斜帶石斑魚種苗轉餌模式研究
2016-12-20 03:47:15孫坤剛
水產科學 2016年2期
關鍵詞:生物
周 暉,陳 剛,孫坤剛
( 1.廣東海洋大學 水產學院,廣東 湛江 524025;2.廣東省普通高校南海水產經濟動物增養殖重點實驗室,廣東 湛江 524025 )
斜帶石斑魚種苗轉餌模式研究
周 暉1,2,陳 剛1,2,孫坤剛1
( 1.廣東海洋大學 水產學院,廣東 湛江 524025;2.廣東省普通高校南海水產經濟動物增養殖重點實驗室,廣東 湛江 524025 )
對36日齡平均體質量(0.21±0.01) g的斜帶石斑魚種苗的轉餌模式進行研究。設計5種轉餌模式:轉餌1組前10 d投喂生物餌料,然后轉投配合飼料;轉餌2、轉餌3和 轉餌4組前10 d進行聯合投喂(轉餌2:25%配合飼料+75%生物餌料;轉餌3:50%配合飼料 +50%生物餌料;轉餌4:75%配合飼料 +25%生物餌料),然后轉投配合飼料;轉餌5組直接投喂配合飼料;轉餌試驗至種苗60日齡結束。試驗結果表明,由于魚體長度變異系數顯著高于其他各組(P<0.05),轉餌4組試驗魚死亡率最高,而其他各組之間差異不顯著(P>0.05);試驗結束時,轉餌1組試驗魚體質量和質量增加率最低,顯著低于其他各組(P<0.05),而轉餌2組又顯著低于轉餌4和轉餌5組(P<0.05);轉餌1組種苗36—45日齡的特定生長率顯著低于其他各組(P<0.05),而46—60日齡的特定生長率顯著高于其他各組(P<0.05)。轉餌試驗結束后,各組種苗全魚水分和粗蛋白含量差異不顯著(P>0.05),轉餌5組種苗全魚粗脂肪含量顯著低于其他各組(P<0.05)。綜合比較各項指標,轉餌3的模式種苗質量增加率較高而死亡率較低,是本研究中效果較好的轉餌模式。
斜帶石斑魚;聯合投喂;生物餌料;配合飼料
斜帶石斑魚(Epinpheluscoioides)是我國南方沿海養殖的主要名貴海水魚之一,生長快、適應性強、肉質鮮美、營養豐富,深受消費者歡迎。雖然斜帶石斑魚人工繁育和苗種培育技術已有研究報道[1-4],但其種苗從開口至變態完成均需要使用生物餌料喂養,育苗成本較高。Person等[5]曾估算,在齡舌齒鱸(Dicenntrarchuslabrax)種苗培育的前3個月,生物餌料占育苗總成本的50%;而在種苗培育的前45 d,生物餌料占培育成本的79%。Rosenlund等[6]研究發現,由于生物餌料含水率約為90%,其營養含量較低;海水魚種苗對生物餌料的攝食率會隨其生長而逐漸降低,若未在適當時機進行轉餌,則難以獲得較高的生長率和存活率。與生物餌料相比,配合飼料具有性價比高、使用方便、營養均衡、成分可人為調控等優點[7],在海水魚類育苗過程中往往會選擇一個合適的時期投喂配合飼料來逐漸替代生物餌料,以降低育苗成本并減少生物餌料可能帶來的病害風險。已有很多研究者對各種魚類種苗的轉餌模式開展了研究,并且在配合飼料的制備、使用和轉餌模式等方面取得了一定的進展,促進了這些魚類育苗技術的進步[5-15],但有關斜帶石斑魚種苗轉餌模式的研究尚未見報道。本試驗通過5種模式對36日齡斜帶石斑魚種苗進行轉餌,測定了各種轉餌模式下試驗魚的死亡率、特定生長率、質量增加率、魚體長度變異系數和魚體成分等指標,分析了各種轉餌模式的優缺點。
1 材料與方法
1.1 斜帶石斑魚種苗和餌料
斜帶石斑魚種苗在室外水泥池中培育,3-10日齡投喂室外水泥池培養的輪蟲,11-16日齡投喂室外水泥池培養的橈足類幼體,17-33日齡投喂人工孵化后以強化劑強化4 h或12 h的鹵蟲無節幼體,投喂至飽食。育苗池水溫27~29 ℃,鹽度29~31,pH 8.0~8.4,溶氧>6 mg/L。種苗培育至33日齡開始進行分苗處理,篩選規格一致的種苗移至試驗水槽中暫養2 d,36日齡試驗種苗平均全長(2.23±0.04) cm,體長(1.72±0.04) cm,濕質量(0.21±0.01) g。
生物餌料:經海帥蝦片(湛江市海神生物科技發展有限公司)強化10 d以上的鮮活大鹵蟲(Artemiasp.),體長0.9~1.2 mm。
配合飼料:日清海水魚配合飼料(日清丸紅飼料株式會社-知多工場)。36—45日齡投喂C1(0.6~0.9 mm),46—52日齡投喂S1(1.0 mm),53—60日齡投喂S2(1.4 mm)。
1.2 轉餌試驗
轉餌試驗在流水式養殖系統中進行,養殖試驗過程均采用飽食投喂,轉餌方式參考文獻[15],轉餌1組種苗在前10 d投喂生物餌料,46日齡后投喂配合飼料;轉餌2組~轉餌4組種苗在前10 d進行聯合投喂(轉餌2:25%配合飼料+75%生物餌料;轉餌3:50%配合飼料+50%生物餌料;轉餌4:75%配合飼料+25%生物餌料),46日齡后投喂配合飼料;轉餌5組種苗自33日齡起直接投喂配合飼料,轉餌試驗至種苗60日齡結束。每個轉餌組有3個70 cm×60 cm×50 cm水族箱,每箱試驗開始時投放種苗200尾。每日上午和下午投餌后及時清污,發現死魚及時清理。流水式養殖系統配有專門的蓄水池,天然海水經雙層沙濾和過濾袋過濾后進入蓄水池。試驗期間每個水族箱的水量約150 L,日交換量為100%~120%。每桶放2個氣石,每日持續充氣,自然光照。試驗期間水溫28~30 ℃,鹽度29~31,pH 7.9~8.2,溶解氧>6 mg/L。
1.3 采樣與分析
試驗開始時隨機取10尾種苗,取其均值作為初始體質量。45日齡和60日齡各采樣一次,每箱隨機取10尾魚測定體質量。餌料和魚體常規生化分析樣品均在105 ℃烘干至恒定質量,粗蛋白采用Foss kjeltecTM定氮儀測定,粗脂肪采用索氏提取法測定[16]。生物餌料和配合飼料脂肪酸分析參考文獻[17]的方法。
1.4 參數計算
死亡率/% =Nd/Ni×100%
特定生長率/%·d-1=(lnmt-lnm0)/t×100%
質量增加率/%=(mt-m0)/m0×100%
魚體長度變異系數按Folkvord[18]的方法計算:
變異系數/% =L/mt×100%
式中,Ni:試驗魚初始數量(尾),Nd:試驗魚死亡數量(尾),m0:初始魚體平均質量(g),mt:試驗t日后魚體平均質量(g),t:試驗天數(d),L:試驗結束時平均魚體長(cm)。
1.5 數據統計分析
試驗結果用SPSS 17.0 進行單因素方差分析及Duncan多重比較,以P<0.05為顯著差異。
2 結果與分析
2.1 生物餌料和配合飼料粗蛋白和粗脂肪含量比較
配合飼料的粗蛋白和粗脂肪含量均顯著高于生物餌料(P<0.05)(表1)。

表1 生物餌料與配合飼料粗蛋白、粗脂肪含量比較(干質量)
注:表中數據為5次測量結果的平均值±標準差,同一行數據上標有相同字母表示差異不顯著(P>0.05),上標無相同字母則表示差異顯著(P<0.05).表2同.
配合飼料的飽和脂肪酸、ω-3多不飽和脂肪酸、二十碳五烯酸、二十二碳六烯酸含量顯著高于生物餌料(P<0.05),單不飽和脂肪酸和ω-6多不飽和脂肪酸含量顯著低于生物餌料(P<0.05),多不飽和脂肪酸含量則無顯著差異(P>0.05)(表2)。
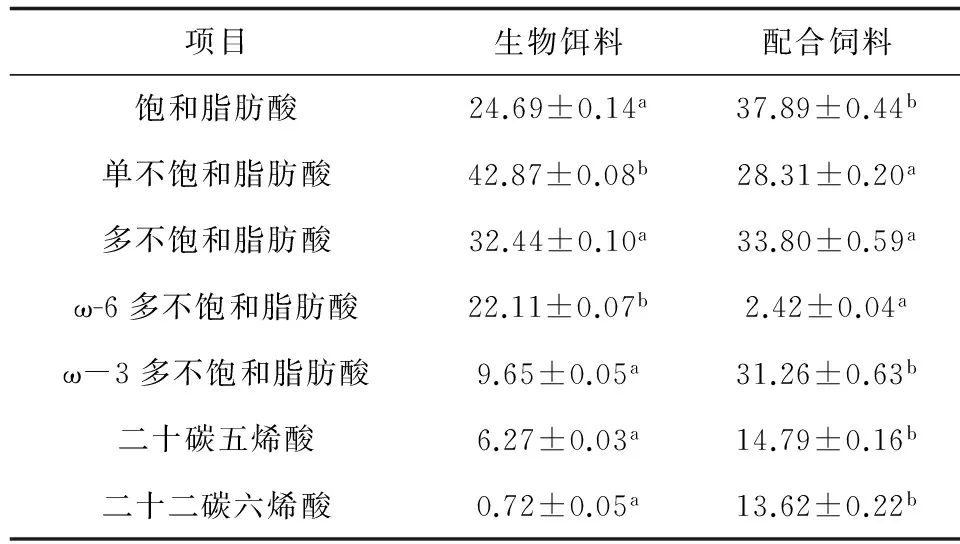
表2 鮮活大鹵蟲與日清配合飼料脂肪酸組成比較 %
2.2 不同轉餌組種苗生長情況
試驗結束時,轉餌4組種苗變異系數顯著大于其他各組(P<0.05),其他各組變異系數差異不顯著(P>0.05)(表3)。轉餌4組死亡率亦顯著大于轉餌1組和轉餌5組(P<0.05),而其他各組死亡率差異不顯著(P>0.05)。36—45日齡轉餌1組種苗特定生長率顯著小于其他各組(P<0.05),其他各組差異不顯著(P>0.05); 46—60日齡轉餌1組種苗特定生長率顯著大于其他各組(P<0.05),其他各組差異不顯著(P>0.05)。試驗結束時,轉餌1組種苗體質量和質量增加率最低,顯著低于其他各組(P<0.05),而轉餌2組又顯著低于轉餌4組和轉餌5組(P<0.05)。
2.3 不同轉餌模式對種苗體成分的影響
各轉餌組種苗水分和粗蛋白含量差異不顯著(P>0.05),轉餌5組種苗粗脂肪含顯著低于其他轉餌組(P<0.05)(表4)。
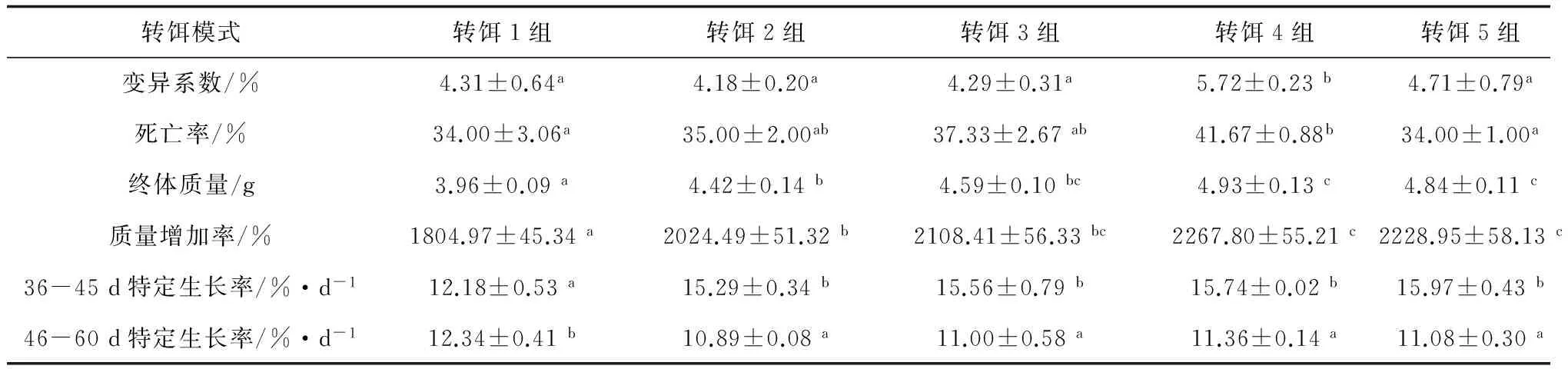
表3 不同轉餌模式組種苗生長情況
注:表中數據為3個平行組測定結果的平均值±標準差,同一行數據上標有相同字母表示差異不顯著(P>0.05),上標無相同字母則表示差異顯著(P<0.05).表4同.
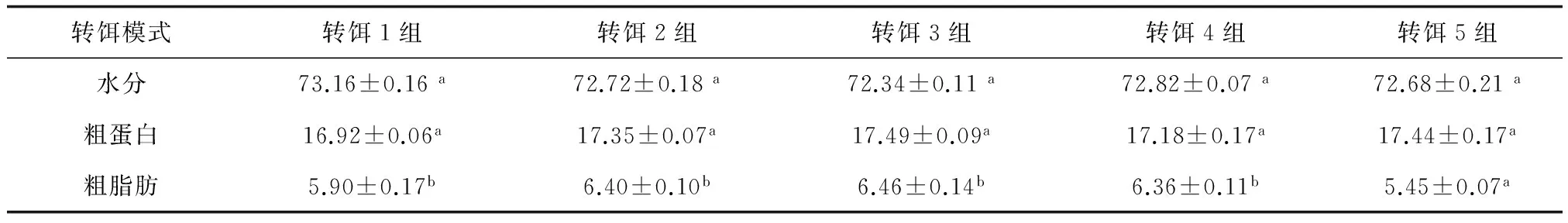
表4 不同轉餌模式對60日齡種苗體成分的影響 %
3 討論
3.1 轉餌模式對種苗特定生長率的影響
在5個轉餌組中,轉餌1組種苗死亡率最低,但60日齡時種苗的體質量亦最小,這可能是由于生物餌料存在營養缺陷,無法滿足種苗生長需要。由表2可見,生物餌料ω-3多不飽和脂肪酸、二十二碳六烯酸和二十碳五烯酸含量顯著低于配合飼料,而ω-3多不飽和脂肪酸是海水種苗的必需脂肪酸,其中以二十二碳六烯酸和二十碳五烯酸尤為重要[19],因此轉餌1組種苗在36—45日齡的特定生長率顯著低于投喂配合飼料的其他各組。在轉投配合飼料之后,轉餌1組種苗的特定生長率顯著高于其他轉餌組,這主要是由于轉餌1組種苗在46日齡時的體質量最小,因此特定生長率相對較高。
3.2 轉餌模式對種苗死亡率的影響
肉食性魚類常會發生相互殘食[20],尤其是在其種苗階段[21]。試驗中筆者觀察到較大斜帶石斑魚種苗會用嘴咬住其他較小個體的頭部并來回甩動,這種攻擊性行為常出現在每天第一次投餌前,與黑線鱈(Melanogrammusaeglefinus)[22]相互殘食行為相似。Hseu等[23]研究發現,個體大小差異超過30%時,斜帶石斑魚種苗就可能相互殘食,因此魚體長度變異系數[18]可作為海水魚種苗發生相互殘食的一個重要指標。試驗中轉餌4組的種苗變異系數顯著大于其他各組,導致組內相互殘食增加,使該組種苗存活率顯著低于其他各組。
3.3 斜帶石斑魚種苗不經過渡直接轉餌的可行性
對于36日齡種苗不經過渡直接進行配合飼料轉餌,觀察到種苗在前2 d的攝食較少,這可能是因為石斑魚喜食運動的食物,而配合飼料下沉速度相對較快,導致種苗在剛接觸時未能及時攝食[24]。自第3 d開始,種苗的攝食量逐漸增加,至第5 d,已經能迅速攝食剛投入的配合飼料。諸多學者認為,由于仔魚的消化器官尚未發育完善,消化酶分泌不足,導致其無法很好地消化和利用配合飼料[6,8,10,25],因此只要消化器官發育完善,對變態期或變態后種苗(0.5~0.75 g)直接進行轉餌亦可獲得成功[5,26-27]。Segner等[27]研究發現,與幼魚相比,仔魚是由于沒有胃而無法有效地消化、吸收配合飼料;而Hart等[28]對未進入變態期,但胃已完全分化的23日齡綠背鰈(Rhombosoleatapirina)進行轉餌,獲得了82.2%的成活率;因此Hart等[28]認為種苗轉餌的恰當時機應為胃完全分化時。吳金英等[29]對斜帶石斑魚種苗消化系統發育的觀察發現,4日齡仔魚消化系統已明顯分化成食道、胃和直腸等,而45—60日齡幼魚的胃已經很發達。筆者在試驗開始前曾采集36日齡斜帶石斑魚種苗消化道進行消化酶類檢測,發現此時種苗已經具有較高的消化酶活力,因此本試驗設計的直接轉餌從36日齡開始投喂配合飼料。試驗結果也顯示,36日齡斜帶石斑魚種苗從攝食生物餌料直接轉為攝食配合飼料,對其生長和存活未產生顯著影響。
3.4 轉餌模式對種苗體成分的影響
轉餌5組60日齡種苗粗脂肪含量顯著低于其他轉餌組,這可能與轉餌過程中魚體吸收的脂肪酸種類有關,攝食過兩種飼料(生物餌料和配合飼料)的種苗脂肪酸種類比較全面,而轉餌5組種苗僅吸收配合飼料中的脂肪酸,所以試驗魚體粗脂肪含量較低。Kolkovski等[30]的研究也表明,歐洲舌齒鱸 (D.labrax)仔魚在同時攝食配合飼料和豐年蟲無節幼體時魚體粗脂肪含量較高。
3.5 轉餌模式優缺點比較
本研究所采取的5種轉餌模式均能成功實現斜帶石斑種苗的轉餌,5種模式各有優缺點。轉餌1模式種苗死亡率最低,但種苗的質量增加率也最低;轉餌5模式種苗死亡率較低,種苗的質量增加率較高,但試驗魚粗脂肪含量顯著低于其他投喂組;轉餌4模式種苗的質量增加率最高,但種苗死亡率也最高;綜合比較各項指標,轉餌3模式的種苗質量增加率較高而種苗死亡率較低,是本研究中總體效果較好的轉餌模式。
[1] 周仁杰,林濤.斜帶石斑魚人工育苗技術試驗[J].臺灣海峽,2002,21(1):57-62.
[2] 王云新,黃國光,劉付永忠,等.斜帶石斑魚人工育苗試驗[J].漁業現代化,2003,30(6):14-15.
[3] 史海東,辛儉,毛國民,等.斜帶石斑魚人工育苗技術的初步研究[J].浙江海洋學院學報:自然科學版,2004,23(1):19-23.
[4] 張海發.斜帶石斑魚早期生活史及種苗繁育技術研究[D].廣州:中山大學,2005.
[5] Jeannine P L R, Jean C A,Laurence T,et al.Marine fish larvae feeding:formulated diets or live prey [J].Journal of the World Aquaculture Society,1993,24(2):211-224.
[6] Rosenlund G,Stoss J,Talbot C.Co-feeding marine fish larvae with inert and live diets [J].Aquaculture,1997,155(1/4):183-191.
[7] Engrola S,Figueira L,Concei??o L E C,et al.Co-feeding in Senegalese sole larvae with inert diet from mouth opening promotes growth at weaning [J].Aquaculture,2009,288(3/4):264-272.
[8] Cahu C L,Zambonino Infante J L.Substitution of live food by formulated diets in marine fish larvae [J].Aquaculture,2001,200(1/2):161-180.
[9] Alves T T,Cerqueira V R,Brown J A.Early weaning of fat snook (CentropomusparallelusPoey 1864) larvae [J].Aquaculture,2006,253(1/4):334-342.
[10] Kolkovski S.Digestive enzymes in fish larvae and juveniles-implications and applications to formulated diets [J].Aquaculture,2001,200(1/2):181-201.
[11] Barnabé G,Guissi A.Adaptations of the feeding behaviour of larvae of the sea bass,Dicentrarchuslabrax(L.),to an alternating live-food/compound-food feeding regime [J].Aquaculture Research,1994,25(5):537-546.
[12] Curnow J,King J,Bosmans J,et al.The effect of reducedArtemiaand rotifer use facilitated by a new microdiet in the rearing of barramundiLatescalcarifer(BLOCH) larvae [J].Aquaculture,2006,257(1/4):204-213.
[13] Nhu V C,Dierckens K,Nguyen H T,et al.Effect of early co-feeding and different weaning diets on the performance of cobia (Rachycentroncanadum) larvae and juveniles [J].Aquaculture,2010,305(1):52-58.
[14] Bonaldo A,Parma L,Badiani A,et al.Very early weaning of common sole (SoleasoleaL.) larvae by means of different feeding regimes and three commercial microdiets:influence on performances,metamorphosis development and tank hygiene [J].Aquaculture, 2011,321 (3/4):237-244.
[15] 趙金柱,艾慶輝,麥康森,等.微粒飼料替代生物餌料對大黃魚稚魚生長,存活和消化酶活力的影響 [J].水產學報,2008,32(1):91-97.
[16] 張麗英.飼料分析及飼料質量檢測技術[M].北京:中國農業大學出版社,2002:49-59.
[17] 胡述楫,胡人衛.尼羅羅非魚魚脂中脂肪酸的鑒定與評價[J].西南農業學報,1997,10(1):115-118.
[18] Folkvord A,Otter? H.Effects of initial size distribution,day length,and feeding frequency on growth,survival,and cannibalism in juvenile Atlantic cod (GadusmorhuL.) [J].Aquaculture,1993,114(3):243-260.
[19] Watanabe T,Izquierdo M S,Takeuchi T,et al.Comparison between eicosapentaenoic and docosahexaenoic acids in terms of essential fatty acid efficacy in larval red seabream [J].Nippon Suisan Gakkaishi,1989,55(9):1635-1640.
[20] Smith C,Reay P.Cannibalism in teleost fish [J].Reviews in Fish Biology and Fisheries,1991,1(1):41-64.
[21] Hecht T,Pienaar A G.A review of cannibalism and its implications in fish larviculture [J].Journal of the World Aquaculture Society,1993,24(2):246-261.
[22] Hamlin H,Kling L.The culture and early weaning of larval haddock (Melanogrammusaeglefinus) using a microparticulate diet [J].Aquaculture,2001,201(1/2):61-72.
[23] Hseu J R,Chang H F,Ting Y Y.Morphometric prediction of cannibalism in larviculture of orange-spotted grouper,Epinepheluscoioides[J].Aquaculture,2003,218(1/4):203-207.
[25] Lee G W Y,Litvak M K.Weaning of metamorphosed winter flounder (Pleuronectesamericanus) reared in the laboratory:comparison of two commercial artificial diets on growth,survival and conversion efficiency [J].Aquaculture,1996,144(1/3):251-263.
[26] Daniels H V,Hodson R G.Weaning success of southern flounder juveniles:effect of change over period and diet type on growth and survival [J].North American Journal of Aquaculture,1999,61(1):47-50.
[27] Segner H,Roesch R,Verreth J,et al.Larval nutritional physiology:studies with Ckzrius gariepinus,CoregonuslauaretusandScophthalmusmaximus[J].J World Aquaculture Society,1993,24(2):121-134.
[28] Hart P R, Purser G J.Weaning of hatchery-reared greenback flounder (RhombosoleatapirinaGünther) from live to artificial diets:effects of age and duration of the change over period [J].Aquaculture,1996,145(1/4):171-181.
[29] 吳金英,林浩然.斜帶石斑魚消化系統胚后發育的組織學研究 [J].水產學報,2003,27(1):7-12.
[30] Kolkovski S,Tandler A,Izquierdo M S.Effects of live food and dietary digestive enzymes on the efficiency of microdiets for seabass (Dicentrarchuslabrax) larvae [J].Aquaculture,1997,148(4):313-322.
WeaningofOrange-spottedGrouper(Epinepheluscoioides)LarvaebyMeansofDifferentFeedingRegimes
ZHOU Hui1,2,CHEN Gang1,2,SUN Kungang1
( 1.College of Fisheries,Guangdong Ocean University,Zhanjiang 524025,China; 2.Guangdong Provincial Key Lab of Pathogenic Biology and Epidemiology for Aquatic Animals,Zhanjiang 524025,China )
The suitable feeding regime was studied for weaning of orange-spotted grouper (Epinepheluscoioides) larvae 36 days after hatch (DAH) with average body weight of (0.21±0.01) g.The larvae were fed by five feeding regimes as following:in regime R1,the larvae fed live food (LP) in the first ten days,and then weaned onto inert diet (ID); in regimes R2,R3 and R4,the larvae fed the mixed diet in the first ten days (R2:25% ID +75% LP; R3:50% ID +50% LP; R4:75% ID +25% LP),and then weaned onto ID; in regime R5,the larvae fed ID in 36 DAH (at the end of the weaning experiment).The results showed that there was the maximal mortality (DR) in the larvae in regime R4 due to significantly higher coefficient of variance (CV) of body length (P<0.05),significantly higher than those in other regimes (P<0.05),and there were no significant differences in CV and DR among other regimes (P>0.05).The minimal final body weight and weight gain rate (WGR) were observed in the larvae in regime R1 at the end of the experiment,significantly lower than in other regimes (P<0.05),and significantly lower in regime R2 than regimes R4 and R5 (P<0.05).The larvae in regime R1 had significantly lower specific growth rate than the larvae in other regimes did during 36—45 DAH (P<0.05),and significantly higher than in other regimes during 46—60 DAH (P<0.05).After the experiment,there were no significant differences in moisture and crude protein content in the larvae among all feeding regimes (P>0.05). However,the larvae in regime R5 had significantly lower crude lipid content than the larvae in other regimes did (P<0.05).It is concluded that feeding regime R3 is the optimal feeding regime for the larvae which showed higher WGR and lower DR.
Epinepheluscoioides;co-feeding;live prey;formulated feed
10.16378/j.cnki.1003-1111.2016.02.010
S965.334
A
1003-1111(2016)02-0152-05
2015-07-31;
2015-10-08.
國家海洋局海洋公益性行業科研專項項目(201205028);廣東省科技廳省部產學研結合項目(2013B090700010);廣東省海洋經濟創新發展區域示范專項經費(GD2012-A02-003).
周暉(1978-),男,博士;研究方向:魚類生理生態學.E-mail:346005548@qq.com.通訊作者:陳剛(1961-),男,教授,博士生導師;研究方向:水產經濟動物生物學及種苗工程.E-mail:cheng@gdou.edu.cn.
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
科學大眾(2021年9期)2021-07-16 07:02:54
軍事文摘(2020年20期)2020-11-28 11:42:50
航空世界(2020年10期)2020-01-19 14:36:20
小學科學(學生版)(2018年3期)2018-04-18 12:34:19
知識經濟·中國直銷(2017年10期)2017-11-07 02:39:52
知識經濟·中國直銷(2017年3期)2017-04-16 03:08:01
文學少年(有聲彩繪)(2017年2期)2017-03-06 08:10:54
