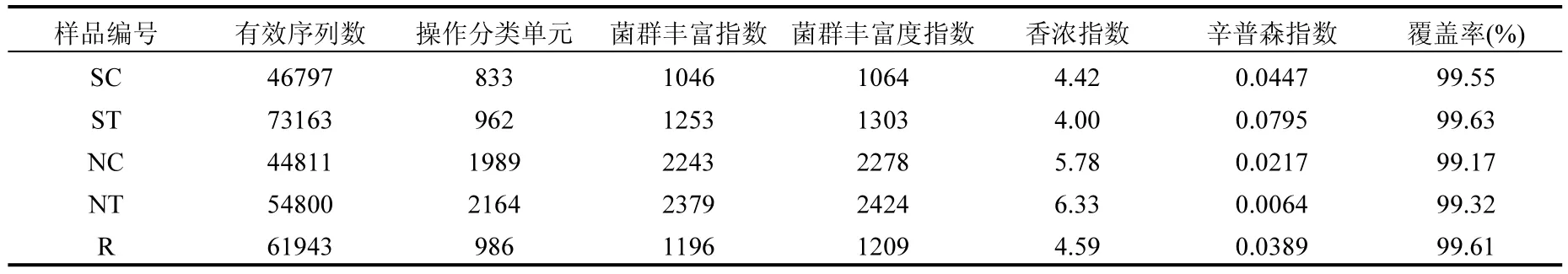
表2 樣品的多樣性指數Table 2 Alpha-diversity of the samples
2.3 菌群結構組成
對有效序列進行系譜分類,統計各個樣品中重要的細菌門類及其相對豐度,共得到了10個重要菌門,分別是變形菌門(Proteobacteria)、藍細菌門(Cyanobacteria)、擬桿菌門(Bacteroidetes)、放線菌門(Actinobacteria)、綠彎菌門(Chloroflexi)、厚壁菌門(Firmicutes)、梭桿菌門(Fusobacteria)、酸 桿 菌 門 (Acidobacteria)、 浮 霉 菌 門(Planctomycetes)和綠菌門(Chlorobi)(表3).研究結果表明,門類學水平上,實驗組和對照組環境微生物中最豐富的優勢菌門是變形菌門和藍細菌門,但2個組別水環境中藍細菌門的相對豐度明顯高于其沉積物,變形菌門的相對豐度明顯低于其沉積物.實驗組和對照組水環境微生物群落中放線菌門和浮霉菌門的相對豐度均明顯高于其沉積物,梭桿菌門和酸桿菌門的相對豐度均明顯低于比其沉積物.空心菜根系微生物群落中的變形菌門和藍細菌門占有絕對優勢(52.80%和25.77%).
篩選出每類樣品中相對豐度最高的前10種明確分類的微生物群落,并在菌屬水平上對每類樣品的菌群結構及分布進行統計分析(圖2).實驗組和對照組的水環境中最主要的優勢菌屬是聚球藻屬(Synechococcus)、hgcI_clade、芽孢桿菌屬(Bacillus)、CL500-29_marine_group、分支桿菌屬(Mycobacterium)、CL500-3和Polynucleobacter等菌屬.實驗組和對照組的沉積物中最主要的優勢菌屬是聚球藻屬、擬浮絲藻屬(Planktothricoides)、芽孢桿菌屬、鯨桿菌屬(Cetobacterium)、Dechloromonas和厭氧粘細菌屬(Anaeromyxobacter).實驗組水環境中的聚球藻屬、芽孢桿菌屬、CL500-3和分支桿菌屬的相對豐度均高于對照組,Polynucleobacter、CL500-29_marine_group和hgcI_clade的相對豐度均低于對照組,其中聚球藻屬的相對豐度明顯高于對照組,分別為42.16%和31.51%.實驗組沉積物中的聚球藻屬的相對豐度明顯高于對照組,分別為5.19%和2.16%;擬浮絲藻屬的相對豐度明顯低于對照組,分別為2.35%和11.05%; Dechloromonas的相對豐度明顯低于對照組,分別為1.95%和3.78%.空心菜根系的前10種優勢菌落是紅細菌屬(Rhodobacter)、噬氫菌屬(Hydrogenophaga)、聚球藻屬、Meganema、紅芽生菌屬(Rhodoblastus)、芽孢桿菌屬、Shinella、玫瑰彎菌屬(Roseiflexus)、球衣菌屬(Sphaerotilus)和羅爾斯通菌屬(Ralstonia),其中紅細菌屬(7.68%)、噬氫菌屬(3.73%)和聚球藻屬(3.40%)占有絕對優勢.除了擬浮絲藻屬是一類水華藍藻,以上提到的其他主要優勢菌屬都是有益菌類.綜合這些結果,發現魚菜共生模式下環境微生物的主要優勢菌屬是聚球藻屬、hgcI_clade、CL500-29_marine_group、芽孢桿菌屬和分支桿菌屬等,空心菜根系微生物群落中主要優勢菌屬是紅細菌屬、噬氫菌屬和聚球藻屬等.研究結果表明空心菜生物浮床能夠有效改善養殖環境中的微生態平衡.

表3 各樣品中優勢細菌門類及相對豐度(%)Table 3 Advantage bacterial phyla and the relative abundance of each sample (%)
2.4 氮循環細菌
根據《伯杰氏細菌分類手冊(第九版)》[19]中對硝化細菌和反硝化細菌的分類和描述,本次研究中主要發現了2類硝化細菌菌屬即亞硝化單胞菌屬(Nitrosomonas)和硝化螺菌屬(Nitrospira)和4類反硝化細菌菌屬即假單胞菌屬(Pseudomonas)、副球菌屬(Paracoccus)、芽孢桿菌屬和噬氫菌屬(表4).根據研究結果可知實驗組硝化細菌的相對豐度高于對照組,分別為0.786%和0.420%,尤其是實驗組和對照組的沉積物微生物群落中含有大量的硝化細菌,分別為0.783%和0.419%;實驗組的反硝化細菌的相對豐度也高于對照組,分別為4.973%和4.407%;空心菜根系微生物群落中含有大量的反硝化細菌(6.679%).這些結果表明沉積物中的硝化細菌的含量比水環境中硝化細菌的含量高,沉積物中的反硝化細菌的含量比水環境中反硝化細菌的含量低.研究結果還表明空心菜生物浮床可以有效地提高養殖環境中硝化細菌和反硝化細菌的含量,尤其是反硝化細菌,進而有利于保持較好的養殖環境.
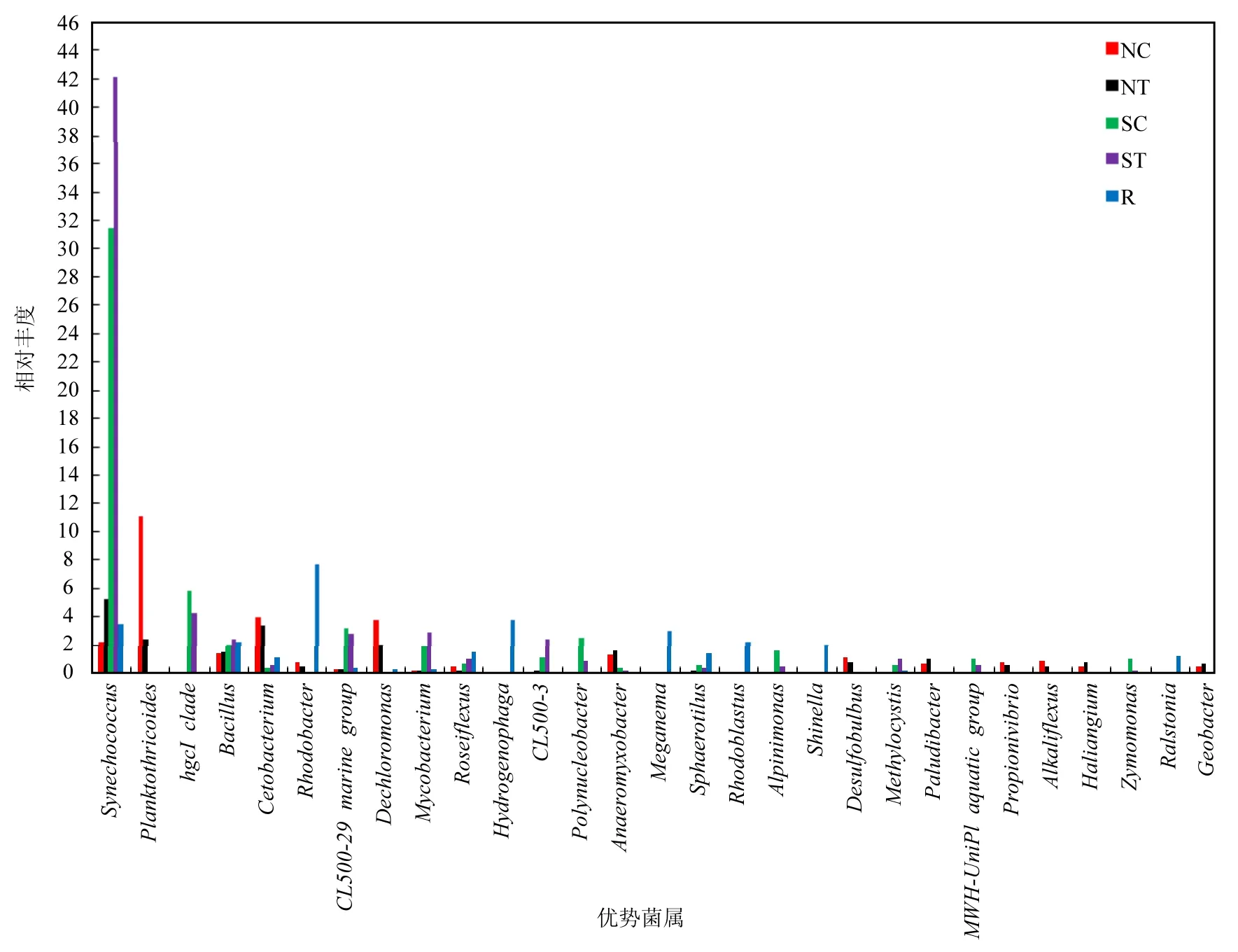
圖2 各樣品中的優勢菌屬及其相對豐度Fig.2 Relative abundance of the predominant genera in the samples
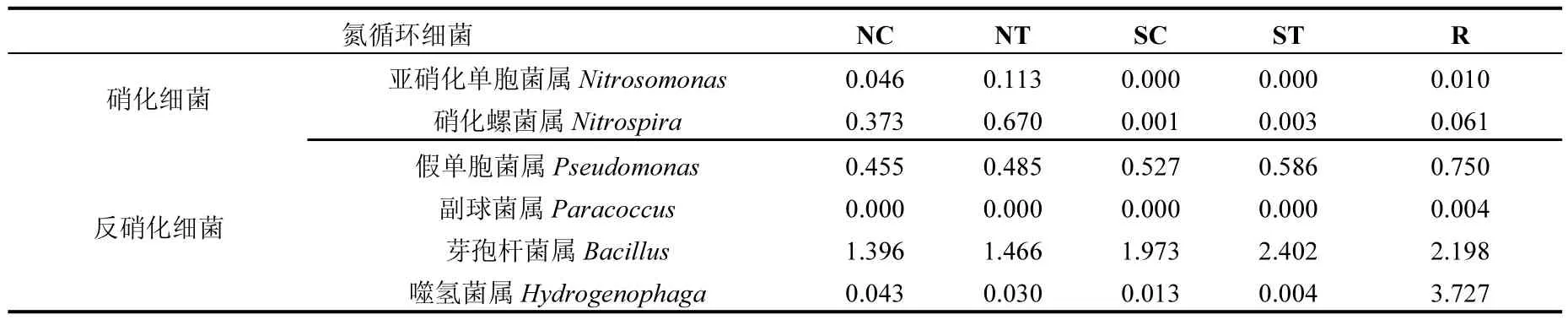
表4 氮循環細菌的組成及相對豐度(%)Table 4 The composition and relative abundance of nitrogen cycle bacteria (%)
2.5 微生物群落與環境因子之間的關系
為研究微生物群落與環境因子之間的關系,本實驗將優勢微生物群落與環境因子進行了冗余分析(RDA),如圖3.
研究結果表明空心菜根系的優勢菌落紅細菌屬、噬氫菌屬、Meganema、紅芽生菌屬、Shinella、玫瑰彎菌屬和球衣菌屬等與DO和硝酸鹽呈正相關關系,與亞硝酸鹽和TAN呈現負相關關系.紅細菌屬與TN呈負相關關系.聚球藻屬和分支桿菌屬與所有的水質因子之間的相關性都很弱.芽孢桿菌屬和羅爾斯通菌屬與DO、硝酸鹽和亞硝酸鹽呈正相關關系,與 TN和 TAN之間呈負相關關系.擬浮絲藻屬和Dechloromonas與TN和TAN呈正相關關系,與DO和硝酸鹽呈負相關關系.
hgcI_clade、CL500-29_marine_group和Polynucleobacter與亞硝酸鹽和TAN呈正相關關系,與DO和硝酸鹽呈負相關關系.
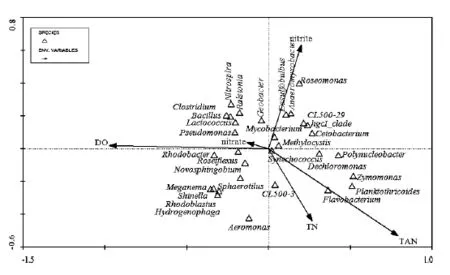
圖3 優勢菌屬和環境因子的 RDA排序圖Fig.3 RDA diagram of predominant genera and environmental factors
3 討論
眾多研究都表明生物浮床能夠有效地去除水體中的污染物.有植物系統對廢水中的總氮、BOD、SS、氨氮和重金屬等的去除能力明顯高于無植物系統[20],且生物浮床處能夠大幅降低污水中的氨氮和總磷等污染物[11-14,21].大量研究表明空心菜生物浮床能夠有效去除水體中的含氮化合物等污染物質[22-24].本研究也表明空心菜生物浮床能夠有效降低水環境中的總氮、亞硝酸鹽和總氨氮的濃度,但本研究還發現養殖過程中含氮化合物會不斷積累[25],需要及時調節水質,保證健康養殖.
已有研究發現生物浮床會降低養殖水體中的溶解氧含量.由于生物浮床覆蓋水面,阻礙了氧氣的水溶作用,并降低了藻類的光合作用,導致水體“氧源”的減少[22],同時植物根系的呼吸作用及根際微生物的硝化與反硝化作用會消耗大量氧氣[26].但也有學者研究發現當生物浮床覆蓋率較低(≤20%)時,生物浮床對水體溶解氧濃度的影響不顯著[27-28],覆蓋率為7.5%的空心菜生物浮床對池塘整體的溶氧水平無顯著影響[29].本研究的結果與黃海平[24]的研究結果一致,都發現魚菜共生系統中水體的溶解氧濃度與對照組無顯著差異,但卻有所增加,這在生產應用等方面具有重要意義.原因可能是在低覆蓋率(3.5%)條件下,空心菜生物浮床有效改善了養殖環境中的微生態平衡,這些微生物的變化最終產生了一種“綜合效應”,有效地改善了養殖環境,促使水體中溶解氧含量增加以及含氮化合物濃度降低.
本研究發現魚菜共生模式下養殖水環境中微生物具有高豐富度和低多樣性,沉積物中微生物具有高豐富度和高多樣性,而且有益微生物明顯提高.本研究中養殖環境中有益微生物主要有聚球藻屬、CL500-29_marine_group、hgcI_clade、芽孢桿菌屬和分支桿菌屬等,空心菜根系微生物菌落中還有大量的有益微生物如紅細菌屬、噬氫菌屬和聚球藻屬等,這些微生物都是有益的優勢菌屬.聚球藻屬是一類原核超微藻類,是初級生產力的主要參與者和貢獻者,在生態系統的光合作用、碳循環及食物鏈中具有重要作用[30].芽孢桿菌屬和分支桿菌屬是以多環芳烴(PAHs)為唯一碳源的微生物[31],且芽孢桿菌屬還兼具反硝化作用.菌群結構組成表明實驗組的芽孢桿菌屬的相對豐度高于對照組,RDA分析表明芽孢桿菌屬與DO、硝酸鹽和亞硝酸鹽之間呈正相關關系,與TN和 TAN之間呈負相關關系,推測芽孢桿菌屬可能通過降解水體中的碳源污染物和反硝化作用,促進養殖環境的氮循環等,并與環境因子相互影響,提高水體DO,改善養殖環境.hgcI_clade具有很強的吸收碳水化合物和富氮化合物的遺傳能力[32],并耐受較低的DO濃度[33],還能利用低溫水中的低濃度溶解有機碳,而CL500-29_marine_ group能夠有效利用多種形式的碳化合物[34].菌群結構組成表明實驗組的hgcI_clade和CL500-29_marine_group的相對豐度低于對照組, RDA分析表明hgcI_clade和CL500-29_ marine_group與亞硝酸鹽和TAN呈正相關關系,與DO和硝酸鹽呈負相關關系,推測hgcI_clade和CL500-29_marine_group通過促進碳循環和氮循環等,并與環境因子相互影響,改善養殖環境.紅細菌屬是一類高效凈化水質的光合細菌,兼具反硝化作用[35],噬氫菌屬是一類反硝化細菌.菌群結構組成表明實驗組空心菜根系微生物中含有大量的紅細菌屬和噬氫菌屬,且RDA分析表明紅細菌屬和噬氫菌屬與DO和硝酸鹽呈正相關關系,與TAN和亞硝酸鹽呈現負相關關系,而且紅細菌屬還與TN呈負相關關系,推測紅細菌屬和噬氫菌屬可能通過光合作用、硝化作用和反硝化作用等促進養殖環境的氮循環等,并與環境因子相互影響,改善養殖環境.本研究還發現空心菜生物浮床可以有效地提高養殖環境中硝化細菌和反硝化細菌的含量,尤其在植物根系存在大量的反硝化細菌.氨氮可在富氧區被硝化細菌以硝化作用氧化為硝態氮,而硝態氮則在缺氧區中被反硝化細菌以反硝化作用還原為分子態氮而進入大氣中[36].生物浮床的覆蓋區存在“富氧—厭氧微環境”區,可為微生物的活動提供有利條件[37-38],使氮循環細菌中的不同類型細菌能夠同時存在,促進水體氮循環[39].“富氧—厭氧微環境”區的同時或交替存在還可促進根區微生物的生長和繁殖,從而提高生物浮床系統對污染物的去除效率[40-41].正是在這些優勢微生物的“綜合效應”下,魚菜共生模式下水環境中溶解氧含量相對較高,含氮化合物濃度相對較低,促使養殖環境更優良,更有利于健康養殖.
本研究中魚菜共生模式下環境微生物主要是優勢菌門是變形菌門和藍細菌門、擬桿菌門、放線菌門、綠彎菌門、厚壁菌門、梭桿菌門、酸桿菌門、浮霉菌門和綠菌門,主要菌屬是聚球藻屬、芽孢桿菌屬和分支桿菌屬等,空心菜根系微生物菌落中主要菌屬是紅細菌屬、噬氫菌屬和聚球藻屬等.本研究的結果與其他研究者的結果存在一定的差異.眾多學者研究表明淡水水體中微生物主要屬于變形菌門、放線菌門、擬桿菌門、藍細菌門、疣微菌門和浮霉菌門等[42-44],優勢菌屬為嗜胞菌屬(Cytophaga)、黃桿菌屬(Flavobacterium)、伯克氏菌屬(Burkholderia)等[42],植物根際優勢菌屬以假單胞菌屬、微球菌屬(Micrococcus)、金黃桿菌屬(Chryseobacterium)和不動桿菌屬(Acinetobacter)等微生物為主[45-47].導致存在差異的主要原因是水體微生物群落結構受自然環境和水體環境因子如光照、溫度、pH值、水流、風速等條件的影響[48],水生植物根際微生物多樣性受植物類型、植物長勢、水體環境因子等多種因素的相互影響[49],另外技術進步和研究方法也會導致差異的出現[48].
4 結論
采用宏基因組學測序技術和生物信息學手段,分析了環境微生物和根系微生物群落的多樣性.研究結果表明魚菜共生模式下優勢菌門是變形菌門、藍細菌門、擬桿菌門、放線菌門、綠彎菌門和厚壁菌門等,主要菌屬是聚球藻屬、hgcI_clade、CL500-29_marine_group、芽孢桿菌屬和分支桿菌屬等;空心菜根系微生物群落中最豐富的優勢菌門是變形菌門和藍細菌門,主要菌屬是紅細菌屬、噬氫菌屬和聚球藻屬等.研究結果還表明低覆蓋率(3.5%)的空心菜生物浮床能夠增加水體的溶解氧含量,降低水體中含氮化合物的濃度和改善養殖環境中的微生態平衡,提高有益菌和氮循環細菌的含量.
[1] Cahill P L, Hurd C L, Lokman M. Keeping the water clean -Seaweed biofiltration outperforms traditional bacterial biofilms in recirculating aquaculture [J]. Aquaculture, 2010:153-159.
[2] Randall D J, Wright P A. Ammonia distribution and excretion in fish [J]. Fish Physiology & Biochemistry, 1987,3:107-120.
[3] Wright P A. Nitrogen excretion: three end products, main physiological roles [J]. Journal of Experimental Biology, 1995,198:273-281.
[4] Randall D J, Tsui T K N. Ammonia toxicity in fish [J]. Marine Pollution Bulletin, 2002,45:17-23.
[5] Thurston R V, Russo R C. Acute toxicity of ammonia to rainbow trout [J]. Transactions of the American Fisheries Society, 1983,112:696-704.
[6] 余瑞蘭,聶湘平,魏泰莉,等.分子氨和亞硝酸鹽對魚類的危害及對策 [J]. 中國水產科學, 1997,6(3):73-77.
[7] 郭增華,王秋燕.亞硝酸鹽氮對方斑東風螺毒性的研究 [J]. 海洋水產研究, 2006,27:88-92.
[8] Sun L P, Liu Y, Jin H. Nitrogen removal from polluted river by enhanced floating bed grown canna [J]. Ecological Engineering,2009,35:135-140.
[9] Lin Y F, Jing S R, Lee D Y, et al. Nutrient removal from aquaculture wastewater using a constructed wetlands system [J]. Aquaculture, 2002,209:169-184.
[10] Zhu L D, Li Z H, Ketola T. Biomass accumulations and nutrient uptake of plants cultivated on artificial floating beds in China's rural area [J]. Ecological Engineering, 2011,37:1460-1466.
[11] 宋祥甫,鄒國燕,吳偉明,等.浮床水稻對富營養化水體中氮、磷的去除效果及規律研究 [J]. 環境科學學報, 1998,18: 489-495.
[12] 劉淑媛,任久長,由文輝.利用人工基質無土栽培經濟植物凈化富營養化水體的研究 [J]. 北京大學學報(自然科學版),1999,35:518-522.
[13] 司友斌,包軍杰,曹德菊,等.香根草對富營養化水體凈化效果研究 [J]. 應用生態學報, 2003,14:277-279.
[14] 周小平,王建國,薛利紅,等.浮床植物系統對富營養化水體中氮、磷凈化特征的初步研究 [J]. 應用生態學報, 2005,16:2199-2203.
[15] HJ 636-2012 水質-總氮的測定-堿性過硫酸鉀消解紫外分光光度法 [S].
[16] HJ 535-2009 水質-氨氮的測定-納氏試劑分光光度法 [S].
[17] GB 7493-87 水質-亞硝酸鹽氮的測定-分光光度法 [S].
[18] GB 7480-87 水質-硝酸鹽氮的測定-酚二磺酸分光光度法[S].
[19] Holt J G, Krieg N R, Sneath P H A, et al. Bergey's Manual of Determinative Bacteriology,9thEdition [M]. Baltimore,Lippincott Williams & Wilkins, 1994,10-786.
[20] Dunbabin J S, Pokorny J, Bowmer K H. Rhizosphere oxygenation by Typha domingensis Pers. in miniature artificial wetland filters used for metal removal from wastewaters [J]. Aquatic Botany, 1988,29:303-317.
[21] Amanda M N, William J M. Tropical treatment wetlands dominated by free-floating macrophytes for water quality improvement in Costa Rica [J]. Ecological Engineering, 2006: 246-257.
[22] 林連升.精養魚池水質生物凈化初步研究 [D]. 南京:南京農業大學, 2006.
[23] Li M, Wu Y J, Yu Z L, et a1. Nitrogen removal from eutrophic water by floating-bed-grown water spinach (Ipomoea aquatica Forsk.) with ion implantation [J]. Water Research, 2007,41:3152—3158.
[24] 常會慶,寇太記,喬鮮花,等.幾種植物去除污染水體中養分效果研究 [J]. 水土保持通報, 2009,5:118-122.
[25] 李 谷,吳振斌,侯燕松,等.養殖水體氮的生物轉化及其相關微生物研究進展 [J]. 中國生態農業學報, 2006,14(1):11-15.
[26] 雷衍之.養殖水環境化學 [M]. 北京:中國農業出版社, 2004: 119-112.
[27] 陳家長,孟順龍,胡庚東,等.空心菜浮床栽培對集約化養殖魚塘水質的影響 [J]. 生態與農村環境學報, 2010,26(2):155-159.
[28] 宋 超,陳家長,戈賢平,等.浮床栽培空心菜對羅非魚養殖池塘水體中氮和磷的控制 [J]. 中國農學通報, 2011,27(23):70-75.
[29] 黃海平.水蕹菜浮床在精養魚池中的應用效果研究 [D]. 武漢:華中農業大學, 2012.
[30] 李勝男,周 建,魏利軍,等.淡水超微型浮游植物多樣性及其研究方法 [J]. 生態學雜志, 2015,34(4):1174-1182.
[31] 霍煒潔,肖晶晶,黃亞麗,等.微生物技術修復水污染的研究進展[J]. 生物技術通報, 2008,4:94-98.
[32] Ghylin T W, Garcia S L, Moya F, et al. Comparative single-cell genomics reveals potential ecological niches for the freshwater acⅠ Actinobacteria lineage [J]. Ⅰsme Journal, 2014,8(12):2503-2516.
[33] Liu J W, Fu B B, Yang H M, et al. Phylogenetic shifts of bacterioplankton community composition along the Pearl Estuary: the potential impact of hypoxia and nutrients [J]. Frontiers in Microbiology, 2015,6:64.
[34] Lindh M V, Lefebure R, Degerman R, et al. Consequences of increased terrestrial dissolved organic matter and temperature on bacterioplankton community composition during a Baltic Sea mesocosm experiment [J]. Ambio, 2015,44Suppl 3(3):402-12.
[35] 鄭耀通,胡開輝,高樹芳,等.高效凈化水產養殖水域紫色非硫光合細菌的分離和篩選 [J]. 福建農業大學學報, 1998,27(3):257-260.
[36] Reddy K R, Patrick W H, Lindau C W. Nitrificationdenitrification at the plant root-sediment interfaces in wetlands[J]. Limnology and Oceanography, 1989,34(6):1004-1013.
[37] Stottmeister U, Wie?ner A, Kuschk P, et al. Effects of plants and microorganisms in constructed wetlands for wastewater treatment[J]. Biotechnology Advances, 2003,22:93-117.
[38] 胡綿好.水生經濟植物浮床技術改善富營養化水體水質的研究[D]. 上海:上海交通大學, 2008.
[39] 吳 偉,胡庚東,金蘭先,等.浮床植物對池塘水體微生物的動態影響 [J]. 中國環境科學, 2008,28(9):791-795.
[40] Huetta D O, Morrisb S G, Smitha G, et al. Nitrogen and phosphorus removal from plant nursery runoff in vegetated and unvegetated subsurface flow wetlands [J]. Water Research, 2005,39:3259-3272.
[41] Keffala C, Ghrabi A. Nitrogen and bacterial removal in constructed wetlands treating domestic waste water [J]. Desalination, 2005,185(1-3):383-389.
[42] Jürg B L, Helmut B, Christopher T R. Progress in the Ecological Genetics and Biodiversity of Freshwater Bacteria [J]. Bioscience,2008,58(2):103-113.
[43] Eiler A, Bertilsson S. Composition of freshwater bacteria communities associated with cyanobacterial blooms in four Swedish lakes [J]. Environmental Microbiology, 2004,16:1228-1243.
[44] Tamaki H, Sekiguchi Y, Hanada S, et al. Comparative analysis of bacterial diversity in freshwater sediment of a shallow eutrophic lake by molecular and improved cultivation-based techniques [J]. Applied & Environmental Microbiology, 2005,71:2162-2169.
[45] 詹發萃,鄧家齊,夏宜琤,等.鳳眼蓮根區異養細菌的群落特征與異養活性的研究 [J]. 水生生物學報, 1993,17(2):150-156.
[46] 張甲耀,林清華,夏盛林,等.不同植物構成的潛流型人工濕地處理系統的凈化能力及其異養細菌數量的研究 [J]. 環境工程,1998,16(3):17-20.
[47] 陸開宏,胡智勇,梁晶晶,等.富營養水體中2種水生植物的根際微生物群落特征 [J]. 中國環境科學, 2010,30(11):1508-1515.
[48] 王紹祥,楊洲祥,孫 真,等.高通量測序技術在水環境微生物群落多樣性中的應用 [J]. 化學通報, 2014,77(3):196-303.
[49] Wasaki J, Rothe A, Kania A. Root exudation, Phosphorus acquisition and microbial diversity in the rhizosphere of white lupine as affected by phosphorus supply and atmospheric carbon dioxide concentration [J]. Journal of Environmental Quality, 2005,34(6):2157-2167.
Influence on water quality and microbial diversity in fish pond by Ipomoea aquatica floating-bed.
LI Jian-zhu, HOU Jie, ZHANG Peng-fei, LIU Yao-quan, XIA Run-hong, MA Xu-fa*(Huazhong Agricultural University, Wuhan 430070,China). China Environmental Science, 2016,36(10):3071~3080
In order to investigate the influence on water quality and microbial diversity in intensive fish pond by Ipomoea aquatica floating-bed, and the composition and diversity of microbial communities in the samples were researched based on high throughput sequencing of 16S rRNA gene. The results revealed that the dominant bacteria were phyla Proteobacteria, Cyanobacteria, Bacteroidetes, Actinobacteria, Chloroflexi and Firmicutes, the dominant bacteria were genera Synechococcus, hgcI_clade, CL500-29_marine_group, Bacillus and Mycobacterium in the Aquaponics System,and the most dominant bacteria of rhizosphere microorganisms were phyla Proteobacteria and Cyanobacteria, the dominant bacteria were genera Rhodobacter, Hydrogenophaga and Synechococcus. Moreover, the results also showed that low coverage (3.5%) of Ipomoea aquatica floating-bed increased the dissolved oxygen content and reduced the concentration of nitrogen compounds, and improved micro-ecological balance and increased the content of the beneficial bacteria and nitrogen cycle bacteria in aquaculture water body.
Ipomoea aquatica floating-bed;environmental microbial diversity;water quality;16S rDNA
X171.1,S917.1
A
1000-6923(2016)10-3071-10
李建柱(1990-),男,湖北孝感人,華中農業大學碩士研究生,主要研究方向為養殖水環境生物監測與調控.
2016-02-18
國家科技支撐計劃課題(2012BAD25B05)和湖北省自然科學基金(2013CFA095)資助
* 責任作者, 副教授, xufama@mail.hzau.edu.cn
猜你喜歡
中老年保健(2021年12期)2021-08-24 03:30:40
中國傳媒大學學報(自然科學版)(2021年1期)2021-06-09 08:43:00
中國生殖健康(2020年6期)2020-02-01 06:28:50
新世紀智能(英語備考)(2019年12期)2020-01-13 06:07:18
瘋狂英語·新策略(2019年9期)2019-10-17 01:51:34
中國生殖健康(2019年11期)2019-01-07 01:28:02
中國生殖健康(2018年6期)2018-11-06 07:09:28
濰坊學院學報(2017年2期)2017-04-20 08:44:31
中國環境監察(2016年5期)2016-10-24 05:25:52
中國商論(2016年33期)2016-03-01 01:59:38
