秦嶺箭竹生物量分配規(guī)律
2017-01-18 07:27:58朱慧男何強利范曉東
綠色科技 2016年18期
朱慧男, 何強利, 范曉東
(陜西太白山國家級自然保護區(qū)管理局, 陜西 楊凌 712100)
?
秦嶺箭竹生物量分配規(guī)律
朱慧男, 何強利, 范曉東
(陜西太白山國家級自然保護區(qū)管理局, 陜西 楊凌 712100)
為探明秦嶺箭竹生物量分布規(guī)律,以太白山保護區(qū)厚畛子保護站管區(qū)后溝、云霧溝地區(qū)為研究區(qū)域,對秦嶺箭竹進行了調查,探討了秦嶺箭竹生物量與海拔高度之間的關系,并分析了秦嶺箭竹單株生物量在地上和地下的分配格局,結果表明:秦嶺箭竹個體水平上在同一海拔高度不同個體間呈現(xiàn)數(shù)值變異。隨著海拔的升高,同一海拔內個體差異降低。不同海拔秦嶺箭竹生物量差異顯著,隨著海拔的升高,秦嶺箭竹個體生物量降低;秦嶺箭竹在個體水平,地上生物量、地下生物量和總生物量的變異系數(shù)均大于1,表現(xiàn)出較大的個體變異,地下生物量平均值為4.01小于地上生物量6.91。個體水平上的R/S值在0.2~1,8之間,平均值為0.57,大部分小于1,0.2~0.7之間分布最多;比較選出幾個關系模型,建立的模型效果良好。
秦嶺箭竹;生物量;規(guī)律
1 引言
生物量是指單位面積上所有生物有機體的干重。森林生物量是森林生態(tài)系統(tǒng)結構和功能的最基本特征之一,生態(tài)系統(tǒng)的能量和營養(yǎng)循環(huán)的研究首先依賴于生物量的數(shù)據(jù),在研究森林生長和森林生態(tài)系統(tǒng)動態(tài)變化過程中,生物量的研究和測定顯得極為重要,世界各國廣泛開展了對森林生物量的監(jiān)測[1,2]。國外就已開始了不同類型森林生物量的研究[3]。國內目前在個體、種群、群落、生態(tài)系統(tǒng)、區(qū)域、全國尺度、生物圈等多尺度上已開展了相應的生物量研究[4~10]。研究主要集中于油松[11~14]落葉松[15~18]、馬尾松[19~20]、山楊、櫟等樹種的生物量分配以及模型構建方面,關于秦嶺林區(qū)竹類的研究主要集中在主食竹種的資源調查、分類與分布、生物生態(tài)學特性等方面,對秦嶺箭竹的單株生物量研究較少。
秦嶺箭竹(Fargesiaqinlingensis)屬禾本科竹亞科箭竹屬,在太白山自然保護區(qū)內,秦嶺箭竹主要分布于海拔1500~2900 m的針闊混交林林內,常為單一竹種在林冠下形成優(yōu)勢層片。對于高山地區(qū)的水土保持和環(huán)境防護具有重要的生態(tài)意義[21~26]。但由于人為打筍、采藥等持續(xù)的干擾破壞及竹子開花、森林火災等自然災害的影響,天然秦嶺箭竹林正面臨著面積減少、生長衰退等問題的困擾。研究太白山保護區(qū)的秦嶺箭竹單株生物量大小及其在地上和地下的分配格局,不僅對探索秦嶺箭竹生物量與大熊貓棲息地選擇關系具有重要意義,而且對評價秦嶺大熊貓森林群落質量和對大熊貓棲息地森林群落進行科學管理和經(jīng)營具有重要作用,對于進一步研究該區(qū)的碳儲量和碳循環(huán)都有著非常重要的價值。
本研究以太白山保護區(qū)厚畛子保護站管區(qū)后溝、云霧溝地區(qū)為研究區(qū)域,對秦嶺箭竹進行調查分析,探明秦嶺箭竹生物量與海拔高度之間的關系,揭示秦嶺箭竹分布規(guī)律,探討秦嶺箭竹單株生物量在地上和地下的分配格局,為確定該地區(qū)秦嶺箭竹的生物量變化規(guī)律提供參考,進而為研究森林生產(chǎn)力和碳儲量及碳匯功能的評價提供定量數(shù)據(jù)基礎,為森林經(jīng)營撫育管理提供一定的理論依據(jù)。
2 材料與方法
2.1 研究區(qū)概況
研究區(qū)域為太白山保護區(qū)厚畛子保護站后溝-云霧溝區(qū)域,地理位置為東經(jīng)107°22 ′25″~107°51′30″和北緯33°49′30″~34°05′35 ″之間,海拔1598~2480 m;該區(qū)氣候為暖溫帶大陸性氣候,年均氣溫7.6 ℃,年均降水量751.8 mm,無霜期158 d,最高氣溫32.8 ℃,最低氣溫-25.5 ℃。最冷月為1月,平均氣溫為-5.0 ℃,最熱月為7月,平均氣溫為19.1 ℃。2002~2007年間,區(qū)域內秦嶺箭竹相繼開花死亡,此后監(jiān)測結果顯示該區(qū)域內大熊貓分布急劇減少,區(qū)域內秦嶺箭竹現(xiàn)處于自然更新階段。各樣地基本情況如下表:
2.2 研究方法
2.2.1 樣地調查
2014年3月~2014年5月,采用樣線調查和典型樣地調查法,以太白山自然保護區(qū)管理局的巡護樣線16A為調查樣線,沿樣線海拔每上升100~200 m設20 m×20 m的樣地,調查內容包括GPS定位點、海拔、坡度、坡向、坡位、林冠層樹種組成、植被類型、林冠郁閉度和竹子總體蓋度。在每個樣地內對角和中心位置布設2 m×2 m的小樣方,調查竹子年齡、高度、基徑、數(shù)量、密度、蓋度等。
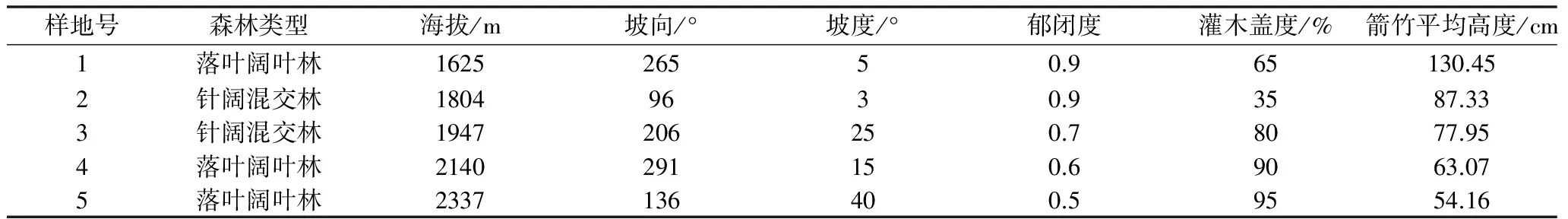
表1 各樣地基本情況
2.2.2 生物量測定
竹子生物量測定采用收獲法,在調查樣地內隨機選擇10棵秦嶺箭竹,按器官分別稱取其鮮重并取200g鮮樣,樣品帶回室內在105 ℃條件下烘干至恒重,計算各器官含水率,根據(jù)含水率計算生物量。
3 結果與分析
3.1 不同海拔單株生物量變化
從圖1可以看出,在秦嶺箭竹個體水平上,同一海拔高度不同個體間呈現(xiàn)數(shù)值變異。隨著海拔的升高,同一海拔內個體差異降低。不同海拔秦嶺箭竹生物量差異顯著,隨著海拔的升高,秦嶺箭竹個體生物量降低。
由于低海拔地區(qū)光照、溫度、土壤等環(huán)境條件優(yōu)于高海拔地區(qū),秦嶺箭竹的生長較好,低海拔內秦嶺箭竹個體生物量積累遠高于高海拔地區(qū)。在同一海拔高度內,外界條件的小范圍變化引起秦嶺箭竹個體生長差異,低海拔地區(qū)秦嶺箭竹生長對環(huán)境變化更敏感,而高海拔地區(qū),環(huán)境條件較差,個體間差異不明顯。
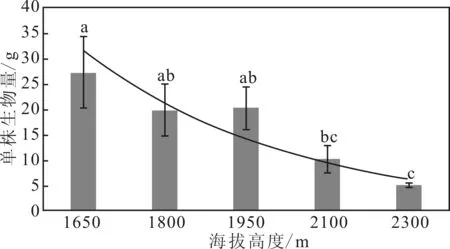
圖1 不同海拔單株生物量
3.2 秦嶺箭竹個體水平上生物量分配情況
由表2可以看出,秦嶺箭竹在個體水平,地上生物量、地下生物量和總生物量的變異系數(shù)均大于1,表現(xiàn)出較大的個體變異,地下生物量平均值為4.01 g小于地上生物量6.91 g。
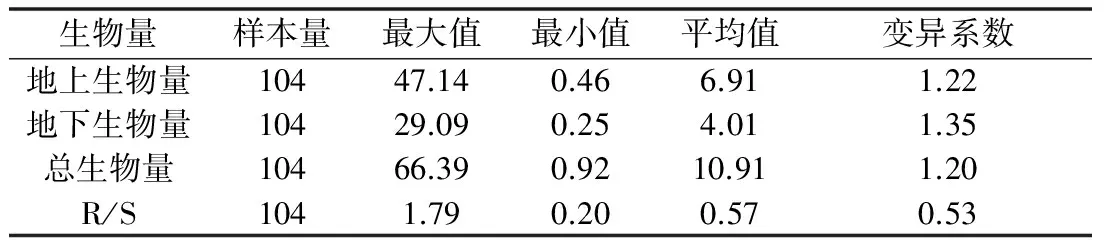
表2 秦嶺箭竹個體水平上生物量分配
結合圖2可知,個體水平上的R/S值在0.2~1.8之間,平均值為0.57,大部分小于1,0.2~0.7之間分布最多。
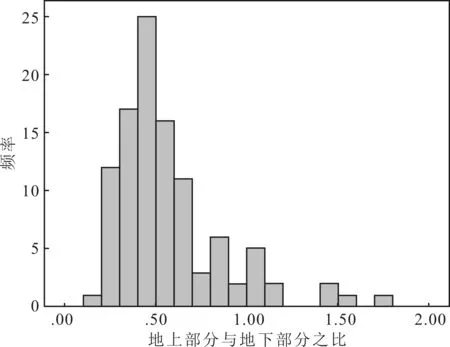
圖2 個體水平上生物量地上與地下部分之比(R/S)頻數(shù)分布
3.3 地上和地下生物量的相關生長關系
決定系數(shù)R2的大小決定了相關的密切程度。當R2越接近1時,表示相關的方程式參考價值越高;相反,越接近0時,表示參考價值越低。F值表示回歸模型的方差與殘差的比值(殘差就是總方差減去回歸模型的方差),理論上來說F值越大,通過檢驗的可能性就越大。可以想象F越大,表示殘差越小,模擬的精度越高。P值是統(tǒng)計學上結果真實程度,p值越大,越不能認為樣本中變量的關聯(lián)是總體中各變量關聯(lián)的可靠指標。在許多研究領域,0.05的p值通常被認為是可接受錯誤的邊界水平。
對地上生物量與地下生物量關系進行常見的關系擬合,可以看出R2最大的是邏輯斯蒂模型,值為0.6992,最小的是指數(shù)函數(shù)模型,值為0.4328,剩余模型R2 均大于0.6,說明模型具有參考價值。P值均為0.0001,認為樣本中變量的關聯(lián)是總體中各變量關聯(lián)的可靠指標。F值均較大,通過檢驗的可能性就較大,幾個關系模型效果良好。在野外調查中,為了方便,可以根據(jù)實際情況,測定地上生物量或者地下生物量推算整體生物量,一定程度上節(jié)約了勞動時間和勞動量。

表3 地上和地下生物量的相關生長關系模型
注:X1地上生物量X2地下生物量
4 結語
由于水災等自然因素,部分預設樣地未發(fā)現(xiàn)秦嶺箭竹分布,后重新布設5個樣地,共調查樣地5個。調查結果表明,秦嶺箭竹個體水平上在同一海拔高度不同個體間呈現(xiàn)數(shù)值變異。隨著海拔的升高,同一海拔內個體差異降低。不同海拔秦嶺箭竹生物量差異顯著,隨著海拔的升高,秦嶺箭竹個體生物量降低。由于低海拔地區(qū)光照、溫度、土壤等環(huán)境條件優(yōu)于高海拔地區(qū),秦嶺箭竹的生長較好,低海拔內秦嶺箭竹個體生物量積累遠高于高海拔地區(qū)。在同一海拔高度內,外界條件的小范圍變化引起秦嶺箭竹個體生長差異,低海拔地區(qū)秦嶺箭竹生長對環(huán)境變化更敏感,秦嶺箭竹個體生長差異很大,而高海拔地區(qū),環(huán)境條件較差,所有個體間均生長不好,個體間差異不明顯。秦嶺箭竹是秦嶺大熊貓食物的主要來源,其空間分布和數(shù)量多少在很大程度上決定了大熊貓的生境分布狀況,在海拔1900~2600m秦嶺箭竹生長最為茂盛的地段,為有效保護大熊貓及其生境,建議保護區(qū)在管護和森林群落經(jīng)營管理方面,應盡量避免在秦嶺箭竹適宜生長的地段海拔1900~2600m范圍內林冠郁閉度0.6以下的落葉闊葉和針闊混交林中進行人為生產(chǎn)及其他干擾活動。
秦嶺箭竹在個體水平,地上生物量、地下生物量和總生物量的變異系數(shù)均大于1,表現(xiàn)出較大的個體變異,地下生物量平均值為4.01g小于地上生物量6.91g。個體水平上的R/S值在0.2~1,8之間,平均值為0.57,大部分小于1,0.2~0.7之間分布最多。
通過數(shù)據(jù)分析,比較選出幾個關系模型,建立的模型效果良好。在野外調查中,為了方便,可以根據(jù)實際情況,測定地上生物量或者地下生物量推算整體生物量,一定程度上節(jié)約了勞動時間和勞動量。
本文僅對太白山自然保護區(qū)后溝—云霧溝區(qū)域秦嶺箭竹單株生物量進行了研究,不能代表太白山南坡秦嶺箭竹生物量分布規(guī)律,應繼續(xù)擴大研究區(qū)域,探明太白山南坡秦嶺箭竹生物量以及其分布規(guī)律。
[1]羅 輯,楊 忠,楊清偉. 貢嘎山森林生物量和生產(chǎn)力的研究[J]. 植物生態(tài)學報,2000(2):191~196.
[2]劉志剛,馬欽彥,潘向麗. 興安落葉松天然林生物量及生產(chǎn)力的研究[J]. 植物生態(tài)學報,1994(4):328~337.
[3]Leith H, Whittaker R H. Primary Productivity of the Biosphere[M].New York: Springer Verlag. 1975.
[4]馮宗煒,陳楚瑩,張家武,等. 湖南會同地區(qū)馬尾松林生物量的測定[J]. 林業(yè)科學,1982(2):127~134.
[5]馬欽彥. 中國油松生物量的研究[J]. 北京林業(yè)大學學報,1989(4):1~10.
[6]Gallis C. Probabilistic assessment of forest biomass storage times and its effect on cost: A beech biomass case study. Forest Products Journal, 2003 ,53 (10): 44~54.
[7]Fang J Y, Chen A P, Zhao S Q,et al. Estimating biomass carbon china 's forests: Supplementary notes on report published in science(291 : 2320-2322) by Fang et al[J]. Acta Phytoecologica Sinca, 2002, 26(2): 243 ~249.
[8]吳 剛,馮宗煒. 中國油松林群落特征及生物量的研究[J]. 生態(tài)學報,1994(4):415~422.
[9]方精云,劉國華,徐嵩齡. 我國森林植被的生物量和凈生產(chǎn)量[J]. 生態(tài)學報,1996(5):497~508.
[10]劉玉萃,吳明作,郭宗民,等. 內鄉(xiāng)寶天曼自然保護區(qū)銳齒櫟林生物量和凈生產(chǎn)力研究[J]. 生態(tài)學報,2001(9):1450~1456.
[11]秦仲燾,李志堅. 淺析天津市油松生物量測定方法[J]. 天津農(nóng)林科技,2013(1):1~2.
[12]王 寧,王百田,王瑞君,等. 晉西山楊和油松生物量分配格局及異速生長模型研究[J]. 水土保持通報,2013(2):151~155+159.
[13]王 明. 森林油松生物量測定方法[J]. 農(nóng)民致富之友,2013(13):142.
[14]張志明,常帥,張亞珍. 不同類型油松林生物量碳密度的研究[J]. 山西林業(yè)科技,2012(3):11~13+24.
[15]郭金堂,姜 鵬,郭凱,谷建才. 針闊混交林華北落葉松生物量模型及碳儲量研究[J]. 西北林學院學報,2016(2):45~48.
[16]宋相祿. 不同營林措施對日本落葉松生物量的影響[J]. 林業(yè)科技,2003(2):4~6.
[17]閆濤,朱教君,楊凱,等. 遼東山區(qū)落葉松人工林地上生物量和養(yǎng)分元素分配格局[J]. 應用生態(tài)學報,2014(10):2772~2778.
[18]符利勇,唐守正,張會儒,等. 東北地區(qū)兩個主要樹種地上生物量通用方程構建[J]. 生態(tài)學報,2015(1):150~157.
[19]符利勇,曾偉生,唐守正. 利用混合模型分析地域對國內馬尾松生物量的影響[J]. 生態(tài)學報,2011(19):5797~5808.
[20]樊后保,李燕燕,蘇兵強,等. 馬尾松-闊葉樹混交異齡林生物量與生產(chǎn)力分配格局[J]. 生態(tài)學報,2006(8):2463~2473.
[21]王 瑞,周建云,曹 慶,等. 不同生境秦嶺箭竹人工種群的生長特性研究[J]. 西北農(nóng)林科技大學學報(自然科學版),2011(5):110~118.
[22]杜焰玲,張 瑩,董國華,等. 秦嶺山地秦嶺箭竹群落物種多樣性研究[J]. 西北農(nóng)林科技大學學報(自然科學版),2013(10):59~66.
[23]吳高潮,李景俠,張國楨,等. 秦嶺箭竹種群生物量研究[J]. 西北林學院學報,2014(1):46~50.
[24]馬青青,劉建軍,余 鴿,等. 佛坪國家級自然保護區(qū)秦嶺箭竹克隆結構的SSR分析[J]. 生態(tài)學報,2016(20):1~10.
[25]王小紅,周建云,張文輝,等. 大熊貓棲息地秦嶺箭竹無性系構件形態(tài)研究[J]. 西北農(nóng)業(yè)學報,2010(11):93~99.
[26]任 毅,劉明時,等.太白山自然保護區(qū)生物多樣性研究與管理[M].北京:中國林業(yè)出版社,2006.
2016-08-10
秦嶺大熊貓及其棲息地保護網(wǎng)絡巡護員能力建設小額基金項目
朱慧男(1989—),男,助理工程師,主要從事森林保護與森林植物群落研究工作。
S795
A
1674-9944(2016)18-0016-03
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
中學生數(shù)理化·七年級數(shù)學人教版(2020年10期)2020-11-26 08:24:50
數(shù)學物理學報(2020年2期)2020-06-02 11:29:24
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
汽車工程學報(2017年2期)2017-07-05 08:13:02
光學精密工程(2016年6期)2016-11-07 09:07:19
